
曼陀罗光合特性研究
2011-12-31 13:49于晓霞阮成江
植物资源与环境学报 2011年1期
于晓霞,阮成江
曼陀罗 (Datura stramonium L.)属 茄科(Solanaceae)曼陀罗属(Datura L.)1年生直立大型草本植物,主要分布于热带和亚热带地区,温带地区也有分布,常见于路边、草地和新翻地。曼陀罗种子含油量约22.85%,其种子脂肪油主要含亚油酸、棕榈酸、硬脂酸、油酸、肉豆蔻酸等脂肪酸成分[1-2],是潜在的能源植物。资料显示[3-6],相关学者对于曼陀罗资源已经进行了大量研究,不仅总结了曼陀罗的栽培技术和应用价值,还在其化学成分和药理作用等方面取得了一些研究成果。目前,野生曼陀罗的资源量处于急剧下降的状态,因此,对曼陀罗资源研究的首要任务就是对其资源的保护。首先是就地保护,扼制过度采挖;其次就是要加强对野生曼陀罗人工栽培技术的研究[7]。
可再生能源植物的研究是当前新型能源研发的重要内容之一,合理的人工栽培方式能够为曼陀罗的开发利用奠定基础。通过人工控制光照强度和CO2浓度等方式提高曼陀罗光合作用效率,促进其生物量的积累,有望提高曼陀罗的能源利用率。为此,作者探讨了曼陀罗净光合速率的日、月变化及其对光合有效辐射强度和CO2浓度的响应,旨在了解曼陀罗的光合生理基本特性,为曼陀罗的人工栽培提供理论依据,并为曼陀罗作为潜在能源植物的开发利用研究提供科学依据和实验基础。
1 材料和方法
1.1 材料
于2010年6月至9月在大连民族学院温室外选择野外自然生长的曼陀罗供试。该区域的土壤为棕壤土,土壤酸碱度为pH 7.91;气候属北温带大陆性季风气候,但具有海洋性气候特征;年平均气温约10℃,其中8月最热,平均气温24℃,年极端最高气温约35℃;年平均降水量550~950 mm。
1.2 方法
1.2.1 光合特性日、月变化的测定 于2010年6月至9月,每月中旬选取典型晴朗天气2 d,采用LI-6400型便携式光合测定仪(美国LI-COR公司生产)进行光合指标的测定。实验在连体条件下进行[8],选用开路气流法(气源引自地上空3 m处)[9],自然光源,叶室面积2 cm×3 cm。在每个测定日选取大小、体形特征一致的植株5株,每株按东、南、西、北4个方向各选4片成熟叶片[10],从当地时间8:00至18:00每隔1 h测1次净光合速率(Pn),同时测定叶片气孔导度(Cs)、叶室内的光合有效辐射强度(PAR)等相关指标。每个指标重复测定3次,结果取平均值。
1.2.2 叶绿素含量的测定 参照文献[11]的方法测定叶绿素含量。采集新鲜叶片,擦净表面后剪碎(去掉中脉),混匀;称取0.3 g剪碎的叶片,置于25 mL体积分数95%乙醇中避光浸泡至叶片全部变白,离心,上清液用Lambda25型紫外/可见分光光度计(美国PerkinElmer公司生产)于波长665和649 nm处分别测定吸光值,并计算叶绿素a和b的含量。实验设3次重复。
1.2.3 净光合速率的光响应曲线和CO2响应曲线的绘制 在实验期间选取典型晴朗天气3 d,于上午9:00至12:00,选长势基本一致的植株3株,取从顶部数第1片完全展开成熟叶片进行净光合速率的测定,结果取平均值。
利用LI-6400型便携式光合测定仪的自动light-curve曲线测定功能绘制净光合速率的光响应曲线。小钢瓶注入系统控制参比室的CO2浓度为400μmol·mol-1,根据净光合速率日变化值,确定其大概的饱和光强,并将待测叶片在接近饱和光强下诱导20 min(仪器自带的红蓝光源)以充分活化光合系统,然后参照文献[12]的测量水平和条件将红蓝光源从高到低设定一系列光合有效辐射强度(PAR)梯度(1 800、1 500、1 200、1 000、800、600、400、200、150、100、50、30、10、0μmol·m-2·s-1),最小和最大等待时间分别为60和180 s,CO2匹配值20μmol·mol-1,测定曼陀罗叶片的净光合速率。
采用上述方法参照文献[12]的测量水平和条件,利用LI-6400型便携式光合测定仪的自动Acicurve曲线测定功能绘制净光合速率的CO2响应曲线。红蓝光源设定的光合有效辐射强度为 800 μmol·m-2·s-1;考虑到植物对 CO2的适应性,将CO2浓度设定为400、300、200、150、100、50、30、10、400、400、600、800、1 000、1 200、1 500、1 800μmol· mol-1,其他测定条件与光响应曲线的测定条件相同。
1.3 数据处理
所有数据均通过Excel软件进行整理,使用SPSS 17.0软件进行数据的相关性分析、线性方程回归分析以及其他统计分析,采用Excel软件进行图表处理。


图1 6月份至9月份曼陀罗叶片净光合速率的日变化Fig.1 Daily change of net photosynthetic rate of Datura stramonium L.leaves from June to September
在其他因子变化不大时,净光合速率的CO2响应曲线也采用非直角双曲线进行拟合,公式同上,其中: A为净光合速率(μmol·m-2·s-1);α为羧化效率; Amax为最大净光合速率(μmol·m-2·s-1);Q为CO2浓度(μmol·m-2·s-1);k为响应曲线曲角;Rday为暗呼吸速率(μmol·m-2·s-1)。
2 结果和分析
2.1 曼陀罗光合特性指标的日变化和月变化分析
2.1.1 各月份净光合速率的日变化 在6月份至9月份曼陀罗叶片净光合速率(Pn)的日变化见图1。由图1可知,实验期间,曼陀罗净光合速率的日变化均表现为“单峰型”,不存在显著的“午休”现象,其净光合速率在上午逐渐升高,中午达到最大,下午逐渐降低。6、7和8月份曼陀罗叶片净光合速率的峰值均出现在11:00左右,平均峰值分别为15.24、19.01和9.85μmol·m-2·s-1;9月份曼陀罗植株叶片净光合速率的峰值出现在10:00左右,平均峰值为6.30μmol·m-2·s-1。7月份的日净光合速率整体上高于其他3个月份。
2.1.2 光合有效辐射强度和气孔导度的日变化 光合有效辐射强度和曼陀罗叶片气孔导度的日变化见图2-A和B。由图2-A可见:光合有效辐射强度的日变化趋势与净光合速率基本一致,为“单峰型”,均在达到最高值之后逐渐下降,峰值出现在11:00左右。曼陀罗叶片气孔导度的日变化则表现为“双峰型”(图2-B),在每天的早、晚较低,在上午随时间的推移气孔导度随之增高,并在12:00左右达到最高峰,随后开始下降,在14:00左右再次出现1个小的峰值,随后加速下降,直至傍晚时分气孔基本关闭,气孔导度降至最低值。
2.1.3 净光合速率和叶绿素含量的月变化 6月份至9月份曼陀罗叶片净光合速率和叶绿素含量的月变化见表1。由表1可知:曼陀罗叶片的月平均净光合速率在6月份时较高,7月份达到最大,8月份开始下降,9月份则较低,植物开始出现枯萎现象。差异显著性分析结果显示,不同月份曼陀罗叶片的净光合速率月平均值均有极显著差异(P<0.01)。
6月份至9月份曼陀罗叶片叶绿素含量(包括叶绿素a、b和总叶绿素)的月平均值的变化趋势与净光合速率月平均值的变化基本一致(表1),最高值也出现在7月份。9月份叶绿素含量较低,并与7月份含量具有极显著差异(P<0.01)。但曼陀罗叶片叶绿素a、b和总叶绿素含量的月变化略有差异。
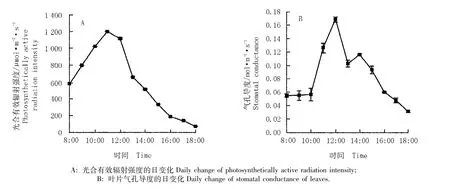
图2 光合有效辐射强度和曼陀罗叶片气孔导度的日变化Fig.2 Daily change of photosynthetically active radiation intensity and stomatal conductance of Datura stramonium L.leaves
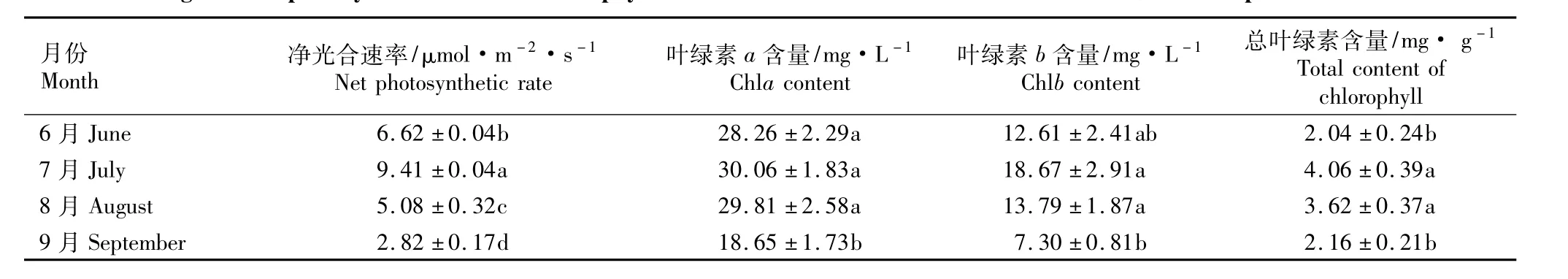
表1 6月份至9月份曼陀罗叶片净光合速率和叶绿素含量的变化1)Table 1 Changes of net photosynthetic rate and chlorophyll content of Datura stramonium L.leaves from June to September1)
2.2 曼陀罗叶片净光合速率的光响应曲线分析
经SPSS软件统计分析,采用非直角双曲线模型理论公式对曼陀罗叶片净光合速率的光响应曲线进行拟合,得出公式中的k值为0.951、Amax值为19.805 μmol·m-2·s-1、Rday值为0.666μmol·m-2·s-1、α值为0.030,拟合曲线的R2=0.968。根据净光合速率的拟合预测值(Pv)绘制光响应曲线(图3-A),与净光合速率实测值(Pn)拟合结果较好,表明用非直角双曲线模型拟合得出的曼陀罗叶片净光合速率的光响应曲线较为理想,能够完整地反映出在外界良好空气条件下叶片净光合速率随光合有效辐射强度的变化趋势。在低光合有效辐射强度下,曼陀罗叶片的净光合速率随光合有效辐射强度的增大呈线性升高,因此,在光合有效辐射强度小于200μmol·m-2·s-1的条件下对曼陀罗叶片净光合速率(y)和光合有效辐射强度(x)进行直线回归分析[15-16],得到的回归方程为:y=-0.665 9+0.029 7x,R2=0.979 9。由此方程计算得出:曼陀罗叶片的光补偿点为22.42μmol· m-2·s-1,光饱和点为689.26μmol·m-2·s-1。
2.3 曼陀罗叶片净光合速率的CO2响应曲线分析
采用非直角双曲线模型理论公式对曼陀罗叶片净光合速率的CO2响应曲线进行拟合,得出公式中的k值为1.000、Amax值为21.636μmol·m-2·s-1、α值为0.017、Rday值为1.286μmol·m-2·s-1,拟合曲线的 R2=0.979。根据净光合速率的拟合预测值(Pv)绘制CO2响应曲线(图3-B),与净光合速率实测值(Pn)拟合效果较好,表明用非直角双曲线模型拟合得出的曼陀罗叶片净光合速率的CO2响应曲线较为理想,能够完整反映出在适当光照条件下曼陀罗叶片净光合速率随CO2浓度的变化趋势。在低浓度CO2条件下,曼陀罗的净光合速率随CO2浓度增大呈线性升高,因此,在CO2浓度小于400μmol·mol-1的条件下对曼陀罗叶片净光合速率(y)和CO2浓度(x)进行直线回归分析,得到的回归方程为:y=-1.273 9+ 0.017 2x,R2=0.984 4。由此回归方程计算得出:曼陀罗植株叶片的CO2补偿点为74.06μmol·mol-1,CO2饱和点为1 331.97μmol·mol-1。

图3 曼陀罗叶片净光合速率的光响应曲线及CO2响应曲线Fig.3 Light and CO2 response curves of net photosynthetic rate of Datura stramonium L.leaves
3 讨论和结论
本研究结果表明,在6月份至9月份,曼陀罗叶片净光合速率的日变化均呈“单峰型”,峰值大约出现在11:00左右,其中,7月份的净光合速率峰值最高,为19.01μmol·m-2·s-1。净光合速率和光合有效辐射强度的日变化均呈先升后降的趋势,这是因为光合有效辐射强度发生改变可引起气温、空气相对湿度等环境因子的变化,而空气相对湿度通过影响气孔开闭来控制CO2流量,从而影响植物的光合速率[17-18]。朱秀苗[7]的研究结果显示:产自青海省的曼陀罗叶片净光合速率的日变化呈“双峰型”,与作者的研究结果有所不同。由于植物光合作用是自身生理特性与环境条件综合作用的结果[19],因此,应对不同产地及不同生境条件下曼陀罗叶片的光合作用特性进行比较后,才能对造成这种差异的具体原因加以验证。
在6月份至9月份间曼陀罗叶片的净光合速率月平均值有极显著差异(P<0.01),这可能是多种环境因子综合作用造成的。这几个月份曼陀罗叶片中叶绿素含量的变化趋势与净光合速率的变化趋势基本一致。有研究者认为,正常情况下植物不会缺少光合作用所需的叶绿素,当叶绿素含量超过某一个限制值后,叶绿素含量的高低与净光合速率的大小则没有必然的相关性[20]。关于曼陀罗叶片光合作用过程中叶绿素含量的限制值,以及达到限制值前后净光合速率对叶绿素含量的响应情况还需进一步研究。
本研究结果表明,曼陀罗叶片净光合速率的光响应曲线和CO2响应曲线的变化趋势基本一致,这与其他植物的光合特性是相同的[12]。具体表现为:在一定阈值内,随着光合有效辐射强度的增加,净光合速率增大;当光合有效辐射强度超过该阈值后,随着光合有效辐射强度的进一步升高,净光合速率的增长速率变缓;当光合有效辐射强度达到一定数值后,净光合速率基本稳定在一定水平。净光合速率与CO2浓度之间也有同样的变化规律,随CO2浓度的升高,净光合速率首先表现为迅速升高,然后增长变缓,当CO2浓度增加到一定数值后,气孔阻力增大,CO2出现饱和现象,净光合速率稳定在一定水平。
曼陀罗叶片光补偿点为22.42μmol·m-2·s-1,光饱和点为689.26μmol·m-2·s-1,说明曼陀罗应属于阳性植物,但具有一定的耐阴性。曼陀罗叶片的CO2补偿点为74.06μmol·mol-1,CO2饱和点为1 331.97μmol·mol-1,比一般C3植物的CO2补偿点(30~70μmol·mol-1)[21]偏高,说明自然条件下曼陀罗的CO2同化能力较弱,不利于光合产物的积累。由于本研究仅在光合有效辐射强度800μmol·m-2· s-1的条件下分析了CO2浓度对曼陀罗叶片净光合速率的影响,研究范围不够全面,因此,对在不同光合有效辐射强度条件下曼陀罗叶片净光合速率的变化尚需深入全面的研究,以便了解曼陀罗在各种条件下的光合特性[22],从而为曼陀罗的人工栽培提供理论依据。
[1]邢维贺,阮成江,李 贺.5种能源植物种子含油量与脂肪酸组成[J].可再生能源,2010,28(2):62-66.
[2]张美淑,金大勇.曼陀罗在植保领域的利用现状及展望[J].中国林副特产,2005(3):80.
[3]娄凤菊,连立峰,周玉秋,等.曼陀罗的栽培技术及应用[J].特种经济动植物,2004,7(5):30.
[4]张宏利,杨学军,刘文国,等.曼陀罗化学成分与生物活性研究现状及展望[J].西北林学院学报,2004,19(2):98-102.
[5]边振考,王翠珍,李 芹.曼陀罗叶、种子可供药用的初步试验[J].时珍国医国药,1998,9(6):521-522.
[6]徐 宁,冉俊祥,杨占臣,等.曼陀罗毒性的研究进展[J].检验检疫科学,2009,19(1):62-65.
[7]朱秀苗.青海省曼陀罗植物资源特性及开发利用前景的研究[D].西宁:青海师范大学生命与地理科学学院,2009.
[8]岳 瑾.菜用枸杞光合特性研究[D].保定:河北农业大学园艺学院,2006.
[9]阮成江,李代琼.半干旱黄土丘陵区沙棘的光合特性及其影响因子[J].植物资源与环境学报,2000,9(1):16-21.
[10]王文佳,吕岩桢,郭传友.构树光合生理特征的初步研究[J].安徽农学通报,2010,16(1):53,100.
[11]李合生.植物生理生化实验原理和技术[M].1版.北京:高等教育出版社,2000:134-137.
[12]宋庆安,童方平,易霭琴,等.虎杖光合生理生态特性研究[J].中国农学通报,2006,22(12):71-76.
[13]刘宇锋,萧浪涛,童建华,等.非直线双曲线模型在光合光响应曲线数据分析中的应用[J].中国农学通报,2005,21(8): 76-79.
[14]陆佩玲,于 强,罗 毅,等.冬小麦光合作用的光响应曲线的拟合[J].中国农业气象,2001,22(2):12-14.
[15]中国科学院上海植物生理研究所,上海市植物生理学会.现代植物生理学实验指南[M].北京:科学出版社,1999:89-91.
[16]Richardson A D,Berlyn G P.Spectral reflectance and photosynthetic properties of Betula papyrifera(Betulaceae)leaves along an elevational gradient on Mt.Mansfield,Vermont,USA[J].American Journal of Botany,2002,89(1):88-94.
[17]刘志龙,方建民,虞木奎,等.三种林-茶复合林分中环境因子和茶的光合特性参数的日变化规律[J].植物资源与环境学报,2009,18(2):62-67.
[18]徐克章,曹正菊,陈 星,等.人参叶片光合作用和气孔开闭日变化的研究[J].植物生理学通讯,1987(3):35-36.
[19]张 颖,呼天明.普那菊苣夏季光合速率日变化及其影响因子的研究[J].西北农业学报,2007,16(5):184-187.
[20]胡利明.柑橘光合特性的研究及C4光合途径的初步探讨[D].武汉:华中农业大学园艺林学学院,2007.
[21]潘瑞炽.植物生理学[M].5版.北京:高等教育出版社,2004:80-81.
[22]孙存华,李 扬,贺鸿雁,等.藜的光合特性研究[J].西北植物学报,2006,26(2):416-419.
猜你喜欢
现代农业研究(2021年4期)2021-04-11
空间科学学报(2020年6期)2020-07-21
农业与技术(2020年12期)2020-07-04
河北渔业(2020年2期)2020-04-08
农学学报(2019年1期)2019-02-21
老年教育(2018年12期)2018-12-29
新课程·下旬(2017年7期)2017-08-14
天津城建大学学报(2015年5期)2015-12-09
空间控制技术与应用(2014年2期)2014-04-17
绿色科技(2012年11期)2012-11-16
