
海拉尔河浮游植物功能群时空特征及其与水环境因子的关系
2023-10-19 06:05刘子暄王莹莹赵昶常畅然窦华山陈星瑶陶玉龙马成学
大连海洋大学学报 2023年4期
刘子暄,王莹莹,赵昶,常畅然,窦华山,陈星瑶,陶玉龙,马成学*
(1.东北林业大学 野生动物与自然保护地学院,黑龙江 哈尔滨 150040;2.呼伦湖国家级自然保护区管理局,内蒙古 海拉尔 021008)
浮游植物是水生态系统中的初级生产者,其物种变化、群落演替及其与水环境因子间的关系可以直观地反映水生态系统的动态变化[1]。相对于其他方法,Reynolds等[2]和Padisák等[3]提出并完善了浮游藻类功能群理论,共划分和描述了39 组浮游植物功能群。这种方法能够更加准确地描述浮游植物的生境特征,表现浮游植物群落同水环境因子的相关性,预测特定生境下浮游植物的群落分布状况。
河流生态系统是地球上最为复杂的生态系统之一,是陆生生态系统和水生生态系统之间物质循环、能量流动和信息传递的重要媒介,人类活动通过直接或间接的方式影响着河流生态系统的健康[4]。近年来,基于浮游植物功能群理论对中国热带[5]、亚热带[6-12]、温带[13-15]和北方寒区河流[16-19]不同类型的水体研究逐渐增多。作为北方寒区呼伦湖主要入湖河流之一的海拉尔河,与其相关的研究主要集中在流域水质评价[20]和水文学[21]等方面,而利用生物指标[22]进行的相关研究较少。李星醇等[23]首次将浮游植物功能群理论应用于海拉尔河,但其缺少在时间尺度上完整的浮游植物功能群变化研究。海拉尔河流域是呼伦贝尔市人口及经济产业集中带,所面临的人口、经济与环境发展问题更加复杂且严峻。因此,作为额尔古纳河重要上源及呼伦湖主要入湖河流之一的海拉尔河,其水生态环境的优良程度对其下游及呼伦湖乃至整个呼伦贝尔市水生态安全具有重要意义。本研究中,对海拉尔河水域浮游植物群落进行了功能群划分,并分析了功能群与水环境因子间的关系,以期为海拉尔河未来的浮游植物功能群研究提供一定的数据参考,同时尝试从浮游植物功能群角度分析海拉尔河的主要污染原因,进而维护整个呼伦湖流域的生态安全。
1 材料与方法
1.1 调查区域及采样站位的设置
海拉尔河流域(117° 48′30″ E~122° 28′ 05″ E,47°32′30″N~50°16′02″N)位于内蒙古自治区呼伦贝尔市西南部,发源于大兴安岭雷勒奇克山西麓,自东向西流向,至扎赛诺尔区北部阿巴该图山附近右转向北,是额尔古纳河上源之一,流域面积为5.48万km2。海拉尔河干流全长1 430 km,由东向西依次流经牙克石市、海拉尔区、陈巴尔虎旗和新巴尔虎左旗,汇入额尔古纳河;其主要支流有免渡河和伊敏河等[20]。流域属中温带半湿润半干旱大陆性季风气候,多年年均气温和降水量分别为5.34 ℃和348 mm,冬季漫长,冬雪期从每年10月末—11月上中旬开始,次年4—5月初结束[20]。2007年实施的呼伦湖水资源配置工程(引河济湖工程),引入海拉尔河河水,使呼伦湖恢复了与海拉尔河的水文联系。
根据海拉尔河的水体和水域范围的实际情况,以上、中、下游为依据,共设置了14个采样点。上游点位分别为S1(海拉尔河上游)、S2(牙克石),并包括上游支流免渡河点位S3(免渡河上游)、S4(免渡河下游),中游点位为S5(海拉尔河中游)、S6(海拉尔桥)、S7(北山口)和S8(呼和诺尔),并包括中游支流伊敏河点位S9(伊敏河上游)、S10(伊敏河中游)和S11(伊敏河下游),下游点位为S12(东乌珠尔)、S13(嵯岗)和S14(海拉尔河引河)。具体采样点设置见图1。
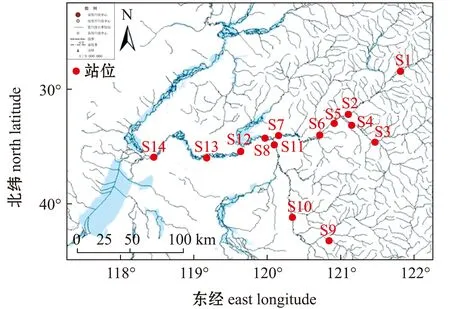
图1 海拉尔河流域采样点分布Fig.1 Sampling sites in Hailaer River
1.2 方法
1.2.1 样品采集与鉴定 浮游植物采集使用5 L采水器采集上、中、下层混合水样1 L,加入15 mL鲁哥氏液进行固定。带回实验室的水样静置沉淀72 h后,用虹吸管吸去上清液至沉淀物体积约50 mL,放入标本瓶中继续静置沉淀48 h后,定容至30 mL。在光学显微镜下进行物种鉴定、细胞计数等分析操作。浮游植物种类鉴定参考《中国淡水藻类:系统、分类及生态》[24]等相关文献。
依照文献[25],计算获得各采样点浮游植物的生物量。
1.2.2 水环境因子的测定 水环境因子的测定采用现场测定与实验室测定相结合的方式。现场使用表面水温计对水温(WT)进行测定,使用YSI-6600多功能水质分析仪对pH、溶解氧(DO)、电导率(Cond)进行测定。各采样点采集2 L水样,低温条件下48 h内送至实验室,按照《水和废水监测分析方法》中的方法测定总氮(TN)、总磷(TP)、化学需氧量(CODCr)和高锰酸盐指数(CODMn)。
1.2.3 除趋势对应分析和冗余分析 对浮游植物重要功能群生物量数据和水环境因子数据(除pH)进行量纲化处理[lg(x+1)]后,采用Canoco 4.5软件对生物量和水环境因子数据进行除趋势对应分析(detrended correspondence analysis,DCA)。处理后4个排序轴梯度长度最大为1.319,小于3.0,再通过蒙特卡洛置换检验(Monte carlo cermutation test,MCPT),采用冗余分析(redundancy analysis,RDA)获得功能群与环境因子相关性的数据[26]。
1.2.4 浮游植物功能群划分 浮游植物功能群的分类方法参照Reynolds[2]和Padisk等[3]提出的方法,将相对生物量大于10%的功能群定义为重要功能群。
1.3 数据处理
浮游植物数据的统计和分析采用Excel 2010软件进行初步整理。采用SPSS 19.0软件进行单因素方差分析,采用Waller-Duncan法进行多重比较,分析水环境因子和生物量的时空差异显著性,显著性水平设为0.05;使用重要浮游植物功能群,通过Pearson相关分析研究浮游植物重要功能群与水环境因子间的相关性。
2 结果与分析
2.1 海拉尔河水环境因子的分布特征
海拉尔河水环境因子的时空变化见表1。从季节分布看:海拉尔河春、夏、秋3个季节WT及DO有显著性差异(P<0.05),其他水环境因子均无显著性差异(P>0.05);春季DO、CODCr平均值明显高于其他季节,且CODCr值达到了国家Ⅴ类水标准(40 mg/L);夏季WT、TP、TN平均值明显高于其他季节;秋季CODMn平均值明显高于其他季节,且超过国家Ⅲ类水标准(6 mg/L),接近国家Ⅳ类水标准(10 mg/L)(表1)。
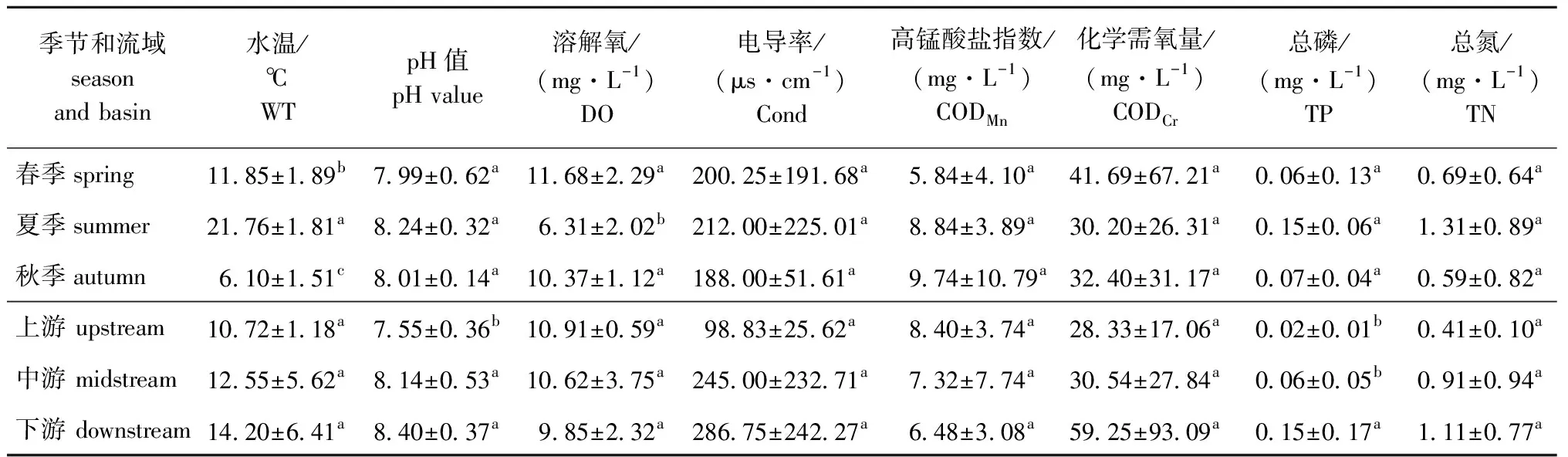
表1 海拉尔河环境因子的时空变化Tab.1 Temporal and spatial variation in environmental factors in the Hailaer River
从空间分布看:海拉尔河上、中、下游水环境因子pH及TP有显著性差异(P<0.05),其他水环境因子均无显著性差异(P>0.05);WT、pH、Cond、CODCr、TP、TN均呈现下游>中游>上游的趋势,下游水域氮磷含量明显增加;DO、CODMn均呈现上游>中游>下游的趋势,海拉尔河流域CODMn平均值较高,超过国家Ⅲ类水标准(6 mg/L),CODCr平均值超过国家Ⅲ类水标准(20 mg/L),下游CODCr达到国家Ⅴ类水标准(40 mg/L)。
2.2 海拉尔河浮游植物功能群分布
海拉尔河浮游植物生物量时空分布结果如表2所示,时间尺度上生物量呈现夏季>春季>秋季的趋势,且春夏两季生物量显著高于秋季;空间尺度上,平均生物量呈现下游>中游>上游的趋势(P<0.05),且下游平均生物量显著高于上游和中游(P<0.05)。
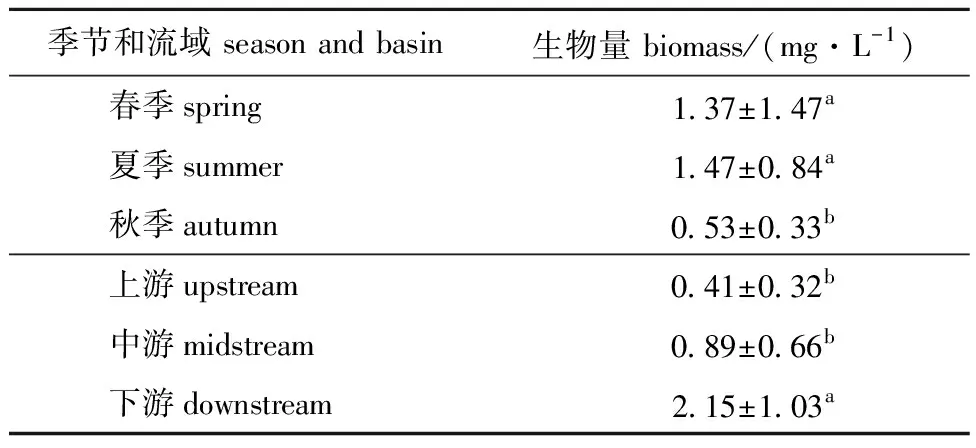
表2 海拉尔河浮游植物平均生物量的时空变化
海拉尔河水域春、夏、秋3个季节共鉴定出6门80种浮游植物,可划分为20个功能群,功能群组成见表3。对整体功能群生物量进行筛选,将每个季节采样点中相对生物量>10%的功能群定义为重要功能群,则海拉尔河春、夏、秋3个季节共筛选出重要功能群4个,分别为功能群C、D、MP、X2,重要浮游植物功能群春、夏、秋3个季节相对生物量时空分布见图2。其中,海拉尔河重要功能群季节组成变化不明显,时间变化呈现C+X2+D(春季)→X2+MP+D+C(夏季)→D+C+MP(秋季)。夏季浮游植物生物量高于春季,主要是由于春季到夏季功能群C生物量极显著减少(7.69 mg/L→2.42 mg/L)(P<0.01),功能群X2生物量显著增加(4.20 mg/L→7.48 mg/L),而其余功能群生物量变化不明显;秋季浮游植物生物量最少,主要是由于温度的显著降低导致整体生物量显著减少。
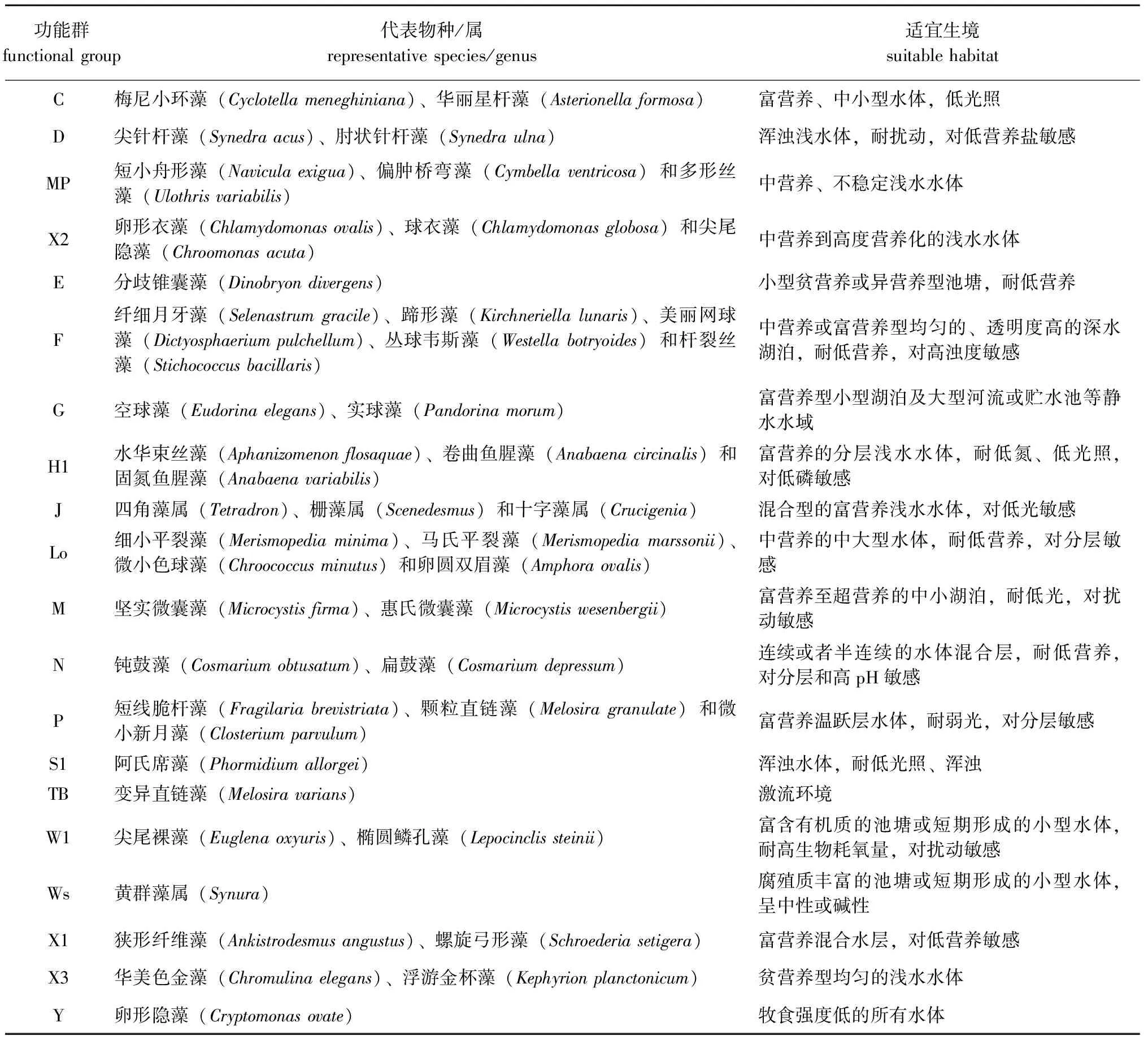
表3 海拉尔河浮游植物功能群组成Tab.3 Composition of phytoplankton functional groups in the Hailaer River

图2 海拉尔河浮游植物功能群相对生物量的时空分布Fig.2 Temporal and spatial distribution of relative biomass of phytoplankton functional groups in the Hailaer River
2.3 海拉尔河重要浮游植物功能群与水环境因子的相关关系
2.3.1 Pearson相关性分析 从表4可见,重要功能群主要与WT、pH、Cond、CODCr、TP和TN相关;功能群C与pH呈显著正相关(P<0.05);功能群D与WT和pH呈极显著正相关(P<0.01);功能群MP与WT呈极显著正相关(P<0.01);功能群X2与WT和TN呈显著正相关(P<0.05),与pH、Cond、CODCr和TP呈极显著正相关(P<0.01);其他功能群则与CODCr呈显著正相关(P<0.05),与pH和TP呈极显著正相关(P<0.01)。
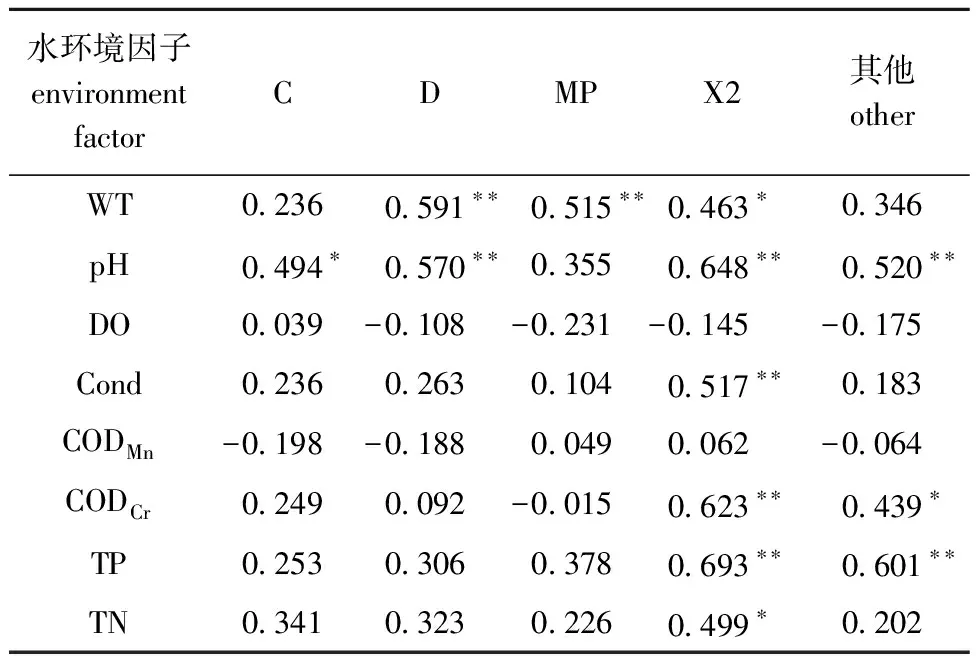
表4 浮游植物重要功能群生物量与水环境因子的Pearson相关性分析
2.3.2 RDA分析 从表5可见,特征值轴1大于轴2,前两轴物种数据累计变化率为66.4%,相关性累计变化率达到95.8%,表明浮游植物重要功能群与环境变量之间存在显著相关性。从图3可见,轴1最主要的正相关因子为pH(相关性系数为0.768 4),其余主要的相关性因子分别为TP(相关性系数为0.581 0)、TN(相关性系数为0.576 8)、Cond(相关性系数为0.564 3)及WT(相关性系数为0.501 2);轴2最主要的正相关因子为CODMn(相关性系数为0.492 4),最主要的负相关因子为DO(相关性系数为-0.467 9)。
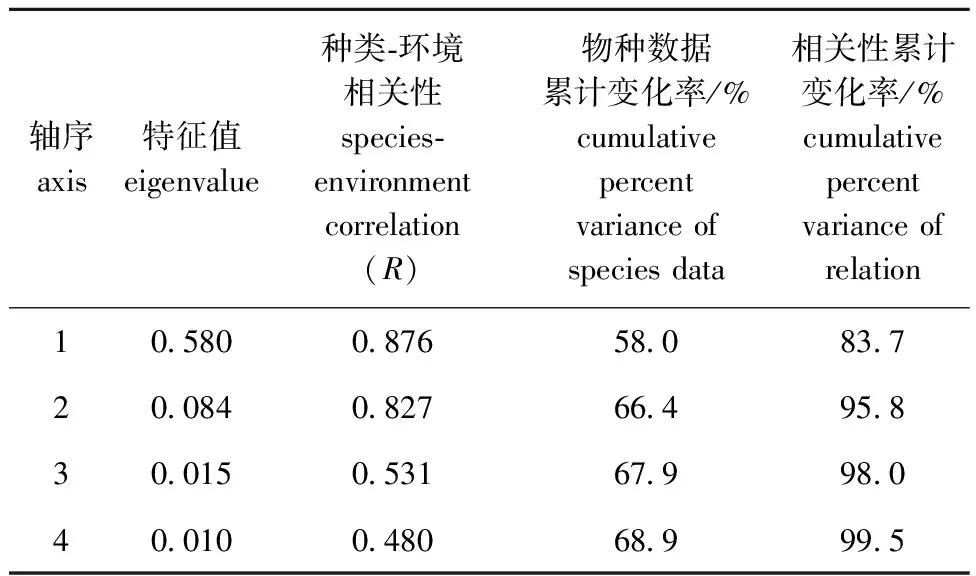
表5 排序轴特征值、浮游植物与水环境因子的相关性

图3 重要功能群与环境因子的RDA分析Fig.3 Redundancy analysis between the main functional groups and environment factors
重要功能群C、D、MP、X2与pH、TP、TN、Cond和WT均呈正相关关系;CODMn与除功能群C以外的其他功能群呈正相关关系,DO则相反,与除功能群C以外的功能群呈负相关关系(图3)。
3 讨论
3.1 海拉尔河浮游植物功能群特征
相较于湖泊,河流生态系统浮游植物的组成受温度和营养盐等因素影响相对较小,不同纬度、不同营养水平条件下优势种类均以硅藻为主,差异并不十分显著。洪松等[27]对20世纪70年代末至80年代前期的黑龙江、松花江、黄河、长江和珠江等中国南北方主要河流的相关研究显示,浮游植物组成以硅藻门和绿藻门为主,而优势种基本以硅藻门种类为主,主要为功能群C中的小环藻属,功能群D中的针杆藻属、菱形藻属,功能群MP中的舟形藻属、双菱藻属,以及功能群P中的直链藻属、脆杆藻属。从近年研究也可看出,不同纬度富营养化河流和贫营养河流重要功能群的差异均较小。亚热带地区富营养化河流[6]全年以功能群P、D占优,贫营养河流[17]全年则以C、MP、D占优;温带地区富营养河流[13]春、秋季节以功能群MP、D、L0、J占优;北方寒区富营养河流[16]全年以功能群C、D、MP占优,贫营养河流[17]春季以L0、MP、P、C占优。从本研究结果来看,北方寒区富营养化偏碱性河流海拉尔河浮游植物重要功能群为C、D、MP、X2,且呈现C+X2+D(春季)→X2+MP+D+C(夏季)→D+C+MP(秋季)的季节变化特征,功能群C、D和MP同样占重要地位。表明海拉尔河浮游植物功能群生物量季节变化较小。
河流系统浮游植物功能群生物量时空差异较大,通常下游水域浮游植物功能群生物量高于中、上游水域。杜红春等[6]对汉江中、下游水域浮游植物功能群生物量研究表明,下游水域生物量明显高于中游,下游功能群C和P显著增加。白海峰等[14]研究也表明,渭河流域浮游植物功能群生物量呈现下游>中游>上游的趋势。从本研究结果来看,海拉尔河水域浮游植物功能群生物量也呈现下游>中游>上游的趋势,下游功能群C也显著增加。
功能群C、X2是海拉尔河最具有代表性的功能群。功能群C代表中营养到富营养的中小型水体,海拉尔河春季进入融雪期,冰雪融化冲刷泥沙携带腐殖质入河,水体营养程度逐渐增大的同时,水体浑浊程度也增大,同时也验证了功能群C耐低光照的特性。这是由于海拉尔河发源于大兴安岭山间溪流,上游无人为污染干扰,而中游经过牙克石、海拉尔等人类活动密集区域,城市生活污水和工业废水等导致水体污染加剧[20-21],下游浮游植物大量生长,致使在下游区域的功能群C占据显著优势。功能群X2代表中营养到富营养化的浅水水体,且功能群X2以喜高温的绿藻门种类为主,使其在高温和高营养盐的夏季占据优势。功能群D和MP适宜生活在频繁扰动的浑浊水体中,夏季雨水增多,雨水带来的扰动使更适应此类环境的功能群D、MP占据优势,同时由于功能群D具有喜光的特性,使其在水质更好、更清澈的上游区域占据优势。
3.2 海拉尔河浮游植物功能群与水环境因子的关系
浮游植物功能群的演替同水温、营养盐、光照强度、水动力学,以及水生生物之间的相互作用联系十分紧密,在不同环境条件及不同水环境因子下,耐受特定环境的浮游植物群体会在其适宜条件下生存下来并占据主导位置[25]。pH对海拉尔河浮游植物功能群产生着重要的影响。海拉尔河pH为7.5~8.5,属于弱碱性水体。有研究显示,水体的酸碱性会影响浮游植物的生长,当水体处于弱碱性时,会加快浮游植物吸收二氧化碳,促进浮游植物的生长[28]。本研究中,海拉尔河重要功能群C、D、MP、X2均与pH呈正相关关系,说明在适宜范围内,pH会促进这些功能群浮游植物的生长。这与于洪贤等[18]对春季牡丹江浮游植物研究的结果相似。
富营养化水体中,营养盐也是影响浮游植物功能群的主要因子[5],营养盐的浓度与占比均会影响浮游植物的生长与活动。安睿等[29]对小兴凯湖浮游植物功能群研究表明,TP是影响浮游植物功能群的最主要影响因子之一;李晓钰等[19]对松花江哈尔滨段浮游植物功能群研究表明,CODCr、WT和TP是影响浮游植物功能群的最主要影响因子。对不同区域、不同类型的富营养化水体研究均表明,TN、TP是影响浮游植物功能群的主要因素,尤其是对淡水生态系统,相较于TN,TP对浮游植物功能群的影响更大[30]。于孝坤等[15]对芜湖河道浮游植物研究表明,功能群C与TN、TP均呈正相关关系,相较于TP,功能群C与TN相关性较大;陈楠等[31]对泰湖浮游植物的研究也证明了功能群X2与TN、TP呈显著正相关关系;对长寿湖[8]和新丰江水库[9]的研究均表明,功能群D和MP与TN、TP也呈正相关关系。与上述研究结果一致,本研究表明,TN、TP均是影响海拉尔河流域浮游植物重要功能群的主要相关因子,其中,TP对浮游植物功能群的影响更大。海拉尔河浮游植物重要功能群C、X2、D和MP均与TN、TP呈正相关,其中,功能群X2和D相比其余功能群与TN、TP的相关性较大,功能群MP与TP相关性较大,而功能群C则与TN相关性较大。由此可见,TN、TP明显能促进重要功能群浮游植物的生长。
WT同样是影响海拉尔河浮游植物功能群的重要因子之一。WT是通过控制光合作用的酶促反应或呼吸作用的强度来直接影响藻类的生长[32],并通过影响藻类酶的活性间接影响其对光能和营养盐的获取、利用,进而影响藻类的生长[33]。相对于热带、亚热带区域,WT通常是受温度变化影响较为明显的寒区浮游植物功能群的主要因素之一。林岿璇等[34]对镜泊湖浮游植物研究表明,WT是影响寒区浮游植物功能群的主要影响因子。张俊芳等[10]对汤浦水库研究表明,WT与功能群X2呈显著正相关;韩丽彬等[11]对贵州百花水库浮游植物研究表明,功能群D与WT呈显著正相关;杨雅兰等[12]对云南大理洱源西湖浮游植物研究表明,功能群MP与WT呈正相关关系,但不是主要正相关因子。可以看出,WT对不同浮游植物功能群的影响程度也不同,功能群X2和D通常与WT呈显著正相关,功能群MP和C虽然也与WT呈正相关,但相关性不显著。与此结果相似,本研究表明,功能群X2、D、MP和C均与WT呈正相关,尤其是功能群X2、D、MP与WT呈显著正相关。这是由于功能群X2主要以绿藻门的衣藻属种类为主,衣藻属种类适宜生长温度为25 ℃[35],其在WT较高的夏季占比较高,而在WT较低的春、秋季节占比显著下降。而以硅藻门为主的功能群更适宜低温环境,且最适生长温度很少超过25 ℃[36]。针杆藻属既能在冬、春等低温季节生长[37],同时也能在夏季快速生长,说明针杆藻属是对水温适应较广的种类。由此可见,以针杆藻属为主的功能群D与WT会呈现显著的正相关关系。
4 结论
1)海拉尔河浮游植物重要功能群组成季节变化呈现C+X2+D(春季)→X2+MP+D+C(夏季)→D+C+MP(秋季),浮游植物生物量空间变化呈现下游>中游>上游的趋势。说明海拉尔河浮游植物功能群组成较稳定,受时间变化影响较小,浮游植物生物量在人类活动强度的影响下,空间变化较大。
2)浮游植物功能群C在下游比例显著升高,说明海拉尔河下游污染相较上游及中游较重。
3)影响海拉尔河流域浮游植物功能群的主要水环境因子为pH、TP、TN和WT,均与功能群C、D、MP、X2呈正相关,尤其是功能群X2与pH、TP、TN和WT呈显著正相关,D与WT、pH呈显著正相关,功能群MP相较于TN,与TP相关性更大,功能群 C则与TN相关性更大。说明控制海拉尔河污染应着重于N、P的点源及面源污染,改善下游工业、生活及牧业的废水污水排放处理体系,逐步减轻对海拉尔河水域的危害,进而维护整个呼伦湖流域的生态安全。
猜你喜欢
数学物理学报(2021年4期)2021-08-30
中老年保健(2021年12期)2021-08-24
中国传媒大学学报(自然科学版)(2021年1期)2021-06-09
今日农业(2020年19期)2020-12-14
中等数学(2020年1期)2020-08-24
中国生殖健康(2020年6期)2020-02-01
文化创新比较研究(2020年8期)2020-01-02
中国生殖健康(2019年11期)2019-01-07
特别健康(2018年3期)2018-07-04
中国科技信息(2015年2期)2015-11-16
