
外源硫对Cd胁迫下番茄植株生长及抗逆代谢的影响
2024-01-03 13:12贾红磊张春平吴广放
陕西科技大学学报 2024年1期
贾红磊, 王 晴, 陈 翠, 魏 婷, 张春平, 吴广放
(1.陕西科技大学 环境科学与工程学院, 陕西 西安 710021; 2.中国建筑材料工业建设西安工程有限公司, 陕西 西安 710061)
0 引言
由于对有限矿产资源的探测和大力开发、工业生产制造、施用化肥农药和污水灌溉等一系列人为因素,使得大量有害重金属被排放到自然环境中,造成严重的环境污染问题[1],其中受到重金属污染的农田土壤正面临土地退化、粮食减产的问题.而重金属污染问题中,镉(Cd)污染问题尤为突出[2].调查发现,广东、江西、浙江、湖南、河北、陕西等地区土壤均受到轻重不一的Cd污染[3].Cd由于可以取代酶活性位点中起关键作用的基本元素及对巯基的高亲和力,被认为是强毒性物质[4].Cd通过根系进入植物体扰乱植物正常生理和代谢,如降低植物生物量[5]、破坏叶绿体结构,降低植物光合作用[6]、抑制土壤中营养物质的吸收转运等[7].植物自身具有响应Cd胁迫的应答机制,例如增加可溶性蛋白含量对生物膜和维持细胞内环境稳态起到保护作用,调节抗氧化系统提高抗氧化能力,改变细胞壁组成及结构固定Cd等[8,9].
硫(S)作为植物必须的大量元素,是某些氨基酸(半胱氨酸、甲硫氨酸)、抗氧化剂(还原型谷胱甘肽)、辅酶、维生素、植物螯合素和脂质的组成部分,这些物质在调节植物生长发育、响应非生物胁迫等方面发挥着关键作用[4,10].此外,S还以硫化氢(H2S)的形式存在于植物体内,与NO、CO和生长素相互作用调节根的形成,缓解各种非生物胁迫对根生长的抑制[11,12].植物根际通过吸收土壤中的硫酸盐完成自身对S的同化过程,并激活植物根部的硫酸盐转运蛋白(SUT)使该过程持续进行[13].在不同的含S化合物中,半胱氨酸(Cysteine,Cys)和谷胱甘肽(Glutataione,GSH)在缓解重金属诱导的氧化应激中起着重要作用[14].S可以通过维持高水平的植物络合素(Phytochelatin,PCs)或其前体GSH来限制重金属向地上部的转运.Khan等[15]发现外源S可以提高植物的新陈代谢,增加其生物量,降低Cd对植物产生的毒性.
氮(N)和磷(P)在调节植物营养平衡和生长发育过程中发挥重要作用,S、N和P同化途径可以在逆境条件下通过改善植物生理增强其抗逆性,它们的同化作用是各种生物合成途径的基础,是植株调节的重要手段[16].目前尚不清楚S是否可以通过调节植物N和P代谢提高植物响应Cd胁迫的能力.本研究旨在通过对植物体内N、P含量、存在形式、分布及代谢关键酶的研究,为外源S调控植物响应重金属胁迫提供理论依据.
1 实验部分
1.1 实验材料与设计
实验植物为番茄(石红三号,Solanum lycopersicum L.),种子由新疆石河子蔬菜研究所番茄研究开发中心提供.将适量的种子盛于干净的烧杯中,用3%的次氯酸钠浸泡30 min后,用去离子水清洗干净完成消毒.将消毒后的种子均匀播种(每穴5~6颗)在装有蛭石的穴盘中,保证蛭石湿度便于种子的萌发.穴盘避光放置于人工气候室,待种子发芽长至约4~5 cm,用泡沫棉包裹植株茎部转移至含有1/4 Hoagland的水培箱培养两周,替换为浓度为1/2的营养液,继续培养2周.人工气候室设置:光照周期16 h/8 h(光照/黑暗),相对湿度40%~60%,温度26±2 ℃(昼)/20±2 ℃(夜).
番茄幼苗培养至两叶一心时即可准备处理,选取生长良好且一致的幼苗,移至不同的培养罐中处理,每个培养罐中移植4株番茄幼苗,根据实验需求收样测定相关指标.处理组中缺S营养液(0 mmol·L-1S)即为将营养液中的27.80 mg·L-1FeSO4换为73.50 mg·L-1FeCl2·4H2O,其余不变;外源S以Na2SO4的形式加入.
本试验Cd浓度为3 mg·L-1,试验共设置8组处理,分别为0 mmol·L-1S(缺S)、1 mmol·L-1S(对照)、2 mmol·L-1S、4 mmol·L-1S、0 mmol·L-1S+Cd(胁迫缺S)、1 mmol·L-1S+Cd(胁迫对照)、2 mmol·L-1S+Cd、4 mmol·L-1S+Cd.
1.2 实验方法
1.2.1 脯氨酸和可溶性蛋白含量的测定
采用茚三酮法测定脯氨酸含量[16],0.50 g待测样品加入5 mL磺基水杨酸溶液,沸水浴浸提10 min,过滤浸提液,显色反应后在520 nm波长处测定,根据标准曲线计算脯氨酸含量.
利用考马斯亮蓝G-250法测定可溶性蛋白含量.称取0.50 g鲜样加入少量蒸馏水研磨,直至呈现匀浆状态倒入10 mL离心管,用6 mL蒸馏水反复冲洗,洗涤液也一并收集在离心管中,4 000 r/min离心10 min后取出定容得到提取液.显色后在595 nm波长下测定并根据标准曲线计算其含量[16].
1.2.2 氮(N)代谢相关指标的测定
硝酸还原酶(NR)活性测定根据北京索莱宝科技有限公司的NR活性检测试剂盒(Griess显色法)进行.取植物鲜样0.2 g于预冷的NR Lysis缓冲液中,冰浴研磨匀浆、离心,留取上清液即为硝酸还原酶粗提液(4 ℃保存待用).取适量的亚硝态氮标准样制备标准曲线,利用酶标仪检测计算活性.其单位为μg·g-1·h-1
谷氨酰胺合成酶(GS)活性的测定参照Lillo C[19]的方法.称取植物鲜样0.2 g于3 mL提取缓冲液(pH 8.0,0.05 mol·L-1Tris-HCL)中,冰浴上研磨匀浆,4 ℃,15 000 g离心20 min,上清液即为粗酶液.取1.6 mL反应混合液B(pH 7.4,含盐酸羟胺),加入0.7 mL粗酶液和40 mmol·L-1ATP溶液,混匀,于37 ℃下保温半小时,加入显色剂1 mL,5 000 g离心10 min,取上清液在540 nm处测定吸光值.单位为A·mg-1protein·h-1.
1.2.3 磷(P)代谢相关指标的测定
总磷(TP)含量的测定参照现有方法并稍作修改[20-22].称取植物干样0.25 g(精确至0.001 g)加入6 mL HNO3浸泡1 h后加入2 mL H2O2.置于电热消解仪上按照如下程序消解:120 ℃-10 min,150 ℃-15 min,190 ℃-25 min,冷却后取出,100 ℃-30 min后取出冷却并定容,利用电感耦合等离子体质谱仪(Inductively Coupled Plasma Mass Spectrometer,ICP-MS)进行测定.
醋酸钠溶液研磨浸提后得到含酸性磷酸酶(Acid Phosphatase,APA)提取液,过滤后取1 mL置于试管中,加入0.05 mol·L-1对硝基苯磷酸二钠溶液2 mL,100 ℃条件下反应10 min后依次加入2 mL CaCl2和NaOH溶液终止反应.离心后测定吸光值,根据标准曲线计算其活性[20].
2 结果与讨论
2.1 外源硫对Cd胁迫下番茄植株生物量累积的影响
为了确定外源S对番茄植株生长的调节作用,分析了不同浓度S在-Cd或+Cd条件下每个培养罐中番茄植株指定位置的叶片鲜重.结果如图1(a)所示,无Cd情况下,与0 mmol·L-1(缺S组)相比1 mmol·L-1S,2 mmol·L-1S与4 mmol·L-1S均显著提升番茄植株地上、地下鲜重,与1 mmol·L-1(对照组)相比2 mmol·L-1S显著提升番茄植株地上、地下鲜重(P<0.05),但4 mmol·L-1S处理与对照组无显著性差异,缺S组生物量最低,这说明Cd显著抑制植物的生长,外源S明显缓解由于Cd胁迫导致的生物量下降(P<0.05).

图1 外源硫对Cd胁迫下番茄植株生长的影响
S对根生物量累积的影响与地上部相似,2 mmol·L-1S处理具有较好的调控效果(如图1(b)所示).在-Cd情况下,2 mmol·L-1S处理生物量累积分别是缺S和对照组的2.1倍和1.25倍(P<0.05).Cd胁迫导致0 mmol·L-1S和1 mmol·L-1S组别中生物量下降32%~36%,S浓度为2 mmol·L-1时生物量提升42%(P<0.05).结果表明,S具有显著增加植物生物量的效果且能够缓解由于Cd对番茄植株造成的生物量减少效应.
2.2 外源硫对Cd胁迫下番茄植株抗逆代谢物的影响
脯氨酸和可溶性蛋白伴随植物N代谢生成,是一种调节植物渗透势的大分子物质,是植物体内响应逆境胁迫抵抗脂质过氧化等过程的关键调节物[23,24].此外,可溶性蛋白也是植物响应胁迫的重要代谢物[25].为评价外源S对番茄植株抗逆代谢物质的调节作用,分别在无Cd和有Cd胁迫条件下研究不同浓度S处理下番茄植株对脯氨酸、可溶性蛋白的积累.
2.2.1 外源硫对Cd胁迫下番茄植株体内脯氨酸含量的影响
由图2(a)可知,与对照组相比,外源S显著提升番茄根中脯氨酸含量.S浓度为2 mmol·L-1时,根中脯氨酸含量是对照组的1.28倍.Cd胁迫时,根中脯氨酸含量比对照增加39.58%,2 mmol·L-1+Cd处理组是1 mmol·L-1+Cd(胁迫对照组)的1.20倍,0 mmol·L-1+Cd(胁迫缺S组)的1.8倍(P<0.05).表明Cd胁迫导致番茄根中脯氨酸累积,外源S的添加诱导根中脯氨酸含量上升响应Cd胁迫.
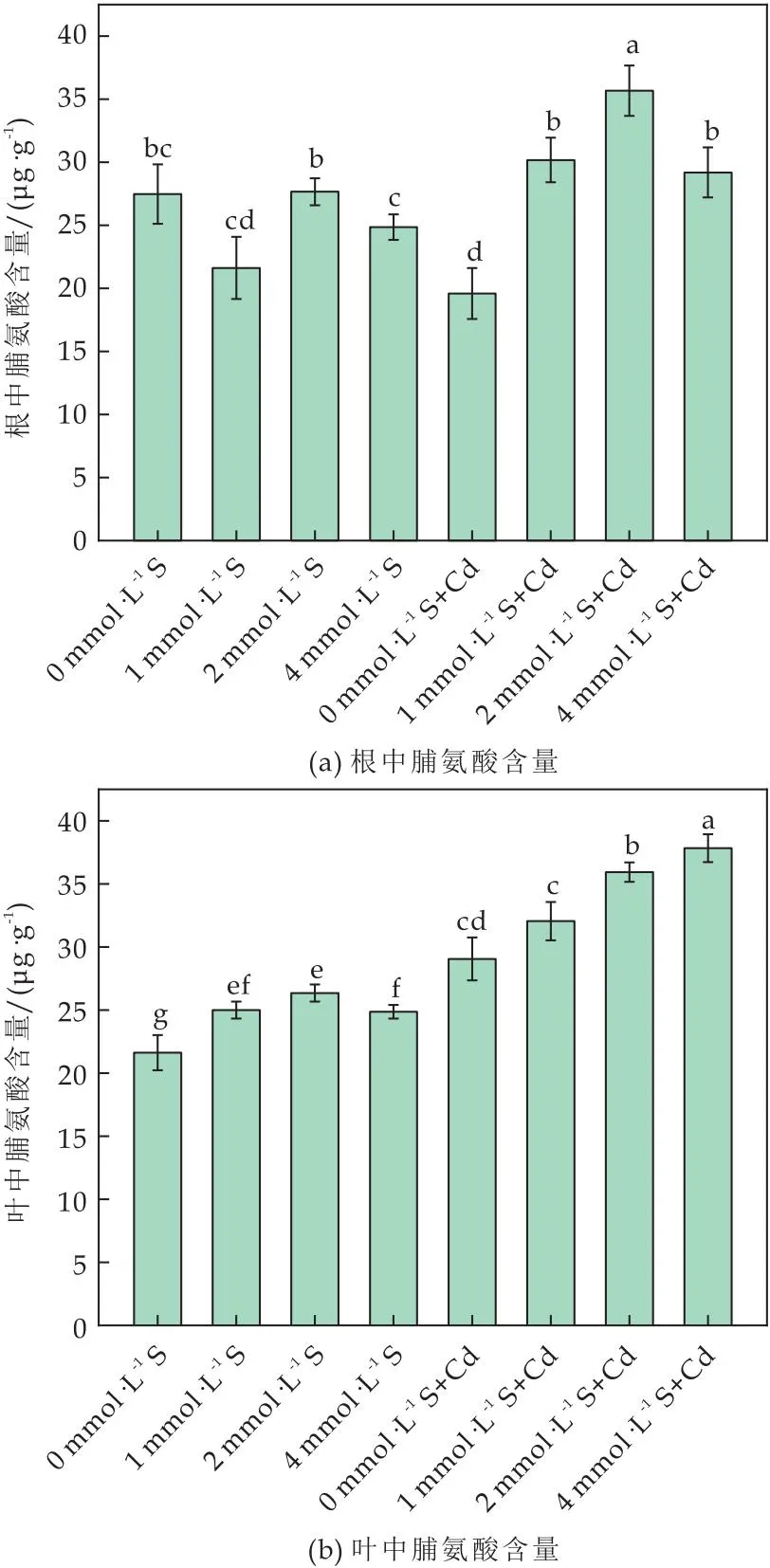
图2 外源硫对Cd胁迫下番茄植株脯氨酸含量的影响
叶中脯氨酸含量如图2(b)所示,单独S处理时,脯氨酸含量呈现小幅度的剂量依赖变化.Cd胁迫下,叶中脯氨酸含量随S浓度呈剂量依赖性提升.S浓度为2 mmol·L-1和4 mmol·L-1时叶中脯氨酸含量分别是胁迫对照组的1.12、1.18倍(P<0.05).表明外源S可以促进Cd胁迫下脯氨酸的累积响应Cd胁迫.脯氨酸含量增加可视为S调控番茄植株对Cd胁迫响应的信号,可能是其抵御Cd毒害的重要调控途径之一[26].
2.2.2 外源硫对Cd胁迫下番茄植株体内可溶性蛋白含量的影响
在植物中,可溶性蛋白是渗透调节剂和活性氧清除剂,因此可以减少重金属对植物细胞的损害[27].番茄植株可溶性蛋白变化如图3所示,在无Cd条件下,外源施加2 mmol·L-1和4 mmol·L-1S时,根中可溶性蛋白量分别是对照组的2.87、2.30倍.Cd胁迫条件下,番茄根中可溶性蛋白累积量随着S剂量增大同步增加(P<0.05).当S浓度为2 mmol·L-1和4 mmol·L-1时,番茄的根中可溶性蛋白含量是胁迫对照组的1.39、1.65倍(如图3(a)所示).叶中可溶性蛋白含量如图3(b)所示,与单独Cd处理相比,2 mmol·L-1+Cd和4 mmol·L-1+Cd处理番茄植株叶中可溶性蛋白含量增加21.23%和9.41%(P<0.05).由此可见,Cd胁迫条件下,外源施加S能够有效增加番茄叶片中可溶性蛋白含量,缓解Cd对植株带来的危害.
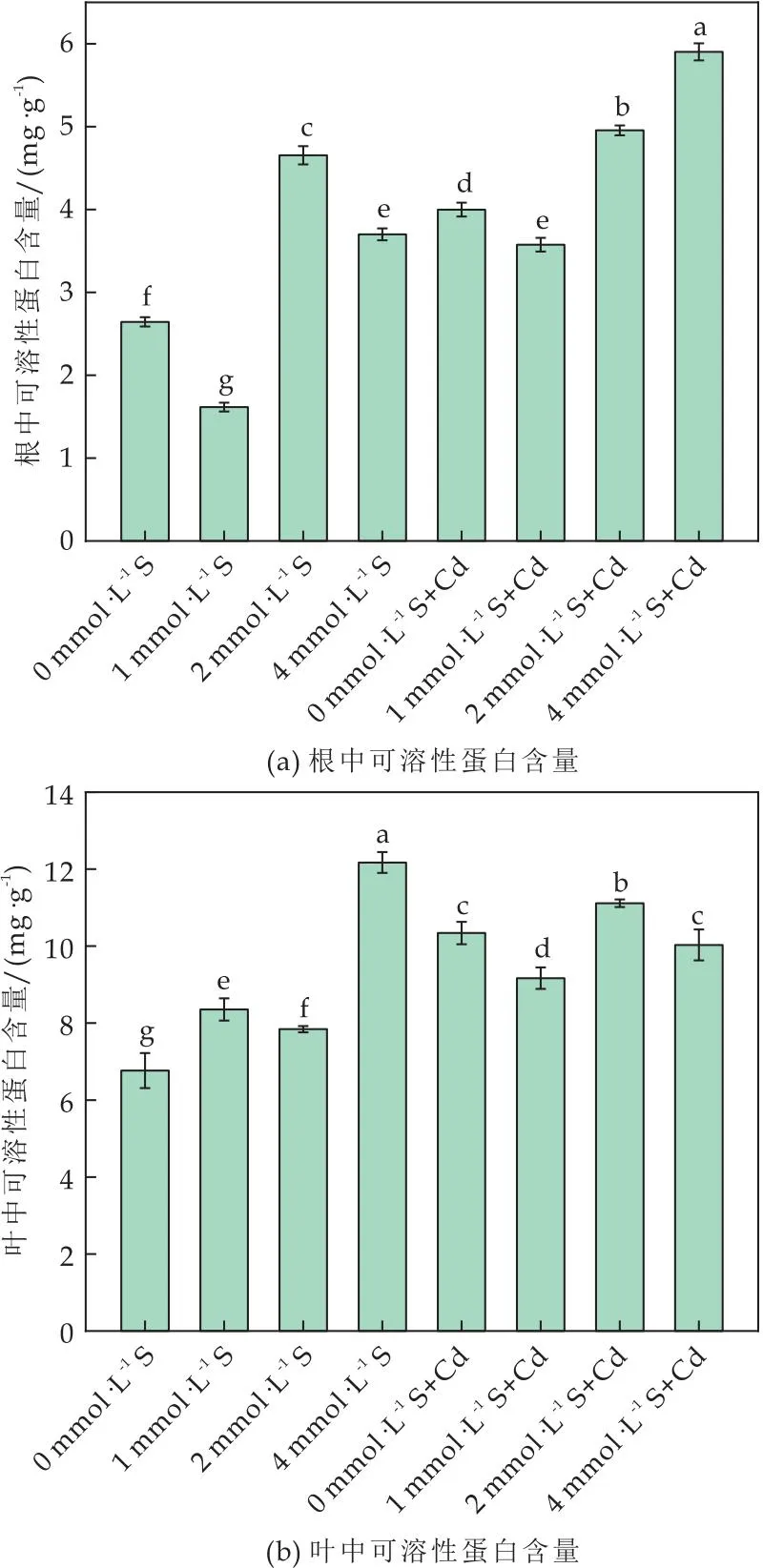
图3 外源硫对Cd胁迫下番茄植株可溶性蛋白含量的影响
综上所述,Cd胁迫诱导番茄植株可溶性蛋白含量增加可归因于番茄植株的自我平衡机制,缓解Cd对植物细胞膜结构的损伤所导致的渗透电位失衡问题[25,27].而外源S使植株体内可溶性蛋白含量进一步累积,降低植株的Cd毒性效应,从而提高番茄植株对Cd胁迫的抗性.
2.3 外源硫对Cd胁迫下番茄植株氮代谢的影响
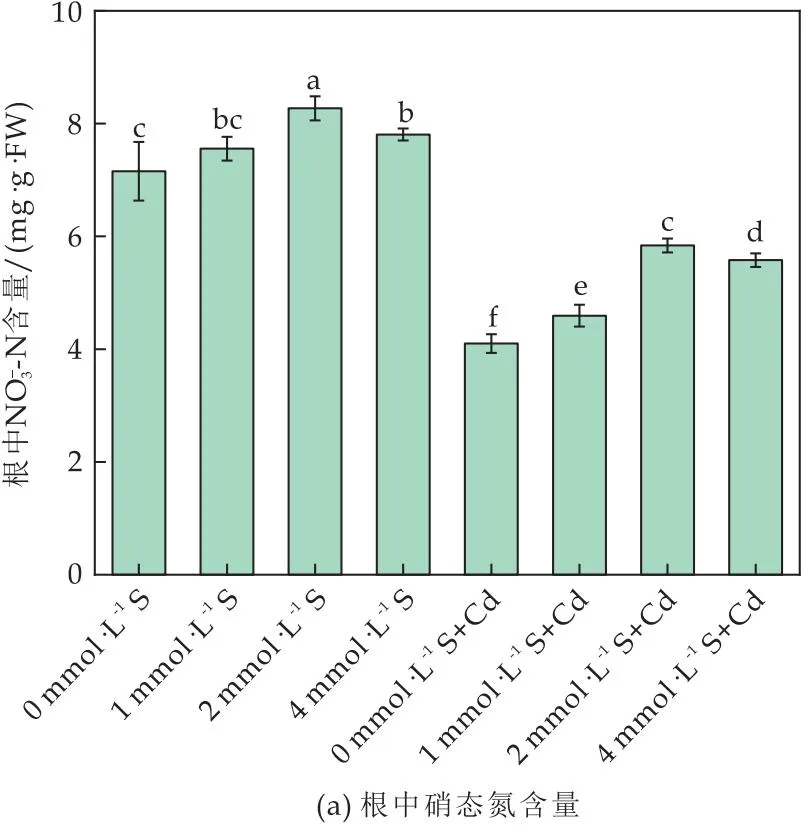
图4 外源S对Cd胁迫下番茄植株和的影响
2.3.3 外源硫对Cd胁迫下番茄植株N代谢关键酶活性的影响
在本实验中,NR活性受Cd和S的影响显著(如图5(a)、(b)所示).单独S处理条件下,随着S浓度的增加,根系中NR活性上升(P<0.05),但在叶片中含S条件下NR活性无显著变化.Cd胁迫下,番茄植株根和叶中NR活性均明显下降,外源S能在一定程度上缓解该抑制作用,2 mmol·L-1S处理时植株根和叶中NR活性分别是胁迫对照组的1.16、1.06倍;S浓度为4 mmol·L-1时无显著变化(如图5(a)、(b)所示).结果表明,外源S对Cd胁迫下的NR活性有一定的激活作用,2 mmol·L-1时的效果最为显著,且在根中作用效果优于叶片.
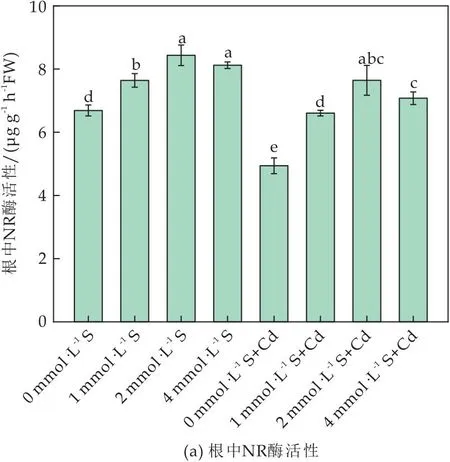
图5 外源S对Cd胁迫下番茄植株NR酶和GS酶活性的影响
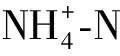
2.4 外源硫对Cd胁迫下番茄植株磷代谢的影响
P参与许多生理过程,如光合作用和碳水化合物的生产,增强应激环境中抗氧化防御系统的活动[34,35].为探索外源S在Cd胁迫条件下对植株P代谢的调节作用,在无胁迫和Cd胁迫条件下添加不同浓度S分析番茄植株P含量和酸性磷酸酶(APA)活性变化.
2.4.1 外源硫对Cd胁迫下番茄植株P含量的影响
在无Cd胁迫条件下,随着S浓度的增加根系中P的含量上升,叶片中磷含量下降;Cd胁迫下,番茄根中P含量显著下降,叶中P含量上升(如图6所示).
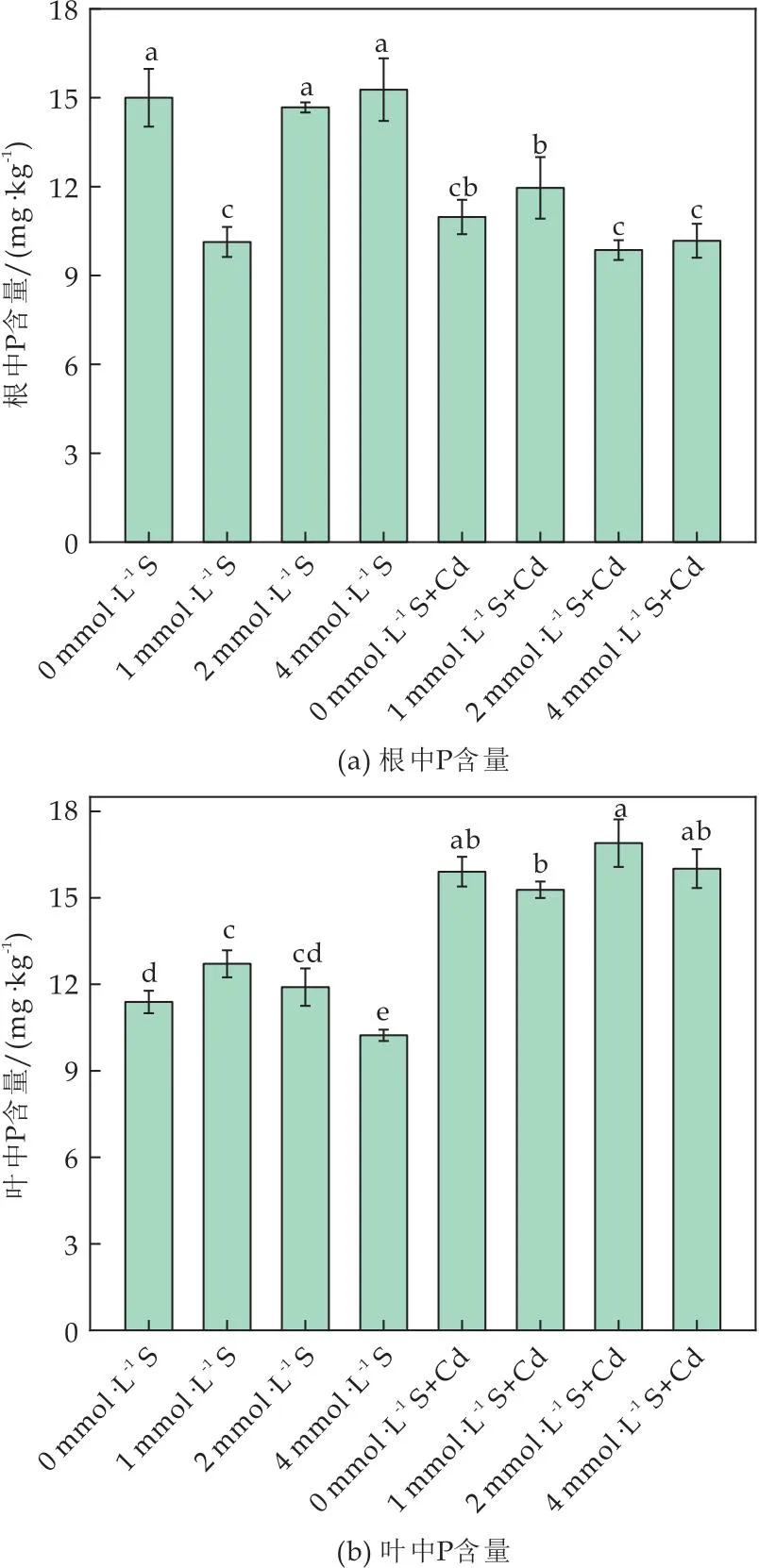
图6 外源硫对Cd胁迫下番茄植株P含量的影响
表明叶中可能需要更多的P维持Cd胁迫下植物的代谢,因此S促进P从根中转移到叶中.Cd进入植物后,P可以通过促进液泡中不溶性Cd-磷酸络合物的形成和细胞壁中果胶酸盐/蛋白质螯合Cd的形式来降低Cd的迁移率[35].因此在Cd胁迫下,植物体内P可能一部分与Cd形成Cd-磷酸络合物,使植物体内可利用P缺乏.外源S处理后可能促进番茄P元素由根向叶中转移,一方面加强对Cd的螯合,另一方面维持自身代谢以缓解Cd胁迫.在缺硫情况下,植物可能通过抗逆响应调节或影响P摄取相关转录因子的表达来提高植物对磷的摄取量[36-40].
2.4.2 外源硫对Cd胁迫下番茄植株P代谢关键酶活性的影响
APA是植物体内的一种诱导酶,可以水解有机磷,APA活性的增加与许多物种和植物部分的低水平Pi相关[41].无Cd条件下,S诱导APA活性上升,但在Cd胁迫时根和叶中APA的变化趋势不同(如图7所示).
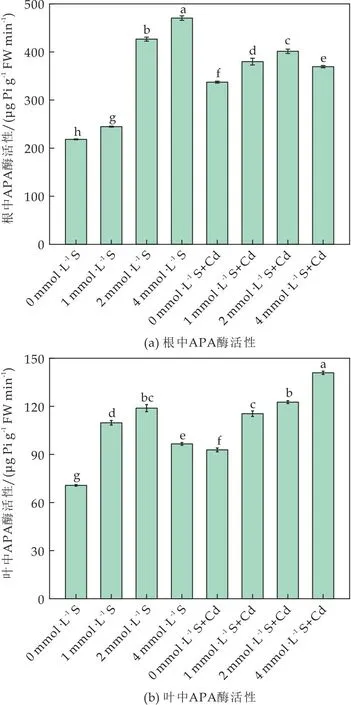
图7 外源硫对Cd胁迫下番茄植株APA酶活性的影响
在根中,与对照相比Cd处理后APA活性升高1.55倍(如图7(a)所示);在叶中,Cd胁迫下APA活性是对照的1.05倍(如图7(b)所示),可能由于Cd胁迫造成番茄体内Pi缺乏,导致APA酶活性上升水解有机磷维持体内Pi的需求[42].与Cd胁迫对照相比,S浓度为2 mmol·L-1和4 mmol·L-1时,叶中APA活性是其1.06、1.22倍(P<0.05).
3 结论
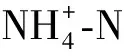
猜你喜欢
中老年保健(2022年3期)2022-11-21
舰船科学技术(2022年11期)2022-07-15
西藏农业科技(2019年3期)2019-11-04
科学与财富(2019年15期)2019-10-21
发酵科技通讯(2018年2期)2018-07-06
现代园艺(2018年3期)2018-02-10
上海农业学报(2017年3期)2017-04-10
湖南林业科技(2017年1期)2017-02-06
中成药(2016年4期)2016-05-17
现代食品(2016年14期)2016-04-28
