
葡萄酒发酵香气物质调控研究进展
2024-01-26 08:09徐超胡文效陈明光朱明宣邱磊赵先炎
中外葡萄与葡萄酒 2024年1期
徐超,胡文效*,陈明光,朱明宣,邱磊,赵先炎
(齐鲁工业大学(山东省科学院)/生物基材料与绿色造纸国家重点实验室,山东济南 250353)
葡萄酒的香气物质按来源分为品种香、发酵香和陈酿香。葡萄酒香气的香韵和香气强度特征综合反映着葡萄酒原料的品种特性、产地风土特点和葡萄酒发酵、陈酿方法及技术水平。其中,葡萄酒发酵环节是彰显葡萄酒品种香、形成葡萄酒发酵香、影响葡萄酒陈酿香的关键环节。葡萄酒发酵本质是借助微生物将葡萄汁成分转变为葡萄酒成分的系列生物化学反应,微生物(主要是酵母菌、乳酸菌)在发酵过程中的作用显著影响葡萄酒的“表型”,操纵葡萄酒的风味、香气和风格[1]。基于生态多样性、微生物物种多样性和遗传多样性,以及高通量测序技术和气相色谱-质谱联用分析技术(GC-MS)的发展,葡萄酒发酵微生物群落组成及演替与葡萄酒香气相关性[2-4]逐渐被揭示出来;基于酶调节作用和转录组、蛋白组表达结果,通过调控葡萄酒发酵微生物菌群或香气化合物生物合成途径关键酶合成与酶活性水平,使实现调控目标代谢物的代谢通量成为可能。
1 葡萄酒发酵与葡萄酒香气的关系
葡萄向葡萄酒的转化涉及真菌(酵母菌)、细菌(乳酸菌、醋酸菌)等多种微生物的协同作用,这些微生物形成了一个复杂的生态系统,它们的代谢活动独立进行又相互作用,共同塑造了葡萄酒的风味风格。微生物代谢产生的高级醇、酸、酯类等构成葡萄酒发酵香的物质基础,发酵过程中酶催化葡萄原料中的香气前体物释放出C13降异戊二烯类化合物、游离态的萜烯、硫醇类等物质,对于增强葡萄酒品种香气和产区风味特色具有极其重要的意义。葡萄酒中被检出的香气物质有800余种[5],这些香气物质通过协同、加成或掩盖等作用,形成了葡萄酒丰富而独特的香韵结构,并决定了葡萄酒的风格特点。
1.1 葡萄酒发酵作用增强品种香
葡萄品种香气物质主要包括萜烯类、降异戊二烯类、芳香族类、脂肪族类、甲氧基吡嗪类和挥发性含硫化合物等,这些化合物多以香气前体物的形式存在于葡萄果实中,在发酵过程中香气前体物在酶促(糖苷酶、酯酶、C-S裂解酶等)和非酶促作用(酸催化水解和化学重排)下,进一步转化为挥发性化合物,香气物质被释放出来,对葡萄酒风味的感官感知有积极的促进作用[6]。
1.1.1 萜烯类香气物质
萜烯是分子骨架以异戊二烯(C5单元)为基本结构单元的化合物及其衍生物,其分子式符合(C5H8)n通式,目前在酿酒葡萄果实中已经发现了70多种萜烯类物质[7],其中单萜是果实中含量最高、种类最为丰富的萜类物质,常见的单萜包括芳樟醇、香叶醇、香茅醇、橙花醇等。
葡萄浆果中萜烯类物质通过2-甲基-D-赤藓糖醇-4-磷酸(MEP)途径合成[8],主要存在于果皮中,且糖苷结合态萜烯的含量高于游离态。例如在葡萄牙不同产区的两种白葡萄品种中,糖苷类香气物质的结合态萜烯分别占总香气物质的54%和67%[9]。葡萄果实中萜烯类物质的萜烯苷元主要是单萜醇、单萜二醇和单萜三醇[10],而糖苷的主要形式是双糖苷,占总糖苷的80%以上,双糖苷类香气前体物质的主要酶解方式如图1所示[11]。
通常在葡萄酒发酵环境条件下,葡萄原料中的糖苷类化合物在糖苷酶作用下部分被水解[12-13],但一般糖苷酶的最适条件是pH5.0~6.0、温度40~50 ℃,在葡萄酒发酵实际环境(高糖、低pH、低温、高乙醇)下糖苷酶活性受到抑制,导致其活性仅为最适条件下的10%左右[14],因此,葡萄原料糖苷结合态香气化合物的有效释放成为葡萄酒发酵品种香调控的关注点之一。
1.1.2 降异戊二烯类
降异戊二烯类化合物是一类由类胡萝卜素(四萜)如β-胡萝卜素等降解产生的13个、11个、10个、9个碳原子的衍生物。目前有关葡萄与葡萄酒降异戊二烯化合物的研究多集中于C13类,C13降异戊二烯类化合物及其衍生物,如α-紫罗兰酮、β-紫罗兰酮、β-大马士酮、香叶基丙酮等,虽然这类化合物在葡萄果实中含量较低,但因其具有阈值低(如β-紫罗兰酮阈值0.007 μg·kg-1[15],β-大马士酮阈值0.002 μg·kg-1[16])、香气活度(OAV)值较高的特点,对葡萄酒品种香气具有重要贡献。
类胡萝卜素双加氧裂解酶(Carotenoid cleavage dioxygenase,CCDs)是参与类胡萝卜素裂解产生香气化合物的主要酶,植物CCD亚基因家族中CCD1、CCD4与香气物质的形成有关,在葡萄中发现并鉴定了3个参与类胡萝卜素裂解并产生香气化合物的基因VvCCD1、VvCCD4a和VvCCD4b[17],近来才发现异戊二烯路径的首个负调控因子VvWRKY70[18]。β-胡萝卜素在CCD1和CCD4作用下能在9,10和9',10'双键裂解产生α-紫罗兰酮和β-紫罗兰酮等[19]。在葡萄酒酿造过程中,葡萄中类胡萝卜素可以转化为游离态β-大马士酮、β-紫罗兰酮、葡萄螺烷、雷司令缩醛、三甲基二氢萘(TDN)等[12],成为葡萄酒发酵品种香调控的又一关注点。
1.1.3 硫醇类
大部分挥发性硫化物对葡萄酒香气产生消极影响,但部分硫醇如4-巯基-4-甲基戊烷-2-酮(4MMP)、3-巯基己醇(3MH)、3-巯基己醇乙酸酯(3MHA)等表现出生动的果香(番石榴、西番莲),它们是由酵母分泌的C-S裂解酶催化,从半胱氨酸和谷胱甘肽结合的前体中释放[20-21],有研究表明,3MH可以通过酿酒酵母转化为3MHA[22]。这些硫醇及其前体物质广泛存在于白葡萄品种的葡萄酒中,如‘长相思’‘雷司令’‘灰比诺’‘霞多丽’等,其中4MMP在‘长相思’葡萄酒中含量一般每千升超过40纳克,而3MHA和3MH每千升分别可达几百纳克和几百到几千纳克[23-24]。有报道显示,在发酵过程中只有很少部分的芳香硫醇被释放出来[25-26],也有研究表明,只有3.2%的胱硫醚类香气前体物被裂解[27],这可能与酿酒酵母中胱硫醚β-裂解酶的活性较低有关。
品种香气还包括甲氧基吡嗪类、芳香族类和脂肪族如C6化合物等,其中甲氧基吡嗪是由氨基酸代谢中产生的含氮杂环物质,其较低的阈值赋予葡萄酒浓郁的青椒味。欧亚种酿酒葡萄及葡萄酒中其浓度约0.5~50 ng·L-1,强烈的青椒气味往往被认为对葡萄酒香气质量有消极影响。脂肪族C6化合物是由前体物质脂肪酸经脂氧合酶(LOX)途径逐步氧化、裂解、脱氢等作用生成,为葡萄酒带来青草味及生青味[28]。
1.2 发酵作用合成发酵香
葡萄酒发酵香香气化合物主要包括醇类、酯类和酸类化合物,属于微生物次生代谢产物,其中,乙酯、乙酸酯、高级醇对发酵香贡献较大,其代谢合成示意图如图2[29]。葡萄果皮自然携带酵母菌群,包括非酿酒酵母(non-Saccharomyces)和酿酒酵母(S.cerevisiae),并且非酿酒酵母占据种群优势(1×105~1×106cfu·mL-1),随着发酵的进行,非酿酒酵母的优势地位很快被酿酒酵母取代。酿酒酵母发酵力强,可以耐受更高的SO2和酒精度,在酒精发酵中占据主导地位(1×106~1×108cfu·mL-1)[30]。酿酒酵母在酒精发酵阶段可将葡萄糖转化为酒精、CO2及其它副产物,其代谢活动对葡萄酒的风味和质量有着重要的影响[31]。
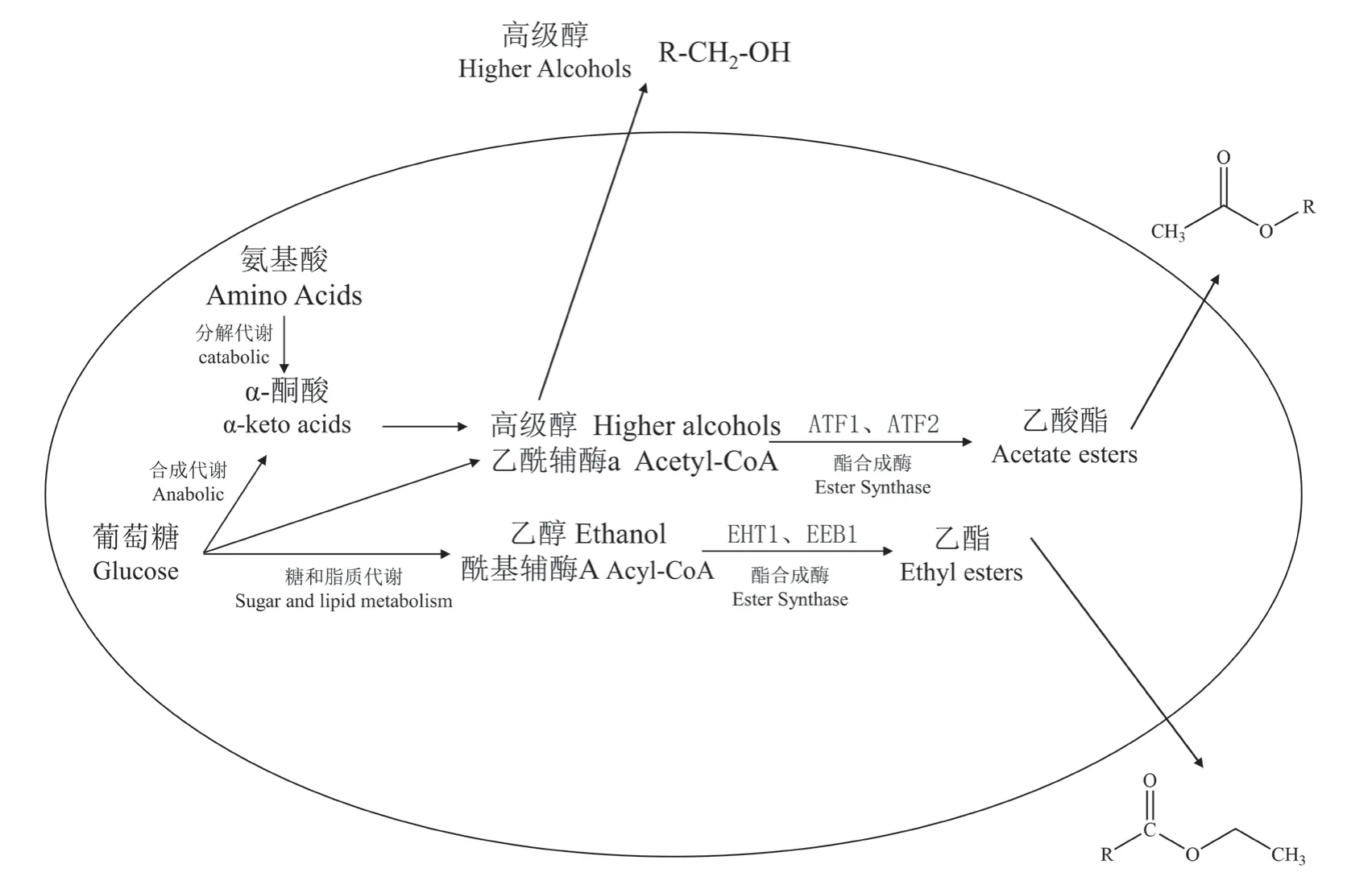
图2 酿酒酵母中高级醇和酯的形成[29]Figure 2 Formation of higher alcohols and esters in Saccharomyces cerevisiae
1.2.1 醇类化合物的合成
葡萄酒中对香气有影响的醇类物质包括脂肪族醇和芳香族醇两大类,主要有正丙醇、正丁醇、异丁醇、异戊醇、β-苯乙醇等[32]。文献报道[33]中高级醇对葡萄酒香气的贡献观点并不完全相同,有的倾向于负面影响,因为高级醇的感官属性可能依赖于特定的环境,如在高级醇总浓度低于300 mg·L-1时,能对葡萄酒芳香特征作出积极贡献,而在总浓度大于400 mg·L-1时,则会出现刺鼻和令人不快的气味。de-la-Fuente-Blanco等[33]评估了4种常见的高级醇对模拟葡萄酒香气的贡献,结果表明,异丁醇和异戊醇对葡萄酒整体香气强度有显著的影响,能够增强葡萄酒的香气感知,而β-苯乙醇和甲硫基丙醇的添加对模拟酒的香气特征基本没有影响,尽管β-苯乙醇被多次报道为葡萄酒花香特征的潜在贡献者。
高级醇的合成是由相应的中间体α-酮酸经脱羧还原生成相应的醛,再经进一步脱氢反应生成[34]。根据α-酮酸的来源,高级醇的形成分为氨基酸脱氨分解生成α-酮酸的分解代谢途径(Ehrlich代谢途径)和糖酵解、三羧酸循环生成α-酮酸的合成代谢途径(Harris途径),并且发酵液中75%的高级醇由Harris途径产生,25%是由Ehrlich途径产生[35]。
1.2.2 酯类化合物的合成
在葡萄酒中酯类化合物主要为乙酸酯和乙酯。乙酸酯类化合物主要包括乙酸乙酯、乙酸异戊酯、乙酸异丁酯、乙酸-2-苯乙酯等;乙酯类化合物主要为己酸乙酯、辛酸乙酯和癸酸乙酯等。
乙酸酯合成的底物是乙酰辅酶A(Acetyl coenzyme A,acetyl-CoA)和醇,乙酰辅酶A主要是通过丙酮酸的氧化脱羧形成,醇包括乙醇和来自氨基酸代谢产生的高级醇;乙醇或高级醇和乙酰辅酶A在醇乙酰基转移酶(Alcohol acetyltransferase,AATase)的催化作用下生成相应的乙酸酯。酿酒酵母细胞中参与乙酸酯合成的醇乙酰基转移酶有ATF1和ATF2。有研究证明ATF1对乙酸酯的生成量影响较大,可通过调控ATF1的表达来改变乙酸酯的生成量[36]。
中链脂肪酸乙酯(MCFAEE),如己酸乙酯、辛酸乙酯和癸酸乙酯,是一组广泛存在于葡萄酒、白兰地、威士忌、朗姆酒等饮料酒中的重要香气化合物,具有优质的花果香和低阈值。MCFAEE是乙醇和中链脂肪酸(MCFA)在酰基转移酶(Acyltransferase)的催化作用下生成。MCFA主要由酵母细胞通过细胞质中脂肪酸合成酶(FASs)复合体的作用释放而来,酰基转移酶主要有乙醇酰基转移酶(Ethylester biosynthesis I,EEB1P)和乙醇己酰转移酶(Ethanol hexanoyl transferase I,EHT1P),分别由EEB1和EHT1编码。
1.3 发酵作用影响陈酿香
在葡萄酒陈酿过程中,既进行着葡萄酒与环境间的物质交换,同时还发生着氧化与还原、酯化与水解和缩聚等化学反应,使酯类、醇类、醛类、酮类和酸类等香气物质达到新的平衡,通过陈酿可以促使葡萄酒色泽稳定,口感更加圆润[37]。
1.3.1 氧化、聚合反应
在葡萄酒陈酿过程中,葡萄酒中的单宁与其它多酚类物质发生微氧化和聚合反应[38]。单宁及游离花色素苷与乙醛发生聚合反应形成花色素苷-乙醛-单宁聚合物等不同芳香物质,有利于颜色稳定,从而改善了葡萄酒品质[39]。
1.3.2 橡木香气物质的溶解与扩散
与橡木相关的芳香化合物如香草醛、丁子香酚、愈创木酚等200余种香气物质选择性的在陈酿过程中转移到葡萄酒中去[40-41],增加了葡萄酒香气复杂性。
1.3.3 酯类的水解与合成反应
葡萄酒陈酿阶段,酯类物质与酸类物质之间的平衡对葡萄酒的感官品质影响最大,由发酵产生的酯在低pH环境下不断水解,结合态的萜烯类化合物被水解释放且发生分子间重排并互相转化,合成C13降异戊二烯类化合物的前体物类胡萝卜素也在不断地降解,同时,葡萄酒中的酸类和乙醇也会缓慢生成相应的酯。
陈酿过程中,酯类的构成和比例会发生改变,直链脂肪酸乙酯如己酸乙酯、辛酸乙酯、癸酸乙酯等因挥发性强,含量会逐渐降低;支链脂肪酸乙酯如琥珀酸乙酯、乳酸乙酯等因挥发性相对较差,以及陈酿期间达到新的酸酯平衡,含量会上升。对瓶储1~5年的葡萄酒中乙酯含量变化的研究发现,酸酯平衡对乙酯的形成影响较大,且随着陈酿时间的延长,酯化率明显提高,在陈酿过程中,葡萄酒中支链脂肪酸乙酯(如异丁酸乙酯、2-甲基丁酸乙酯和异戊酸乙酯)的含量增加;一部分萜烯类化合物在陈酿过程中也会发生改变,香叶醇和橙花醇可以缓慢地转化成芳樟醇和α-萜品醇,橙花叔醇能转化为法尼醇[42];C13降异戊二烯类化合物如β-大马士酮和TDN的含量会随着陈酿时间的延长而上升[43],也有研究发现在陈酿早期β-大马士酮和3-氧代-α-紫罗兰酮的含量会上升,随着陈酿时间的延长而降低[44]。由此可见,葡萄酒在陈酿过程中也发生着十分复杂的反应,由发酵作用产生的醇、酯等,以及发酵过程中新形成的萜烯和C13降异戊二烯类化合物会影响陈酿香的香气组分和比例。
2 葡萄酒发酵香气物质调控
2.1 葡萄酒品种香香气物质释放调控
2.1.1 葡萄酒酒精发酵前处理
葡萄酒酒精发酵前处理如冻融、迟采、晾晒、浸渍等,会导致发酵基质的浓度发生变化,或致使葡萄内源酶促反应剧烈,或者产生对葡萄细胞活体风味物质代谢的更大刺激,以及增强非酿酒酵母等微生物的作用,从而影响葡萄酒品种香的香气化合物组分与浓度。
(1)冻融。冻融会加速葡萄浆果细胞膜系统结构的破坏,葡萄内源酶与底物结合从而发生各种复杂反应[45],并且在冷冻过程中,脱水胁迫会诱导挥发性化合物合成相关基因(如LOX、ADH、CCD)的转录变化[46-47]。在冰酒的酿造过程中,葡萄果实经过冻融和脱水,酿造的冰酒中挥发性酚、内酯、β-大马士酮和苯乙醛的含量比干型葡萄酒更高[48]。
(2)迟采、晾晒。迟采和晾晒会导致浆果脱水,使葡萄总可溶性固形物的浓度和总酸度增大。延迟采收的果实所酿造的葡萄酒中,中链脂肪酸和乙酸酯含量变低,而γ-壬内酯和β-大马士酮的含量变得更高[49];葡萄采后经脱水处理,利用质量损失率20%和30%的果实酿造葡萄酒,部分挥发性化合物种类减少,但大马士酮、橙花醇和1-己醇等化合物的含量有所提高[50];在室内自然通风脱水7、14、21 d的‘美乐’果实(质量相应损失10%、20%和27%),其萜烯如α-松油醇、6-甲基-5-庚烯-2-酮和芳樟醇的浓度增加,这可能是由于脱水促进了葡萄中真菌的生长,使得糖苷态香气前体充分释放[51]。
(3)浸渍。葡萄果实中香气物质主要位于果皮中,发酵过程中浸渍处理利于果皮中香气物质的浸出。浸渍方式按温度不同分为3种:热浸渍、常温浸渍和冷浸渍,其中冷浸渍更有利于萜烯类物质的释放。低温处理(4、10 ℃)葡萄样品后进行发酵,使葡萄酒中倍半萜烯普遍增加,推测低温诱导了萜类生物合成的关键基因(VvHDR、VvDX3、VvTER、VvGT14)的表达[52];在一定范围内,葡萄酒中萜烯类含量的增加与浸渍时间成正比,在14 ℃下浸渍24 h后萜烯含量最高[53]。近几年衍生出一些新的浸渍工艺来替代传统方法以提高香气,常用的浸渍新技术有CO2浸渍法[54]、脉冲电场[55]、闪蒸法[56]和干缩法[57],均能显著提升葡萄酒的品质。
2.1.2 接种微生物调控
品种香香气化合物的种类、含量和组成比例受葡萄品种、生态环境、栽培模式和果实成熟度等因素的影响。目前在葡萄酒实际生产中,常利用非酿酒酵母与酿酒酵母混合发酵以及乳酸菌(酒酒球菌、植物乳杆菌等)与酿酒酵母混合发酵来调控萜烯类香气物质的含量;而果实中降异戊二烯类化合物的合成在转录水平上受前体物质类胡萝卜素含量和CCD家族基因表达量的调控,此类物质的调控研究主要集中在产CCDs相关微生物资源的挖掘方面。
(1)酿酒酵母。根据最新的分类方法,酿酒酵母属有23个种,葡萄酒酿造中常见的酿酒酵母有5个种:酿酒酵母(Saccharomyces cerevisiae)、贝酵母(Saccharomyces bayanus)、原葡萄汁酵母(Saccharomyces uvarum)、巴斯德酵母(Saccharomyces pastorianus)和奇异酿酒酵母(Saccharomyces paradoxus)。大部分酿酒酵母本身并不具有内源糖苷酶,仅有少量酿酒酵母能够表现出糖苷酶活性,从而增加葡萄酒中萜烯含量[58]。提高酿酒酵母在酒精发酵过程中对极端胁迫的抵抗力,对于有效调节和改善香气化合物的产生至关重要[59]。
(2)非酿酒酵母。非酿酒酵母(non-Saccharomyces)可以通过代谢产生糖苷酶和C-S裂解酶,能够增加葡萄酒中萜烯类、降异戊二烯类、挥发性硫醇等化合物,提高香气成分种类和含量,增强葡萄酒的品种香气和发酵香气特征,并且在酒精发酵后期非酿酒酵母可以通过细胞自溶增加氨基酸、脂肪酸、甘露糖蛋白的含量进而影响葡萄酒的口感[60-61]。目前,从葡萄汁中分离出来的非酿酒酵母已超过40种,常见的非酿酒酵母属主要包括有孢圆酵母属(Torulaspora)、有孢汉逊酵母属(Hanseniaspora)、假丝酵母属(Candida)、接合酵母属(Zygosaccharomyces)、毕赤酵母属(Pichia)、裂殖酵母属(Schizosaccharomyces)、美奇酵母属(Metschnikowia)、类酵母属(Saccharomycodes)、德巴利酵母属(Debaryomyces)、克鲁维酵母属(Kluyveromyces)等。由于一般非酿酒酵母的酒精耐受能力较差,酒精转化效率较低,生产上常与酿酒酵母混合(同时接种和顺序接种)发酵来生产干型葡萄酒[61]。
Whitener等[62]使用‘西拉’和‘长相思’葡萄顺序接种非酿酒酵母与酿酒酵母,结果发现非酿酒酵母假丝酵母菌、哈萨克斯坦酵母、耐热克鲁维酵母、美极梅奇酵母、戴尔有孢圆酵母、克鲁维毕赤酵母均能对整体香气有不同程度的贡献,美极梅奇酵母、假丝酵母等对葡萄酒发酵游离萜烯和酯类合成贡献最为突出。由此可见,不同非酿酒酵母对葡萄酒风味的影响存在明显差异。周洪江等[63]使用3种非酿酒酵母分别与酿酒酵母混合发酵,发现美极梅奇酵母(M.pulcherrima Flavia)使芳樟醇释放量信号强度增加1.33倍,戴尔有孢圆酵母(T.delbrueckii Prelude)使α-松油烯的信号强度增加1.47倍。申静云等[64]选用非酿酒酵母仙人掌有孢汉逊酵母(Hanseniaspora opuntiae)和葡萄汁有孢汉逊酵母(Hanseniaspora uvarum)分别与商业酿酒酵母DV10混合发酵,结果发现,H.opuntiae与DV10同时接种混合发酵增加了冰葡萄酒中21.40%的萜烯类物质;铁红假丝酵母(Candida pulcherrima)与本土酿酒酵母MMf9混合发酵,增加了α-松油醇、橙花醇、香叶醇的含量[65];Barbosa等[66]对葡萄牙葡萄酒产区美极梅奇酵母(Metschnikowia pulcherrima)进行分离,发现该酵母具有较高的β-葡萄糖苷酶活性。
Hu等[67]通过对菌株的筛选发现,葡萄汁有孢汉逊酵母菌株的糖苷酶提取物能够催化糖苷态C13降异戊二烯类化合物的释放,增强了花香、甜味、浆果和坚果的香气特征。尤雅等[68]采用不同接种方式在两个温度(13、23 ℃)下进行扁平云假丝酵母(Candida humilis)与酿酒酵母的发酵,结果发现,13 ℃时混合发酵显著提高了β-大马士酮含量,其中同时接种发酵增加了26.19%。类似地,与单纯的酿酒酵母发酵相比,使用仙人掌有孢汉逊酵母(Hanseniaspora opuntiae)与商业酿酒酵母DV10混合发酵使冰酒中β-大马士酮含量提高8.85%[64]。
(3)乳酸菌。乳酸菌除了分泌苹果酸脱氢酶外,还产生多种胞外酶,如β-葡萄糖苷酶、酯酶、氨基酸脱羧酶、柠檬酸裂解酶、酚酸脱羧酶等[69],从而增加葡萄酒中萜烯类、硫醇类等物质的含量。葡萄酒启动苹果酸-乳酸发酵有利于形成柔和的酸度,促进花果香气的释放,提高葡萄酒香气的复杂性。与酵母菌相似,不同属、种的乳酸菌对葡萄酒风味的贡献也有明显的差异,其中酒球菌属(Oenococcus)中的酒酒球菌(O.oeni)和乳植杆菌属(Lactiplantibacillus)的植物乳杆菌(L.plantarum)广泛应用于葡萄酒生产过程中[70]。
近些年来,植物乳杆菌凭借其高产酯酶、糖苷酶的潜力,作为酒酒球菌的替代者用于启动葡萄酒的苹果酸-乳酸发酵,在葡萄酒香气、颜色等特征的改变中起重要作用[71]。Iorizzo等[72]从葡萄酒中分离出11株植物乳杆菌,发现所有菌株在葡萄酒工业化生产条件下均表现出较强的苹果酸-乳酸发酵性能和β-葡萄糖苷酶活性,能够释放出柠檬烯、里哪醇等萜烯苷元。Wang等[73]以植物乳杆菌和酒酒球菌为原料,对‘赤霞珠’葡萄进行苹果酸-乳酸发酵,结果发现植物乳杆菌能够保留更多的香气成分,表明植物乳杆菌可以作为较好的发酵剂运用于红葡萄酒生产中。葡萄酒中的酚类物质对乳酸菌的生长有抑制作用,Devi[74]的研究结果发现,酒酒球菌对酚胁迫的耐受性优于植物乳杆菌,但植物乳杆菌具有更多功能的酚类解毒酶系统,可以使酚类化合物降解为挥发性酚、芳香醇和苯基丙酸。
(4)其它。魏涛等[75]从红茶中得到一株高效降解β-胡萝卜素菌株肠杆菌(HC-3),该菌株对β-胡萝卜素的降解率达86.82%,主要降解产物为5,6-环氧-β-紫罗兰酮、二氢猕猴桃内酯和反式-β-紫罗兰酮等香味物质。龙章德等[76]从烟叶中筛选出能高效降解β-胡萝卜素的菌株霍氏肠杆菌亚种(Enterobacter hormanii subsp.),用气相色谱质谱联用技术(GCMS)进行检测发现,其降解的主要产物为β-紫罗兰酮、二氢猕猴桃内酯、β-环柠檬醛等香味物质。杨小雪等[77]从铁观音茶叶中筛选到了3株具有β-胡萝卜素降解能力的菌株Aspergillus niger、Bacillus velezensis、Saccharopolyspora gregorii,它们的降解率分别为94.54%、72.97%、61.26%,能产生β-紫罗兰酮等香气物质。
2.2 葡萄酒发酵香生物合成调控
葡萄酒发酵香气化合物由菌株特性与葡萄汁特性决定,受发酵环境因子(温度、pH、氧浓度、营养素)调控。葡萄酒发酵过程中,酵母菌受不断变化的环境条件影响,诱导了2000多个基因的表达,涉及数百个基因编码蛋白质参与各种细胞过程[78]。
2.2.1 醇类化合物合成调控
高级醇的生成量受多重因素调控,酵母发酵副产物形成的主要因素有酵母菌株、发酵工艺和果汁类型等[79-80]。
(1)酵母菌株及接种量。不同的酿酒酵母产高级醇的能力不同,有研究发现,采用12株不同的酿酒酵母发酵相同葡萄,所产生的高级醇含量显著不同[81],其中国产酵母CC17有更高的感官评价,这种差异主要取决于酵母细胞高级醇脱氢酶(ADH)的活力。在葡萄发酵过程中,酵母的接种量对高级醇的生成有很大的影响,接种量低时,酵母的繁殖倍数会相应增大,高级醇的含量则会升高。宋含等[82]发现,酵母的增值倍数控制在3~4以内为宜,可以减少高级醇的生成量。
(2)发酵温度。发酵温度影响酿酒酵母生长速率和生物量,能够调整酿酒酵母次生代谢产物通量。研究发现,随着发酵温度的升高(10、15、20 ℃),异丁醇、异戊醇和苯乙醇的含量逐渐升高[83]。
(3)发酵醪pH。pH也能影响酿酒酵母生长速率和生物量。在葡萄酒发酵过程中,高级醇的生成量与发酵液pH具有相关性,pH越低时高级醇的含量也相应减少。当葡萄酒醪液pH控制在3.6以下时,高级醇的生成量最低[84]。
(4)葡萄醪液可同化氮(YAN)。氮素是影响微生物生长的常见因素,YAN的种类及比例影响酿酒酵母生长代谢及发酵产物含量。适宜的α-氨基氮含量会使高级醇生成量保持较低的水平,当α-氨基氮含量过低时,酵母为满足自身生长所需会不断的通过糖代谢合成氨基酸,进而形成更多的中间体α-酮酸,当α-氨基氮含量过高时,酵母增殖倍数加大,从而导致代谢副产物增多;Liu等[85]发现,不同YAN能改变高级醇合成相关基因的表达,当NH4Cl添加量为200 mg·L-1时,总高级醇含量减少了39%,且对高级醇含量的控制更有效,因此在葡萄酒生产中根据实际情况考虑添加YAN。
(5)葡萄醪液糖浓度。低浓度的糖会使高级醇前体物质α-酮酸产生减少,对高级醇的脱氨基作用也不强烈,因此葡萄酒发酵可以采取分批补糖的方式来控制高级醇的生成量[80]。
2.2.2 酯类化合物合成调控
酯的产生受底物和酶活性的影响。对于乙酸酯的形成,乙酰辅酶A和高级醇的浓度是重要限制因素。有研究表明,乙酰辅酶A的通量受温度、脂肪酸、氮源和氧的影响,这些因素在促进酵母生长的同时也能促进乙酰辅酶A的利用,使较少的乙酰辅酶A用于酯的生产;另外,可利用基因工程、蛋白质工程过表达ATF1或敲除竞争抑制途径的相关基因,提高乙酸酯的含量[86-87]。
中链脂肪酸的存在形式是影响乙酯类化合物含量的主要因素,通过对脂肪酸从头合成关键基因(ACC1、FAS1、硫酯酶编码基因)的强化可达到提高MCFAEE生成量的目的,这些基因的表达量与酵母细胞数量相关,同样受温度、脂肪酸、氮源和氧的调控[88-89]。所以通过调控葡萄发酵过程中酿酒酵母生物量、比生长率和时序寿命,可以实现调控脂肪酸合成水平进而提高乙酯含量的目的。
(1)发酵温度。发酵温度通过影响酵母生长速率和生物量从而显著影响脂质代谢,在13 ℃和25 ℃发酵条件下,转录组分析发现酿酒酵母显著差异表达的535个基因中,在25 ℃时与胞质脂肪酸合成有关的几个基因下调;而在13 ℃时与线粒体短链脂肪酸合成有关的基因上调,更有利于乙酯的积累。Schwinn等[90]研究了‘雷司令’发酵过程中3个不同温度下对酵母生长、挥发物等的影响,结果显示,大多数酯的形成在较高温度下的发酵后半段更为明显,在19 ℃发酵时,癸酸乙酯、辛酸乙酯和己酸乙酯的终浓度最大。所以,温度可以通过影响脂肪酸合成途径相关基因的表达来影响MCFAEE产量。
(2)脂肪酸。不饱和脂肪酸(UFAs)是酿酒酵母生长繁殖的必需营养物质,在发酵过程中,酿酒酵母主要从葡萄汁中获得UFAs[91]。在发酵培养基脂质组成(葡萄汁与合成葡萄汁)对酒中挥发性成分影响的研究中发现,两者完成发酵的时间分别为14、25 d,葡萄汁更快完成发酵且积累的挥发性化合物浓度更高,其辛酸乙酯的含量是合成葡萄汁发酵的两倍[92]。这与合成葡萄汁中缺乏UFAs等生长因子、厌氧条件下限制了酵母菌生长有关。同时,在发酵过程中添加外源中链脂肪酸(如己酸、辛酸)也能显著提高乙酯类化合物的含量[93]。
(3)氮源。在发酵过程中,低氮胁迫下酯类的总量降低,在500 mg·L-1以内,酯类生成量与葡萄汁可同化氮水平成正比[94]。当向初始YAN为250 mg·L-1的葡萄汁中添加磷酸氢二铵(DAP)时,脂肪酸乙酯的含量升高,乙酸酯含量却无显著差异,表明氮的利用也具有菌株特异性[95]。
(4)氧气。氧气是葡萄酒发酵过程中十分重要的因素,葡萄酒发酵并非绝对厌氧发酵,酵母菌脂肪酸代谢是在有氧条件下进行的,微量氧气可以促进酵母菌次生代谢产物的生产量。Snoek等[96]分别在需氧和厌氧条件下培养酵母菌,转录组分析表明,在500多个差异代谢基因中只有23个是厌氧菌生长所特有的,这23个基因影响着酵母菌在缺氧条件下的存活,其关键作用机制还需进一步探索。
酯类物质作为酵母的次生代谢产物,其代谢还受葡萄品种与成熟度[97-99]、发酵方式、酵母菌种、陈酿方式等[100-102]的调控,而针对这些调控,前人已作了详细的论述[103-104]。近几年来几项关于氧化还原电位(ORP)的研究证明,ORP有潜力作为一个新的观测量实现香气物质的监测与调控。Xue等[105]揭示了在葡萄酒发酵过程中ORP与MCFAEE生成量的关系;Nelson等[106]在对小规模(10 L)、中规模(1500 L)和大规模(10 000 L)葡萄酒发酵过程中ORP的控制进行了演示,将ORP控制在-40 mV,可实现对发酵过程中香气化合物的调控;在葡萄酒发酵过程中利用ORP作为曝气定时开关,最终实现了对酯类物质生成量的控制[107]。葡萄汁ORP反应了溶氧量、温度、pH、基质组分特性与浓度的关系,通过调控ORP可以影响细胞内NAD(P)/NAD(P)H平衡和MCFAEE代谢相关关键基因的表达,从而影响酿酒酵母碳通量分布。这为实现在线实时监测调控酯类化合物的合成提供了新的思路。
综上,葡萄酒发酵过程中采用适度低温、适当溶氧和控制葡萄汁脂质、适量可同化氮,提高细胞群体峰高和细胞时序寿命,降低细胞比生长率,可以达到提高酯类物质产量的目的。
2.3 葡萄酒发酵过程中香气物质转移的控制
葡萄酒发酵过程中,醪液香气化合物的浓度取决于葡萄原料和微生物代谢通量,还会随CO2的排出而发生挥发损失。发酵温度对香气挥发损失的影响比对香气合成产生的影响更为显著。在30 ℃发酵时,己酸乙酯的损失高达70%[108]。因此,有酿酒师在低温条件下(8~12 ℃)进行发酵以减少香气损失来改善葡萄酒的香气特征[29],但引发了发酵周期增长、发酵不彻底的现象。在发酵过程中的气液传递方面已有大量研究,也有多种技术用来减少芳香损失,但其中一些可能会对发酵动力学产生负面影响,如能量消耗过大而增加了生产成本,或增加停滞发酵的风险[109]。Breniaux等[110]发现,在葡萄酒发酵过程中利用膜接触器进行气液传递,可以溶解CO2并能去除O2,从而提升葡萄酒的质量;Diban等[111]基于膜分离技术提出一种蒸发渗透萃取技术,可以明显减少芳香化合物的损失;然而目前来看,降低温度是最大限度减少香气损失的有效方法,探寻更为有效地控制葡萄酒发酵过程中香气物质转移的方法将会是一个热点。
3 总结与展望
葡萄酒香气物质组成和来源非常复杂,香气质量受多种因素影响。以葡萄香气糖苷、类胡萝卜素等香气前体物水解、裂解为靶标的生物催化资源(非酿酒酵母、乳酸菌等)的挖掘与催化机制、催化技术研究;以特征发酵香香气化合物定向合成为目的的调控技术研究;以提高葡萄酒产品整体质量和开发新产品为目的的葡萄酒酒精发酵前处理原理、方法、技术研究;以降低葡萄酒发酵过程中香气损失为目的的方法、技术研究等,是当前葡萄酒香气发酵调控研究的重点。葡萄酒发酵合成生物研究或成为未来的研究热点,合成生物有望成为葡萄酒香气调控的有效工具。
猜你喜欢
中国兽医学报(2023年8期)2023-09-25
酿酒科技(2022年8期)2022-08-20
酿酒科技(2021年8期)2021-12-06
科学之谜(2021年4期)2021-07-09
军事文摘·科学少年(2021年1期)2021-02-04
林产工业(2020年2期)2020-03-30
橡塑技术与装备(2018年21期)2018-02-19
故事作文·低年级(2016年7期)2016-05-14
中国卫生标准管理(2015年14期)2016-01-15
烟草科技(2015年8期)2015-12-20
