
弹性蛋白酶调节中性粒细胞炎性募集的功能研究
2024-04-09 08:00刘洋孟玲范思佳任春光张欢
中国免疫学杂志 2024年3期
刘洋 孟玲 范思佳 任春光 张欢
(重庆医科大学基础医学院细胞与遗传教研室发育生物学研究室,重庆 400016)
中性粒细胞弹性蛋白酶(neutrophil elastase,NE)是一种丝氨酸蛋白酶,储存在中性粒细胞嗜天青颗粒中[1]。NE最初被认为其主要功能是与髓过氧化物酶和NADPH氧化酶复合物产生的活性氧协作,参与吞噬溶酶体中入侵病原体的杀伤[1]。近年来,NE参与炎症反应调节的其他作用逐渐被报道,中性粒细胞由外界激活,脱粒时释放大量NE,释放的一部分NE以活性形式结合在质膜的外表面,可溶性和膜结合的NE通过蛋白水解切割多种趋化因子、细胞因子、黏附分子和细胞表面的活性受体,进而参与细胞的趋化、黏附等生物学功能的调节[2]。除此之外,NE还作为中性粒细胞外捕获网的组成,在清除入侵病原体和造成组织损伤的过程中扮演重要角色。
中性粒细胞的迅速募集是先天免疫反应的关键,也是急性炎症的标志性事件[3]。尽管目前对于中性粒细胞迁移到炎症部位的级联反应已经有了大量的研究,建立起了包括滚动、黏附、爬行、跨内皮迁移、跨基底膜迁移以及间质迁移在内的中性粒细胞炎症募集级联学说[4]。NE一度被认为通过降解细胞外基质与基底膜促进中性粒细胞的炎性募集[5],但ROSENGREN等[6]及ISHII等[7]指出NE的敲除并不影响中性粒细胞对细胞外基质的降解,可见NE在中性粒细胞炎性募集中的功能还存在争议。
因此,理清NE在中性粒细胞炎性募集级联反应过程中的具体作用,对明确中性粒细胞迁移动力学以及探索中性粒细胞主导炎症的靶向药物筛选至关重要。本研究旨在系统性探索NE对中性粒细胞炎性募集级联反应各关键步骤的调控,及其对中性粒细胞病原体杀伤功能的调节作用。
1 材料与方法
1.1 材料
1.1.1 试剂 小鼠胰岛内皮细胞系(MS-1)购自中国科学院细胞库; N-甲酰-蛋氨酸-亮氨酸-苯丙氨酸(N-formyl-Met-Leu-Phe, f-MLF)、西维来司钠盐水合物购自Sigma公司;重组小鼠TNF-α、重组小鼠GMCSF购自Peprotech公司;D-hanks平衡盐溶液、牛纤维蛋白原(fibrinogen,FN)、D-PBS缓冲液、牛血清白蛋白(bovine albumin,BSA)、辣根过氧化物酶(horseradish peroxidase,HRP)、异硫氰酸荧光素(fluorescein isothiocyanate,FITC)、Hanks平衡盐溶液(HBSS)、羟乙基呱嗪乙硫磺酸溶液(Hepes)、二甲基亚砜(DMSO)、RPMI-1640培养基购自Solarbio公司;胎牛血清购自BI Bioscience公司;Percoll购自Cytiva公司;鲁米诺(Luminol)、异鲁米诺(Isoluminol)购自CTI公司;磷酸肌球蛋白轻链2-Ser19(pMLC)小鼠单克隆抗体购自CST公司;山羊抗小鼠IgG(H+L)耦联647红色荧光二抗、细胞远红外标记(Far Red)购自Thermo Fisher Scientific公司;微丝绿色荧光探针(Actin-Tracker Green-488)、抗荧光淬灭封片剂(含DAPI)、细胞膜红色荧光探针(Dil)、羧基荧光素二乙酸琥珀酰亚胺酯(carboxyfluorescein diacetate,CFSE)、台盼蓝购自Beyotime Biotechnology公司;4%多聚甲醛购自Sangon Biotech公司;流动池装置购自GlycoTech公司;细胞体外迁移小室(Dunn chamber 16802)购自HAWKSLEY公司;DMi8倒置荧光显微镜、SP8 激光共聚焦显微镜(Leica);Infinite Pro 200酶标仪(Tecan)。
1.1.2 实验动物 8周龄SPF级C57BL/6N小鼠,17~23 g,购自浙江维通利华实验动物技术有限公司[SCXK(浙)2019-0001]。本研究所有动物实验由重庆医科大学实验动物伦理委员会批准(IACUCCQMU-2024-0111),小鼠饲养在25 ℃、12 h光/暗循环的SPF环境中,自由饮食,所有小鼠均适应性喂养2周后进入实验,所有小鼠断颈处理后分离骨髓。
1.2 方法
1.2.1 细胞分组 小鼠骨髓来源的中性粒细胞随机分为两组,添加终浓度为10 μmol/L的西维来司钠盐(标记为SIVE组)或1‰DMSO(标记为DMSO组),两组细胞均在37 ℃、5%CO2条件下,于含有10%热灭活胎牛血清和25 ng/ml重组小鼠GM-CSF的RPMI1640培养基中培养4 h。
1.2.2 小鼠骨髓来源中性粒细胞的分离 骨髓用于分离小鼠中性粒细胞。在Hanks-1缓冲液[Hanks平衡盐溶液(HBSS)+0.5%牛血清白蛋白+10 mmol/L HEPES]中收获小鼠骨髓细胞,红细胞裂解液裂解红细胞,进行Percoll不连续密度梯度离心。从分布在Percoll的81%和60%层间收集中性粒细胞。
1.2.3 体内腹膜炎模型检测细胞过继性迁移 提前1.5 h,向小鼠腹腔注射1 ml浓度为1 mg/ml的酵母多糖悬液,诱导无菌腹膜炎;将DMSO组和SIVE组中性粒细胞分别以0.8 μmol/L CFSE和0.2 μmol/L Far Red标记,将两种细胞1∶1混合后冰上静置1 h,向每只腹膜炎小鼠单侧眼眶后静脉窦注射5×107个混合标记细胞;6 h后从对侧眼眶后静脉窦收集外周血,并以预冷的D-PBS灌洗小鼠腹腔,收集腹膜灌洗液。通过流式细胞术检测迁移到外周血及腹膜灌洗液中DMSO预处理 (CSFE标记)或SIVE预处理(Far Red标记)的中性粒细胞比例。
1.2.4 细胞体外趋化 以FITC监测催化剂f-MLF浓度梯度,使用5 μmol/L Dil标记SIVE预处理的中性粒细胞,并将其1∶1混合接种在100 μg/ml FN预包被的玻片上,等待细胞黏附30 min。组装好细胞体外迁移小室Dunn chamber,中性粒细胞会沿f-MLF浓度梯度在迁移小室中发生趋化性迁移,以30 s间隔采集30 min所有的延时图像序列,并使用Image J和Chemotaxis And Migration Tools对单个细胞趋化路径进行分析。
1.2.5 细胞黏附流动室实验 将小鼠胰岛内皮细胞 MS-1接种在100 μg/ml FN预包被的3.5 cm细胞培养皿上并生长至汇合,在37 ℃下采用浓度为50 ng/ml的TNF-α预处理4 h。D-PBS洗涤MS-1的培养皿,并将其置于流动室装置中。分别用1 μmol/L CFSE和5 μmol/L Dil标记DMSO或SIVE组细胞,按1∶1的比例混合后,以0.4 dyn/cm2的剪切流速流入。然后在荧光显微镜下观察黏附细胞并计数,每组计数10个视野。
1.2.6 细胞伸展 100 μg/ml终浓度的FN包被玻片过夜,Hanks洗涤玻片2次,封闭缓冲液(Hanks+0.25%BSA)封闭30 min,Hanks洗涤玻片2次,无尘纸蘸干水分,玻片放入6孔板中备用。细胞悬液在接种玻片上等待贴附15 min,终浓度为10 μmol/L f-MLF严格刺激2 min后用4%多聚甲醛固定后用含DAPI的封片剂封片,每组采用Image J计数不少于100颗伸展之后的细胞面积。
1.2.7 中性粒细胞极化 100 μg/ml终浓度FN包被玻片3 h,D-PBS洗涤玻片3次。细胞悬液接种在玻片上等待贴附15 min,终浓度为10 μmol/L的f-MLF严格刺激2 min后4%PFA固定。1∶200稀释兔抗小鼠p-MLC一抗,4 ℃孵育过夜,1∶200稀释山羊抗鼠耦连647红色荧光二抗避光孵育1 h,D-PBS清洗之后,绿色微丝荧光探针避光孵育45 min,D-PBS洗涤后用含DAPI抗荧光淬灭剂封片。
1.2.8 活性氧(ROS)释放 100 μg/ml FN预包被96孔板5 h,D-PBS清洗后备用;加入细胞悬液和HRP及Isoluminol、Luminol配制的反应工作液;室温放置5 min后读板检测本底值;随后加入f-MLF刺激剂,立刻读取化学发光值,计算ROS释放动力学曲线。
1.2.9 细胞吞噬作用 终浓度为0.5 mg/ml的FITC和热灭活的铜绿假单胞菌室温下孵育20 min以标记细胞,D-PBS洗涤3次去除多余染料;在含20%小鼠血清的D-PBS中37 ℃孵育30 min以调理细菌,细胞与细菌按1∶50比例,在37 ℃下共孵育30 min,用0.4%台盼蓝淬灭黏附在细胞表面的FITC荧光,流式细胞仪检测中性粒细胞吞噬荧光标记的铜绿假单胞菌比例。
1.3 统计学分析 所有数据均使用GraphPad Prism软件版本9.0绘制和分析,数据均采用3个及以上独立实验的±s,两组数据用t检验法分析,多组采用One-way ANOVA分析,组间采用Two-way ANOVA分析,P<0.05表示差异有统计学意义。
2 结果
2.1 NE调节中性粒细胞炎性募集 如图1A所示,在小鼠外周血中回收到了相近数目的DMSO或SIVE预处理的中性粒细胞,但腹膜灌洗液中回收到的SIVE预处理后的中性粒细胞数目与DMSO组相比显著降低(P<0.001,图1B),提示NE的抑制显著减少中性粒细胞在体内的炎性募集。
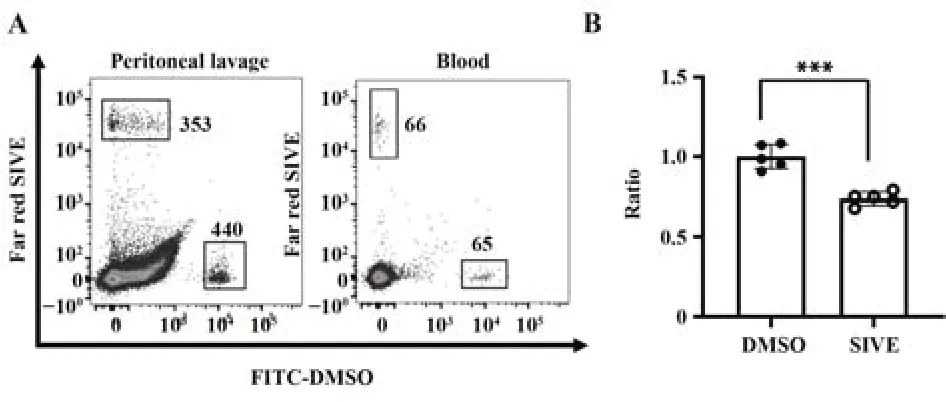
图1 NE在腹膜炎模型中对中性粒细胞过继性迁移的影响Fig.1 Effect of elastase on adoptive migration of NE in peritonitis model
2.2 NE对中性粒细胞趋化的影响 采用Dunn Chamber评估NE是否影响中性粒细胞对催化剂浓度梯度的感知能力及体外定向趋化能力(图2A、B)。Dunn Chamber时间序列图像显示,NE的抑制不仅降低了中性粒细胞顺从催化剂f-MLF浓度梯度的迁移速度(P<0.05,图2C)、干扰了中性粒细胞迁移方向性(P<0.05,图2D),同时缩短了迁移距离(P<0.05,图2E、F),表明抑制NE减弱了f-MLF诱导的中性粒细胞趋化能力。

图2 NE对中性粒细胞体外趋化的影响Fig.2 Effect of elastase on chemotaxis of NE in vitro
2.3 NE对中性粒细胞与内皮细胞黏附的影响 与DMSO预处理组相比,SIVE预处理后的中性粒细胞在存在持续性的流动剪切应力的情况下,黏附在TNF-α预处理的小鼠内皮细胞上的数目显著降低(P<0.000 1,图3A、B),即抑制NE显著减弱中性粒细胞对发炎内皮细胞的黏附能力。

图3 NE对中性粒细胞与内皮细胞黏附的影响Fig.3 Effect of NE on adhesion of neutrophils to endothelial cells
2.4 NE对中性粒细胞伸展的影响 评估两组细胞伸展能力的差异,结果发现在伸展过程中,两组细胞具有相似的形态和相似的面积,即使外源性添加了刺激剂f-MLF,两组细胞仍然具有类似的伸展形变模式(P>0.05,图4A、B),即抑制NE对中性粒细胞的伸展能力无显著影响。
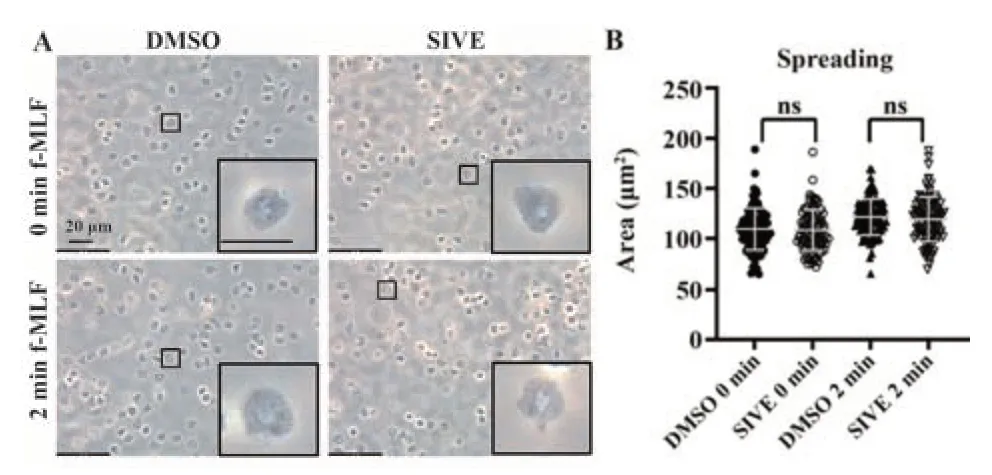
图4 NE对中性粒细胞伸展的影响Fig.4 Effect of NE on neutrophil extension
2.5 NE对中性粒细胞极化的影响 结果显示,无论是静息或f-MLF刺激后,DMSO或SIVE预处理后的中性粒细胞具有相似的极化现象与极化比例,即抑制NE对中性粒细胞的极化无显著影响(P>0.05,图5A~C)。
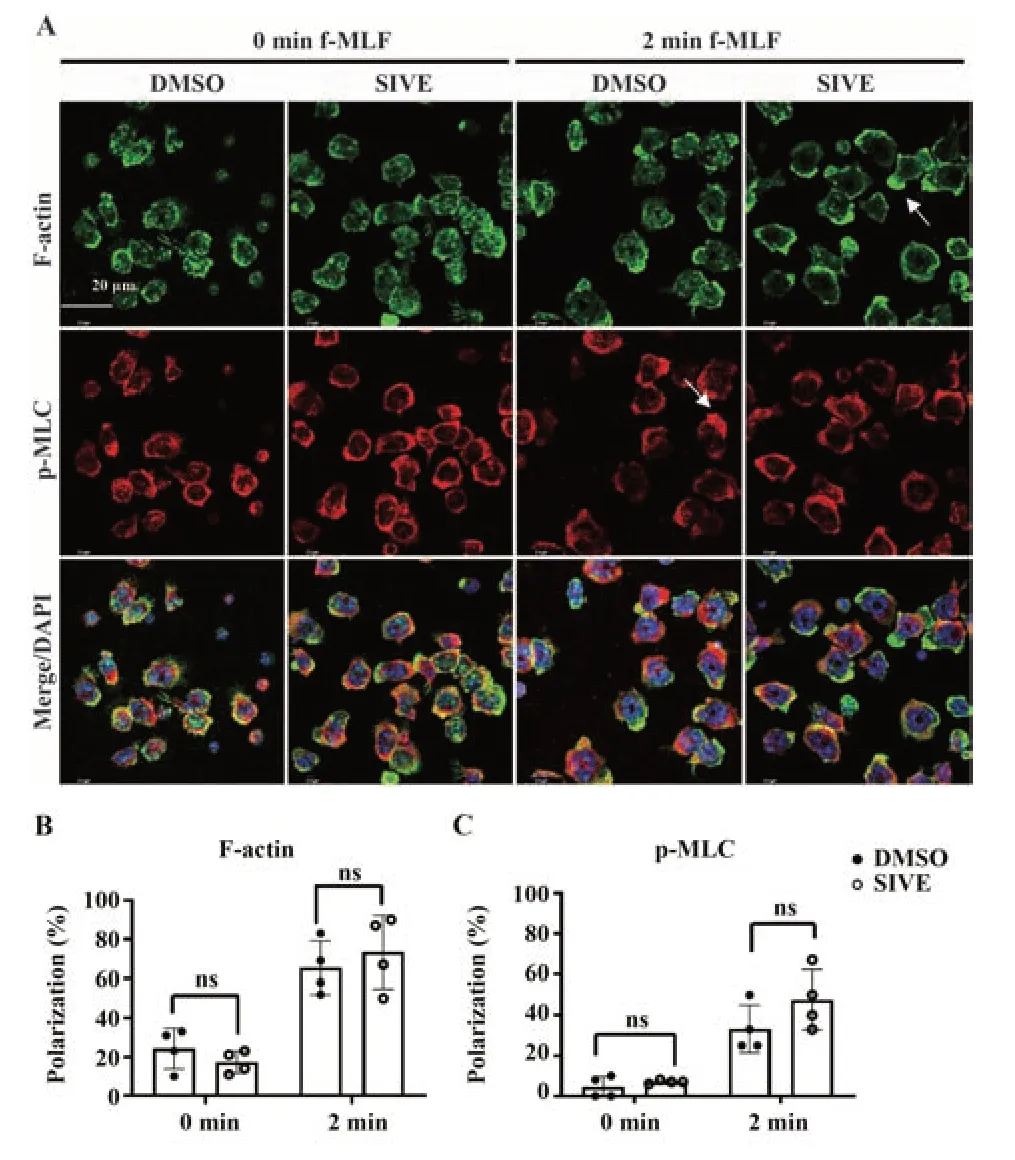
图5 NE对中性粒细胞极化的影响Fig.5 Effect of NE on the polarization of neutrophils
2.6 NE对中性粒细胞ROS释放的影响 结果显示,NE的抑制对中性粒细胞外(图6A、B)及总ROS(图6C、D)的释放无显著影响(P>0.05)。

图6 NE对中性粒细胞ROS释放的影响Fig.6 Effect of NE on ROS release in neutrophils
2.7 NE对中性粒细胞体外吞噬细菌的影响 结果显示,抑制NE可显著降低中性粒细胞吞噬细菌的能力(P<0.05,图7)。
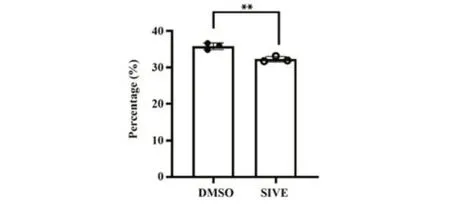
图7 NE对中性粒细胞体外吞噬细菌的影响Fig.7 Effect of elastase on phagocytosis of NE in vitro
3 讨论
本研究系统性探索了NE在中性粒细胞炎症募集级联反应各个环节的独立作用。研究结果显示,抑制NE显著减少了中性粒细胞黏附在发炎内皮上的数目,影响了中性粒细胞对催化剂浓度梯度的感知,降低了趋化速率及距离并最终导致中性粒细胞体内炎性募集及吞噬能力的损害;进一步,本研究发现F-actin及p-MLC的极化不受NE的影响,排除了F-actin聚合减少及细胞极性建立异常在此过程中的贡献,且NE的抑制对中性粒细胞的ROS释放也无显著影响。
中性粒细胞作为急性炎症反应期率先被招募到损伤部位的先锋细胞,对机体免疫反应具有重要调节作用[3]。为了快速到达损伤部位,中性粒细胞具有独特且迅速的细胞迁移机制,到达损伤部位后中性粒细胞通过吞噬、ROS释放及蛋白酶库的释放等手段消灭入侵的病原体,维持体内环境稳态[8]。因此,明确中性粒细胞的迁移动力学过程尤为重要。关于中性粒细胞迁移,最新鉴定出了多种新的分子相互作用机制,他们可能参与介导或调节中性粒细胞迁移,对先天免疫和适应性免疫至关重要[9]。尽管对细胞运动领域的探索在一直深入,但迄今为止,中性粒细胞迁移过程中涉及许多特定分子功能作用的报道仍然存在一些争议,ROS则是其中的代表分子[10]。
中性粒细胞NE由于其对内皮细胞造成损害、切割关键的内皮细胞相关黏附分子(例如,ICAM-1和VCAM-1)及其水解基底膜和血管外组织成分的能力,被视为导致血管和组织损伤的介质[2,11-12]。事实上,NE的不适当释放与许多炎症性疾病的病理相关,例如局部缺血/再灌注损伤和慢性阻塞性肺病[13-14]。
西维来司钠盐作为现阶段唯一投入临床使用的中性粒细胞NE特异性抑制剂,在临床和动物实验中均显示出良好的NE活性抑制能力[15-16]。在动物实验研究中,在许多炎症模型中西维来司钠盐表现出对中性粒细胞介导的组织损伤具有保护作用[17],体外研究也有相似结果的报道,即NE抑制剂损害白细胞的体外迁移[18],但也有研究给出了相悖的结论[6],目前药理学抑制剂对白细胞特别是中性粒细胞迁移能力影响的报道仍具有矛盾性。
考虑到既往研究通常仅关注中性粒细胞炎性募集的单个环节,且体外实验难以完全模拟体内细胞迁移的整体炎症微环境。为了排除潜在因素的影响,本研究首先通过向腹膜炎小鼠眼眶后静脉窦注射等量不同标记的野生型或NE抑制的中性粒细胞,使两种细胞处在完全一致的体内炎症微环境中,证实了NE的抑制显著降低了中性粒细胞体内炎症募集[19]。在明确了抑制NE对体内募集的影响后,本研究将炎症募集过程中的关键步骤进行拆分,逐一验证NE对其是否具有调控作用。化学趋化,即细胞定向迁移,发生在跨内皮和间质迁移过程中[3]。与CAI等[20]报道拮抗NE阻断中性粒细胞的趋化性类似,本研究同样证实了NE的抑制损害了中性粒细胞定向趋化的能力,包括偏移角度的增加、趋化速度的降低,趋化距离的缩短。这将NE对迁移作用的影响节点从DELCLAUX等[21]报道的对细胞外基质与基底膜的降解提前到了定向趋化。本研究进一步探索了定向趋化的前置步骤——牢固黏附,有效的免疫反应取决于白细胞迅速从身体的一个位置移动到另一个位置的能力;中性粒细胞在血管中面临血液流动带来的剪切应力,在遇到激活分子(如受到炎症因子刺激后的内皮细胞上表达的趋化因子)时通过整合素黏附,以驻留在内皮细胞表面,流动室实验在最大程度上模拟了这个过程,等量差异标记的DMSO或SIVE预处理后的中性粒细胞,通过流动室流经炎症因子预处理的内皮细胞,在持续性剪切应力的条件下,NE抑制的中性粒细胞黏附在内皮细胞上的数目显著变少[22]。以上结果可能归因于中性粒细胞受到炎症因子刺激后,分泌到细胞外环境中或结合在质膜外表面的NE活性被抑制,进而无法对趋化因子、细胞因子、生长因子和细胞表面的活性受体进行正确加工,最终导致中性粒细胞黏附及趋化异常[23-24]。
此外,细胞的伸展形变是中性粒细胞黏附和沿血管迁移以及间质组织迁移的初始步骤,由此本研究检测了中性粒细胞伸展形变的形态与面积[4]。但无论是否存在f-MLF刺激,DMSO或SIVE预处理的中性粒细胞都具有类似的伸展能力。肌动蛋白是中性粒细胞伸展形变的关键参与者,肌动蛋白多聚化可直接促进细胞伸展形变,与前一部分研究结果相一致,NE的抑制并没有影响F-actin的聚合,两种细胞具有相似的F-actin极化比例[25]。考虑到细胞极化是细胞发生迁移性行为的前提条件,极化的方向性也决定了细胞迁移的方向[26-27]。但REN等[26]报道化学信号的刺激并不是引起中性粒细胞骨架极化的充要条件,而是细胞贴附引起的形变即诱导了细胞最初极性的建立。由此,本研究探索了外源性化学信号刺激时DMSO或SIVE预处理对中性粒细胞极化的影响,结果显示无论是否添加f-MLF,NE抑制均影响中性蛋白酶的极化。总之,NE抑制中性粒细胞体内炎症募集可能依赖于阻止了膜结合NE对细胞表面趋化因子受体的切割这种独立于F-actin的途径发生。随后,本研究回归到了NE与中性粒细胞免疫学功能的串扰,既往研究报道中性粒细胞ROS的释放可能是细胞伸展形变的下游表型[28],与之一致的是,本研究并没有发现NE的抑制对中性粒细胞ROS的释放有显著影响。与YOUNG等[29]报道NE的敲除影响中性粒细胞吞噬功能一致,本研究同样观察到NE的抑制损害了中性粒细胞的吞噬功能,对吞噬的影响可能是由于NE抑制后对中性粒细胞感知病原体的表面分子无法进行正确的切割导致参与吞噬作用的受体(如CD14、CD206)异常,最终引发吞噬能力减弱[30]。
综上所述,本研究探索了NE在中性粒细胞体内炎性募集各个关键环节的作用,通过体内腹膜炎过继性迁移实验结合体外Dunn Chamber与流动室实验对体内迁移过程的模拟,将NE影响中性粒细胞募集可能环节从既往报道的跨基底膜迁移前移到了牢固黏附。但本研究仍然缺乏对炎性募集过程中中性粒细胞表面受体的定量分析,同时也需要遗传NE缺陷模型和更详细的机制研究来进一步巩固以上发现。本研究系统性地报道了中性粒细胞经由药理学抑制剂处理后对中性粒细胞炎症募集各个关键环节的影响,为阐明中性粒细胞迁移动力学,寻找干预中性粒细胞介导的炎症疾病靶向药物的筛选提供了分子基础。
猜你喜欢
数学年刊A辑(中文版)(2022年1期)2022-08-20
数学物理学报(2021年5期)2021-11-19
中国民间疗法(2021年14期)2021-08-30
数学年刊A辑(中文版)(2020年2期)2020-07-25
英语学习(上半月)(2019年9期)2019-10-10
米娜·女性大世界(2016年8期)2016-08-17
工业设计(2016年11期)2016-04-16
医学研究杂志(2015年3期)2015-06-10
哈尔滨师范大学自然科学学报(2015年2期)2015-04-19
中国当代医药(2015年21期)2015-03-01
