
斜纹夜蛾幼虫肠道不同部位对 Zn2+积累作用的比较研究
2010-11-22 08:00舒迎花袁才亚张古忍
环境昆虫学报 2010年3期
舒迎花,袁才亚,张古忍
(1.华南农业大学热带亚热带生态研究所,广州 510642;2.中山大学有害生物控制与资源利用国家重点实验室/昆虫学研究所,广州 510275;3.深圳市龙岗区实验学校,深圳 518172)
昆虫肠道是摄取、运送、消化食物、吸收营养物质以及控制水分平衡和排泌作用的重要通道,可分为前、中和后肠三个部位。前肠具有摄食、磨碎和暂时贮藏食物的功能;中肠是分泌各种消化酶、消化食物及吸收营养物质的主要部位;后肠除了排泄食物残渣和代谢废物之外,还有吸回水分和无机盐类、调节血液渗透压和离子平衡的功能。
由于肠道各段结构和功能的不同,它们对食物中重金属离子的积累作用必然存在差异。有报道表明中肠是很多昆虫种类金属离子吸收和调节平衡的主要通道,如弹尾目、缨尾目、鞘翅目、同翅目、毛翅目、鳞翅目、膜翅目和双翅目等(Hensbergen,2000;Niu et al.,2002)。Devkota和 Schmidt(1992)用含 Hg的饲料饲养绿纹蝗 Aiolopus thalassinus Fabricius,发现汞 Hg在中肠中的含量最高。家蝇 Musca domestica L中肠是积累砷 As的主要器官,其含量是身体的 6~107倍,身体 15% ~71%的 As都积累在中肠中(Thomas et al.,1987)。家蝇体内的重金属主要储存在腹部,变态后重金属会随中肠的重建回到中肠上皮细胞中(Tylko et al.,2005)。弹尾目昆虫长角跳虫 Orchesella cincta L经 Cd胁迫后,中肠的 Cd吸收量会显著增加(Posthuma et al.,1992)。更有研究提议由于昆虫中肠是调节金属平衡和储存金属的最重要的器官,所以其上皮细胞的改变可以作为重金属污染的检测标记(Pawert et al.,1996)。相对于昆虫中肠,重金属在陆生昆虫前肠、后肠积累特点的研究还未见报道。
生活在陆地环境中的昆虫主要通过呼吸和摄食而受到重金属污染的影响 (Heliövaara and Väisänen,1993)。在自然条件下,斜纹夜蛾 Spodoptera litura Fabricius是一种分布广泛的鳞翅目植食性害虫,其幼虫主要取食庄稼如棉花、大豆、花生、土豆和十字花科蔬菜,老熟幼虫钻进土壤中化蛹,在土壤经过化蛹期,羽化变成蛾,不再取食(Etman and Hooper,1979;Matsuura and Naito,1997;Qin et al.,2004)。土壤中的重金属主要通过植物的吸收作用进入斜纹夜蛾幼虫的食物中而对其产生影响。根据珠江三角洲农田生态系统中的 Zn2+污染程度和受污染环境中十字花科蔬菜叶和花苔中 Zn2+的含量(吴金桂等,1996),本文在斜纹夜蛾幼虫人工饲料中添加不同浓度(0~1000 mg Zn/kg)的 Zn2+,连续三代饲养幼虫,对连续三代 6龄幼虫消化道前肠、中肠和后肠中的 Zn2+含量进行了比较分析,其结果为进一步了解昆虫消化道对重金属的积累和解毒机制提供一定的依据。
1 材料与方法
1.1 斜纹夜蛾幼虫的饲养和 Zn的胁迫处理
斜纹夜蛾是中山大学昆虫学研究所养虫室人工饲料饲养(陈其津等,2000),在人工饲料中添加不同浓度的 Zn2+,并以此连续 3代饲喂斜纹夜蛾幼虫:以第 1代幼虫化蛹后,羽化成虫所产的卵作为第 2代虫源,第 3代以此类推。重金属 Zn2+来源于ZnCl2(Merck,Darmstadt,Germany),将 ZnCl2添加在人工饲料中,使 Zn2+终浓度分别为 0(对照)、50、100、150、500和 1000 mg Zn2+/kg。以不添加 ZnCl2饲料饲养的斜纹夜蛾幼虫为对照。所有的昆虫均饲养在 27℃,相对湿度 65%,光周期 12L:12D条件下。每个处理浓度化的蛹和羽化的成虫也在相同条件下饲养。
1.2 斜纹夜蛾 6龄幼虫消化道不同部分的采集
从每个处理中挑选 30头斜纹夜蛾 6龄幼虫,先用去离子水清洗虫体,虫体自然晾干后,解剖并收集前肠、中肠和后肠,去除肠道内含物后用去离子水反复冲洗肠道内外,防止因食物残渣中的 Zn2+对检测结果的干扰,并再次用去离子水清洗,然后将收集的材料分别放入规格为 30 mm×50 mm的称量瓶中。
由于第二代 1000 mg Zn2+/kg处理下的斜纹夜蛾成虫的羽化率极低,因而本实验未检测第三代此浓度处理后的斜纹夜蛾幼虫肠道各部分的重金属含量。
1.3 斜纹夜蛾 6龄幼虫消化道不同部分中 Zn浓度的检测
将同一世代内收集的前肠、中肠和后肠置于120℃烘箱干燥 4 h后立即放入干燥器中冷却。称量 0.1 g干燥并混匀的样品放入 50 mL安培瓶中,再加入 6 mL浓硝酸,置于电炉上消化。待棕色气体消失冒白烟后移开安培瓶,自然冷却后加入 1 mL高氯酸继续消化。待棕色气体消失,得到透明溶液后取下安培瓶,冷却后用定性滤纸过滤,去离子水定容至 50 mL。以浓硝酸和高氯酸为空白对照,用等离子体原子发射光谱仪测定各个样品中的 Zn2+,每个处理 3个重复。
1.4 数据分析
斜纹夜蛾幼虫不同样品的 Zn含量按下列公式计算:
Zn2+含量 =C×50/W
(Zn含量单位为 mg Zn2+/kg)。
其中 C表示用原子吸收分光光度计测得的浓度(单位:mg/L);W表示样品的干重(单位为 g)。
样品中的 Zn2+含量数据经 lg(x+1)转换后,进行方差分析,并进行多重比较(SAS,1989),以阐明对照与处理之间以及各个处理间差异的显著性,并检验肠道各部分第 1、2代和第 3代之间差异的显著性。对斜纹夜蛾幼虫前肠、中肠和后肠的 Zn2+含量与饲料中的 Zn浓度进行多元回归分析,以了解前肠、中肠和后肠的重金属含量与饲料中 Zn2+浓度之间的关系。
2 结果与分析
2.1 三个世代 6龄幼虫前、中、后肠对 Zn2+的积累作用
检测结果表明斜纹夜蛾三个世代 6龄幼虫前、中、后肠中Zn2+含量均随着人工饲料中Zn2+浓度的升高而增加,相关性显著(图 1~3)。
当饲料中 Zn浓度为 100、150、500和 1000 mg Zn2+/kg时,第一代斜纹夜蛾幼虫前肠中 Zn含量分别为 945.82、1345.68、2870.33和 2883 mg Zn2+/kg,分别是对照的 2.33、3.31、7.07和 7.10倍,且差异显著(df=12,F=67.59,p<0.01)。但取食含有50 mg Zn2+/kg Zn浓度的饲料后,前肠中的Zn含量(780.53 mg Zn2+/kg)虽高于对照,但与之差异不显著。当第二代斜纹夜蛾幼虫取食 Zn2+浓度为50 mg Zn2+/kg的饲料后,前肠中 Zn2+含量(384.47 mg Zn/kg)与对照(299.45 mg Zn2+/kg)差异不显著,显著低于取食饲料中 Zn2+浓度分别为 100、150、500和 1000 mg Zn2+/kg后前肠中 Zn2+含量(df=12,F=291.80,p<0.01),分别是后四者的 56.91%、39.33%、19.26%和 11.68%。当斜纹夜蛾取食含有 Zn2+浓度分别为 50、100和 150 mg Zn2+/kg的人工饲料后,第三代幼虫前肠中 Zn2+含量分别为327.47、429.10和 610.62 mg Zn2+/kg,与对照(276.70 mg Zn2+/kg)以及相互间差异不显著,显著低于饲料中 Zn2+浓度 500 mg Zn2+/kg时幼虫前肠中 Zn2+含量(1756.48 mg Zn2+/kg)(df=12,F=31.29,p<0.01)。

图 1 连续 3个世代 6龄幼虫前肠的 Zn2+浓度与饲料中 Zn2+含量的多项式回归分析Fig.1 Relationships between Zn2+accumulations in foregut of the 6th instar larvae and Zn2+concentrations in artificial diets for 3 generations with regression analysis
第一代各处理斜纹夜蛾 6龄幼虫中肠中 Zn2+含量均显著高于对照,而且各处理之间差异显著(df=12,F=762.02,p<0.01)。当第二代斜纹夜蛾幼虫取食含 Zn2+浓度为 50 mg Zn2+/kg人工饲料后,中肠中 Zn含量为 561.93 mg Zn2+/kg,是对照的2.12倍,但显著低于受 Zn2+浓度为 100、150、500、1000 mg Zn2+/kg胁迫后中肠中 Zn2+含量(df=12,F=172.33,p<0.01),分别为后四者的 41.56%、22.90%、14.97%和 11.45%。当饲料中 Zn2+浓度为 50和 100 mg Zn2+/kg,第三代幼虫中肠中 Zn2+含量分别为 817.68和 829.2 mg Zn2+/kg,是对照的2.39和 2.43倍;两者显著低于取食含 Zn2+浓度分别为 150和 500 mg Zn2+/kg的人工饲料后中肠中Zn2+含量 (df=12,F=1077.42,p<0.01)。

图 2 连续 3个世代 6龄幼虫中肠的 Zn2+浓度与饲料中 Zn2+含量的多项式回归分析Fig.2 Relationships between Zn2+accumulations in midgut of the 6th instar larvae and Zn2+concentrations in artificial diets for 3 generations with regression analysis
第一代斜纹夜蛾 6龄幼虫后肠中 Zn2+含量各处理均显著高于对照,且各处理之间差异显著(df=12,F=414.51,p<0.01)。当饲料中 Zn2+浓度为50 mg Zn2+/kg时,第二代幼虫后肠中 Zn含量为 1107.98 mg Zn2+/kg,与对照(883.37 mg Zn2+/kg)差异不显著,但显著低于饲料中 Zn2+浓度为 100、150、500和 1000 mg Zn2+/kg的后肠中 Zn2+含量(df=12,F=157.03,p<0.01)。当饲料中 Zn2+浓度分别为 50、100、150和500 mg Zn2+/kg时,第三代斜纹夜蛾幼虫后肠中 Zn2+含量分别为 1310.63、1589.45、1693.80和 4159.00 mg Zn2+/kg,显著高于对照(662.6 mg Zn2+/kg)(df=12,F=274.97,p<0.01)。

图 3 连续 3个世代 6龄幼虫后肠的 Zn2+浓度与饲料中 Zn2+含量的多项式回归分析Fig.3 Relationships between Zn2+accumulations in hindgut of the 6th instar larvae and Zn2+concentrations in artificial diets for 3 generations with regression analysis
2.2 三个世代 6龄幼虫前、中、后肠中对 Zn2+积累作用比较
斜纹夜蛾幼虫连续三代取食含不同浓度的Zn2+饲料后,前、中、后肠中 Zn2+含量均随着世代的增加而显著递减,且受各个 Zn2+浓度处理后肠道各部分中 Zn2+含量世代间差异显著(图 4~6)。统计结果如下:前肠中 Zn2+含量除对照以外,各个 Zn2+处理浓度下的 Zn2+含量世代间差异显著(50 mg Zn2+/kg:F=52.33,p<0.01;100 mg Zn2+/kg:F=163.38,p<0.01;150 mg Zn2+/kg:F=308.48,p<0.01;500 mg Zn2+/kg:F=22.92,p<0.01;1000 mg Zn2+/kg:F=27.92,p<0.01)(图 4)。

图 4 Zn2+胁迫下斜纹夜蛾 3个世代幼虫前肠中 Zn2+含量比较Fig.4 Comparison of Zn2+content in the foregut of three generations S.litura larvae feeding the artificial diet treated with different concentrations of Zn2+.
当饲料中的 Zn2+浓度分别为 50、100、150和500 mg Zn2+/kg时,中肠中 Zn2+含量世代间差异显著(50 mg Zn2+/kg:F=24.84,p<0.01;100 mg Zn2+/kg:F=70.16,p<0.01;150 mg Zn2+/kg:F=133.27,p<0.01;500 mg Zn2+/kg:F=46.13,p<0.01)。当饲料中的 Zn2+浓度分别为 0和1000 mg Zn2+/kg时,中肠中 Zn2+含量世代间差异不显著(0 mg Zn2+/kg:F=1.36,p>0.05;1000 mg Zn2+/kg:F=0.96,p>0.05)(图 5)。

图 5 Zn2+胁迫下斜纹夜蛾 3个世代幼虫中肠中 Zn2+含量比较Fig.5 Comparison of Zn2+content in the midgut of three generations S.litura larvae feeding the artificial diet treated with different concentrations of Zn2+.
当饲料中的 Zn2+浓度分别为 50、100、150和500 mg Zn2+/kg时,后肠中 Zn2+含量世代间差异显著(50 mg Zn2+/kg:F=24.84,p<0.01;100 mg Zn2+/kg:F=70.16,p<0.01;150 mg Zn2+/kg:F=133.27,p<0.01;500 mg Zn2+/kg:F=46.13,p<0.01)。当饲料中的 Zn2+浓度分别为 0和 1000 mg Zn2+/kg时,后肠中Zn2+含量世代间差异不显著(0 mg Zn2+/kg:F=1.36,p>0.05;1000 mg Zn2+/kg:F=0.96,p>0.05)(图 6)。

图 6 Zn2+胁迫下斜纹夜蛾 3个世代幼虫后肠中 Zn2+含量比较Fig.6 Comparison of Zn2+content in the hindgut of three generations S.litura larvae feeding the artificial diet treated with different concentrations of Zn2+.
2.3 同一世代 6龄幼虫消化道不同部位对的 Zn2+积累作用比较
三代斜纹夜蛾幼虫取食含不同 Zn2+浓度的人工饲料后,6龄幼虫前肠、中肠和后肠中的 Zn2+含量均随着饲料中 Zn2+浓度的升高而增加(图 7~9)。
第一、二、三代斜纹夜蛾幼虫取食含不同浓度Zn2+的饲料后,一部分 Zn2+通过消化道的吸收进入体内,消化道各段吸收 Zn2+的能力具有差异,Zn2+在消化道各段中分布情况为:前肠 <中肠 <后肠,三个部位对 Zn2+的积累作用差异显著。第一代各Zn2+处理浓度下前中后肠中 Zn2+含量之间的差异(0 mg Zn2+/kg:F=3.34,p>0.05;50 mg Zn2+/kg:F=43.68,p<0.01;100 mg Zn2+/kg:F=156.25,p<0.01;150 mg Zn2+/kg:F=149.06,p<0.01;500 mg Zn2+/kg:F=94.40,p<0.01;1000 mg Zn2+/kg:F=611.04,p<0.01);第二代各Zn2+处理浓度下前、中、后肠中 Zn2+含量之间的差异 (0 mg Zn2+/kg:F=9.23,p>0.01;50 mg Zn2+/kg:F=140.22,p<0.01;100 mg Zn2+/kg:F=301.50,p<0.01;150 mg Zn2+/kg:F=976.95,p<0.01;500 mg Zn2+/kg:F=85.45,p<0.01;1000 mg Zn2+/kg:F=63.57,p<0.01);第三代各 Zn2+处理浓度下前、中、后肠中 Zn2+含量之间的差异(0 mg Zn2+/kg:F=19.76,p<0.01;50 mg Zn2+/kg:F=427.22,p<0.01;100 mg Zn2+/kg:F=105.14,p<0.01;150 mg Zn2+/kg:F=691.81,p<0.01;500 mg Zn2+/kg:F=56.30,p<0.01)。

图 7 Zn2+胁迫下第一代斜纹夜蛾幼虫消化道中Zn2+含量比较Fig.7 Comparision of Zn2+content in enteron of the 1st generation S.litura larvae feeding on the artificial diet treated with different concentrations of Zn2+.

图 8 Zn2+胁迫下第二代斜纹夜蛾幼虫消化道中Zn2+含量比较Fig.8 Comparison of Zn2+content in enteron of the 2nd generation S.litura larvae feeding on the artificial diet treated with different concentrations of Zn2+.
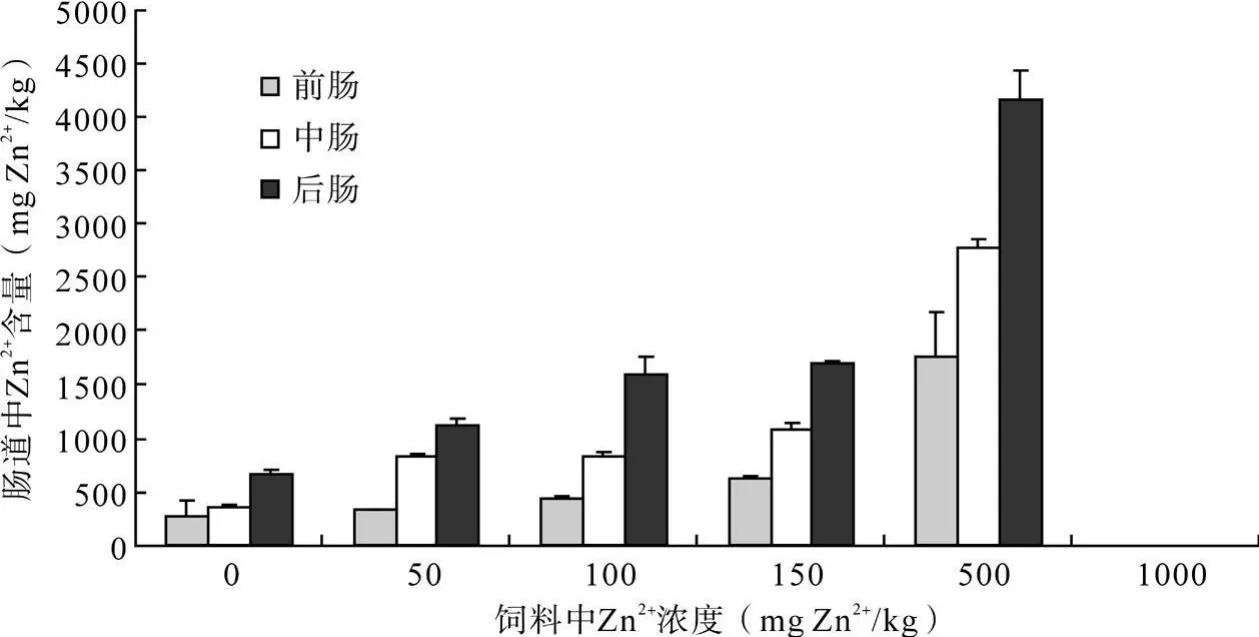
图 9 Zn2+胁迫下第三代斜纹夜蛾幼虫消化道中Zn2+含量比较Fig.9 Comparison of Zn2+content in enteron of the 3rd generation S.litura larvae feeding on the artificial diet treated with different concentrations of Zn2+.
3 结论与讨论
斜纹夜蛾连续 3个世代幼虫取食含有不同浓度 Zn2+的饲料后,过量的 Zn2+能在斜纹夜蛾每个世代 6龄幼虫肠道中积累,积累量与食物中的 Zn2+浓度显著相关;前肠、中肠和后肠三个部位对 Zn2+的积累作用依次显著增加。这与土壤、水生和一些植食性昆虫相应部位对环境中重金属的积累作用相似(孙虹霞等,2007a,2007b;夏嫱等,2005,2008)。
消化道不同部位对 Zn2+的积累作用不同,表现为前肠 <中肠 <后肠,且不同部位中 Zn2+的含量均随着 Zn2+胁迫世代数的增加而降低。当昆虫取食食物后,前肠将食物磨碎并在酶的作用下将食物降解,前肠的主要功能是暂时贮存食物和初步消化食物,对重金属离子的积累能力较弱。对重金属离子的积累作用较弱是正常的。中肠是昆虫负责营养吸收的场所,消化液中过量的重金属离子在吸收营养的过程中进入中肠细胞,其积累量应该大于前肠部位,具有很强的积累能力(Posthuma et al.,1993;Gerhardt et al.,2005);相似报道见诸于蚂蚁,体内的 Zn2+主要储存在中肠中(包括马氏管),浓度可达到 5000 ppm(Schofied et al.,1990)。后肠是排泄消化残渣的管道,食物中的营养和水分被中肠吸收后 Zn2+浓度增加,高浓度的 Zn2+通过表面渗透、吸收等方式大量进入到后肠的细胞中,这应该是后肠中 Zn2+浓度最高的主要原因。这与 Esenin等(2000)所报道的研究结果相反,皮花天牛 Rhagium inquisitor L中肠中 Zn2+浓度显著高于脂肪体,中肠对 Zn2+的积累作用显著高于后肠。
斜纹夜蛾幼虫中肠、表皮、脂肪体和血淋巴中的 Zn含量随着胁迫世代的增加而积累(夏嫱等,2005;2008)。斜纹夜蛾幼虫连续 3个世代受镍 Ni胁迫后,体内不同组织,尤其是中肠中的重金属 Ni含量随着世代增加而上升(孙虹霞等,2007a,2007b,2008)。在长期胁迫下,黄粉甲 Tenebrio molitor L体内的 Cd2+虽在变态后有所下降,但其体内的 Cd2+含量与食物中的量存在显著相关性并随胁迫时间的延长而增加(Linqvist and Block,1995)。而本文发现斜纹夜蛾幼虫前肠、中肠和后肠中的Zn2+含量随着胁迫世代的增加逐渐降低。这可能是因为不同组织对 Zn胁迫产生的不同适应反应,斜纹夜蛾幼虫通过自身调节以减轻重金属 Zn2+的毒害作用。
斜纹夜蛾幼虫连续 3个世代取食含不同浓度Zn2+的饲料后,3个世代 6龄幼虫的前、中和后肠都对 Zn2+具有积累作用,但不同部位和不同世代的积累作用有所差异。过量的 Zn对消化道不同部位细胞超微结构的影响,尤其对积累量最多的后肠细胞的影响有待进一步的研究。同时,斜纹夜蛾幼虫前肠、中肠和后肠中的 Zn2+含量随着胁迫世代的增加逐渐降低,这是否与昆虫抗性有关也有待于进一步探讨。
References)
Chen QJ,Li GH,Pang Y,2000.A simple artificial diet for mass rearing of some noctuid species.Entomol.Knowledge,37(6):325-327.[陈其津,李广宏,庞义,2000.饲养五种夜蛾科昆虫的一种简易人工饲料.昆虫知识,37(6):325-327]
Devkota B,Schmidt GH,2000.Accumulation of heavy metal sin food plant sand grasshoppers from the Taigetos Mountains,Greece.Agr.Ecosyst.Environ.,78:85-91.
Esenin AV,Ma WC,2000.Heavy metals(Cd,Cu,Zn)in wood and wood-feeding insects and other invertebrates associated with decaying pine trees.Bull.Environ.Contam.Toxicol.,4:242-249.
Etman AAM,Hooper GHS,1979.Developmental and reproductive biology of Spodoptera litura(F.)(Lepidoptera:Noctuidae).J.Aust.Entomol.Soc.,18:363-372.
Gerhardt A,Janssens de Bisthoven L,Soares AMVM,2005.Effects of acid mine drainage and acidity on the activity of Choroterpes picteti(Ephemeroptera:Leptophlebiidae).Arch.Environ.Contam.Toxicol.,48:450-458.
Heliövaara K,Väisänen R,1993.Pollution in terrestrial ecosystems.Insects and Pollution.CRCPress,Boca Raton,55-160.
Hensbergen PJ,van Velzen MJ,Nugroho RA,Donker MH,van Straalen NM,2000.Metallothionein-bound cadmium in the gut of the insect Orchesella cincta(Collembola)in relation to dietary cadmium exposure.Comp.Biochem.Physiol.C.Pharmacol.Toxicol.,125:17-24.
Lindqvist L,Block M,1995.Excretion of cadmium during moulting and metamorphosis in Tenebrio molitor(Coleoptera:Tenebrionidae).Comp.Biochem.Physiol.C.Pharmacol.Toxicol.,111:325-328.
Matsumoto T,Nakamura AN,Mori K,Kayano T,2003.Molecular characterization of a c DNA encoding putative vitellogenin from the pacific oyster Crassostrea gigas.Zool.Sci.,20:37-42.
Niu CY,Jiang Y,Lei CL,Hu C,2002.Effects of cadmium on housefly:infuence on growth and development and metabolism during metablorphosis of housefly.Entomol.Sin.,27-33.
Pawert M,Triebskorn R,Gräff S,1996.Cellular alterations in collembolan midgut cellsas a marker of heavy metal exposure:ultrastructure and intracellular metal distribution.Sci.Total Environ.,181:187-200.
Posthuma L,Van Straalen NM,1993.Heavy-metal adaptation in ter-restrial invertebrates:a review of occurence,genetics,physiology and ecological consequences.Comp.Biochem.Physiol.C.Pharmacol.Toxicol.,106:11-38.
Qin H,Ye Z,Huang S,Ding J,Luo R,2004.The correlations of the different host plants with preference level,life duration and survival rate of Spodoptera litura Fabricius.Chin.J.Appl.Ecol.,12:40-42.
SAS Institute Inc,1989.SAS/STAT User's Guider.Ver 6,4thed.Vol 1,2.Cary,NC:SASInstitute Inc.
Schofield R,1990.X-ray microanalytic concentration measurements in unsectioned specimens:A technique and its application to Zn,Mn and Fe enriched mechanical structures of organisms from three phyla.PhD dissertation,University of Oregon,Eugene.
Sun HX,Shu YH,Tang WC,Wang Q,Zhou Q,Zhang GR,2007a.Nickel accumulation and itseffectson the survival rate of Spodoptera litura Fabricius under continuous nickel stress.Chin.Sci.Bull.,52(12):1413-1418.[孙虹霞,舒迎花,唐文成,王祺,周强,张古忍,2007a.重金属Ni连续胁迫导致其在斜纹夜蛾体内积累并降低存活率.科学通报,52(12):1413-1418.]
Sun HX,Xia Q,Zhou Q,Zhang GR,2008.Accumulation and excretion of nickel in Spodoptera litura Fabricius larvae fed on diets with Ni2+.Acta Entomol.Sin.,51(6):569-574.[孙虹霞,夏嫱,周强,张古忍,2008.斜纹夜蛾幼虫对食物中重金属 Ni的积累与排泄.昆虫学报,51(6):569-574.]
Sun HX,Zhou Q,Tang WC,Shu YH,Zou ZW,Zhang GR,2007b.Metallothionein expression induced by nickel accumulation in the midgut of Spodoptera litura Fabricius larvae exposed to nickel.Chin.Sci.Bull.,52(22):2613-2617.[孙虹霞,周强,唐文成,舒迎花,邹志文,张古忍,2007b.Ni在斜纹夜蛾幼虫中肠的积累并诱导金属硫蛋白表达.科学通报,52(22):2613-2617.]
Tylko G,Banach Z,Borowska J,Nikliska M,Pyza E,2005.Elemental changes in the brain,muscle,and gut cells of the housefly,Musca domestica,exposed to heavy metals.Microsc.Res.Tech.,66:239-247.
Wu JG,Li ZM,Xu YD,Lou DR,1996.Research on the luxurious accumulation of zinc in vegetables.Chin.J.Soil Sci.,27(5):228-229.[吴金桂,李正明,徐跃定,娄德仁,1996.蔬菜奢侈吸锌特性研究.土壤通报,27(5):228-229.]
Xia Q,Shu YH,Hu XJ,Sun HX,Zhang GR,2008.Zinc accumulation in different tissues of Spodoptera litura Fabricius larvae.Acta Scientiarum Naturalium Uni.Sunyatseni:Nat.Sc.,47(3):89-93.[夏嫱,舒迎花,胡新军,孙虹霞,张古忍,2008.斜纹夜蛾幼虫不同组织对Zn积累作用的比较研究.中山大学学报(自然科学版),47(3):89-93.]
Xia Q,Sun HX,Hu XJ,Shu YH,Gu DX,Zhang GR,2005.Apoptosis of Spodoptera litura larval hemocytes induced by heavy metal zinc.Chin.Sci.Bull.,50(23):2613-2616.[夏嫱,孙虹霞,胡新军,舒迎花,古德详,张古忍,2005.重金属 Zn胁迫诱导斜纹夜蛾幼虫血细胞凋亡.科学通报,50(23):2613-2616]
猜你喜欢
大连海洋大学学报(2022年5期)2022-11-17
中国生物防治学报(2022年3期)2022-07-09
少儿科学周刊·少年版(2022年10期)2022-06-27
保健文汇(2022年1期)2022-06-06
放射学实践(2020年2期)2020-03-13
贵州农业科学(2019年7期)2019-08-03
江苏农业科学(2016年11期)2017-03-21
中国继续医学教育(2015年1期)2016-01-06
环境昆虫学报(2014年6期)2014-12-03
华东师范大学学报(自然科学版)(2014年1期)2014-04-16

- 环境昆虫学报的其它文章
- 欢迎订阅 2011年《环境昆虫学报》杂志