
基于植物能量功能群的草原植物光合作用研究
2011-03-31 06:12李维立鲍雅静孙丽睢晓光覃名茗
草业科学 2011年4期
李维立,鲍雅静,孙丽,2,睢晓光,覃名茗
(1.大连民族学院环境与资源学院,辽宁 大连 116600;2.内蒙古大学生态与环境科学系,内蒙古 呼和浩特 010021)
植物功能群(plant functional groups,PFGs)是具有确定的植物功能特征的一系列植物的组合,是生态学家为研究植被对气候变化和干扰的响应而引入的生态学概念[1]。植物能量功能群(energy plant functional groups,Energy PFGs)是在对植物热值进行分析测定的基础上,依据植物的能量属性——单位质量干物质在完全燃烧后所释放出来的热量值,采用人为分段的方法进行的植物功能类群的划分(高能值植物功能群、中能值植物功能群和低能值植物功能群)[2]。光合作用是指绿色植物通过叶绿体,利用光能,把二氧化碳和水转化成储存着能量的有机物,并且释放出氧的过程。植物通过光合作用为自身提供可供消耗的能量物质,也为食物链源源不断输入可供下一级利用的物质与能量[3]。随着一天中二氧化碳浓度和光照强度的变化,植物光合作用也发生着变化。热值反映了绿色植物在光合作用中固定太阳能的能力,直接反应植物对太阳能的转化效率,依据植物热值高低划分的功能类群,是否会表现出光合作用特性的差异,其作用机制又是怎样的?为了探究二者的关系,本研究拟对内蒙古草原不同能量功能群的植物光合日进程进行分析,在此基础上探讨不同能量功能群植物的能量固定与利用策略。
1 材料与方法
1.1 研究区概况本研究所选样地位于内蒙古锡林郭勒盟白音锡勒牧场境内的典型羊草草原,毗邻中国科学院内蒙古草原生态系统定位研究站。研究区属于内蒙古锡林郭勒盟东部、大兴安岭南端西侧的锡林河流域(43°26′~ 48°08′N,116°04′~ 117°05′E),属温带典型草原,该类型草地分布面积约20万km2,具有极为广泛的地理代表性。主要区系成分为达乌里-蒙古种,旱生草本植物为主。
研究区处于北半球中纬度接近内陆地区,气候属于温带半干旱草原气候[4]。冬季寒冷干燥,夏季受季风影响,较为温和湿润。3-5月常有大风,月平均风速达4.9 m/s,年均温为0.6℃,1月平均气温为 -21.3℃,7月为18.6℃。无霜期91 d。草原植物生长期约150 d。年降水约为350 mm,5-9月降水量占年降水量的86%。降水量的季节和年度变化非常大,少雨年份283.2 mm(1982年),多雨年份507 mm(1998年)。年蒸发量1 600~1 800 mm,相当于降水量的4~5倍。
实验样地地带性土壤为栗钙土,包括典型栗钙土和暗栗钙土两大亚类。栗钙土的腐殖质层呈较均匀的栗色,厚 25~35 cm,腐殖质含量 1.5%~3.0%;地面50 cm以下有明显的钙积层,紧密而坚实,本区栗钙土多砂壤质地,砂性较重,经开垦或过度放牧后,易遭受风蚀,往往就地起沙,形成沙地。
本研究工作主要是在定位站附近的围封样地(43°38′N,116°43′E)进行 ,该样地原为打草场 ,于2008年围封,可看作是受干扰较轻的羊草草原群落。羊草群落草层生殖枝高度可达50~60 cm,叶层高度30 cm左右,有明显的草层分化。草丛盖度一般为30%~40%,多雨年份可达60%~70%,地面有不连续的薄层凋落物覆盖,主要为高能值植物羊草(Leymuschinensis)、大针茅(Stipa grandis)建群的群落,中能值植物冰草(Agropyron cristatum)、糙隐子草(Cleistogenes squarrosa)、黄囊苔草(Carex korshinskyi)以及低能值植物猪毛菜(Salsola collina)等作为亚优势种、伴生种存在,植物种类16种,地上生物量(178.5±50.1)g/m2,地下生物量1 024.194 g/m2。
1.2 取样与测定方法在所选实验样地内,于7月底至8月中旬,用LI-6400光合仪测量相同条件下的8种草原植物光合作用特征,取其净光合速率(以CO2计)、气孔导度(以CO2计)及蒸腾速率(以CO2计)值。测量从 6:00开始,每2 h测1次,选取了高能值植物2种:羊草、羽茅(Achnatherum sibiricum);中能值植物3种:糙隐子草、冰草、艹洽草(Koeleria cristata);低能值植物3种:刺穗藜(Chenopodium aristatum)、灰绿藜(C.glaucum)、星毛委陵菜(Potentilla acaulis)。植物能量功能群分类依据鲍雅静等[2]提出的分类方法。实验过程中,每种植物分别选取3株,糙隐子草选择一个分枝上的第3或第4片叶面积较大的叶片,羊草、星毛委陵菜、羽茅、冰草 、草、刺穗藜、灰绿藜选择每株植物上第2或第3片叶面积较大的、叶质较好的叶片。测量过程中,用记号笔标记了每片叶片测量的部位,使日进程整个连续的数据测量过程在同一片叶片的同一部位进行,这样可使一株植物的同一叶片被测量8次,每种植物各有3株植物被测量,数据处理时取平均值。测量需在30 min内完成,这样6:00、8:00、10:00、12:00、14:00、16:00、18:00、20:00 各测 1 次,共 8次,以反映植物一整天的光合日进程。
2 结果与分析
2.1 各功能群净光合速率大小比较高能值植物功能群净光合速率曲线大都位于较低位置,说明高能值植物的净光合速率较小(图1)。中能值植物功能群净光合速率曲线大都位于低能值之下,高能值之上,说明中能值植物的净光合速率居中。而低能值植物功能群净光合速率曲线位于图1中最高位置,说明低能值植物反而具有较高的净光合速率。即不同能量功能群植物的净光合速率表现为高能值植物<中能值植物<低能值植物的趋势。
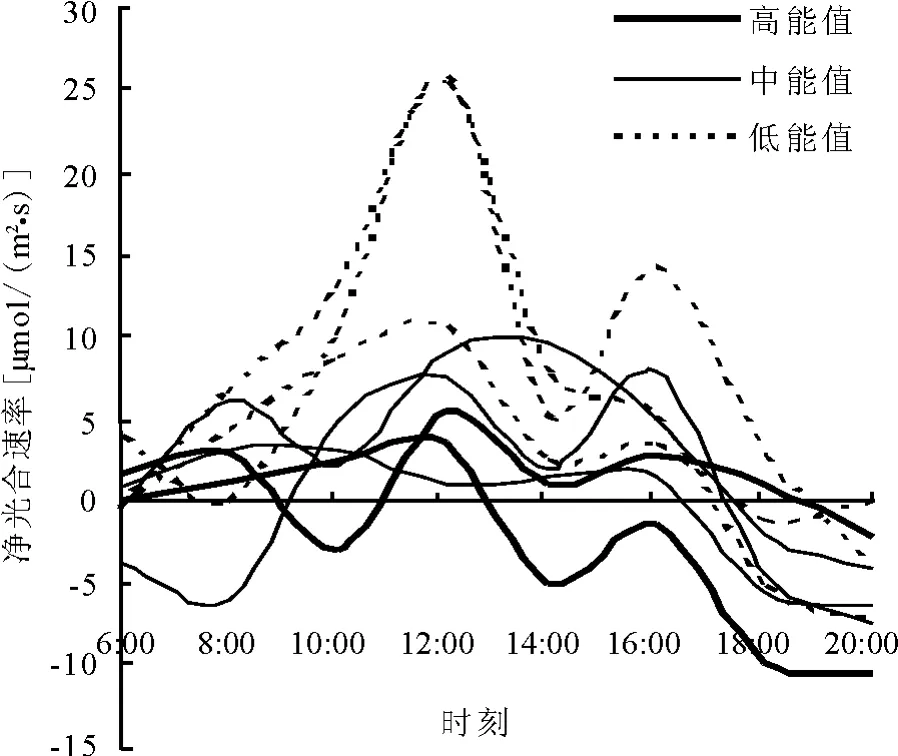
图1 被测植物净光合速率日变化
2.2 各功能群代表植物净光合速率大小比较分别选取不同能量功能群的代表性植物,代表高、中、低能值植物功能群的植物分别是羊草、糙隐子草和星毛萎陵菜,选取原因是这3种植物在草原分布较广,数量较多。在一天中的大部分时间,3种植物的净光合速率表现为:星毛萎陵菜>糙隐子草>羊草(图2),这正符合图1功能群分类时的大小情况,因而说明这3种植物是可以代表3种植物能量功能群的。3种植物的净光合速率日进程曲线都是呈“双峰”的,即都存在2个净光合速率峰值,分别在12:00和16:00,存在1个净光合速率的低点,都是14:00,原因是这个时候内蒙古草原区太阳照射过于强烈,植物自身保护机制启动,降低了净光合速率的值。
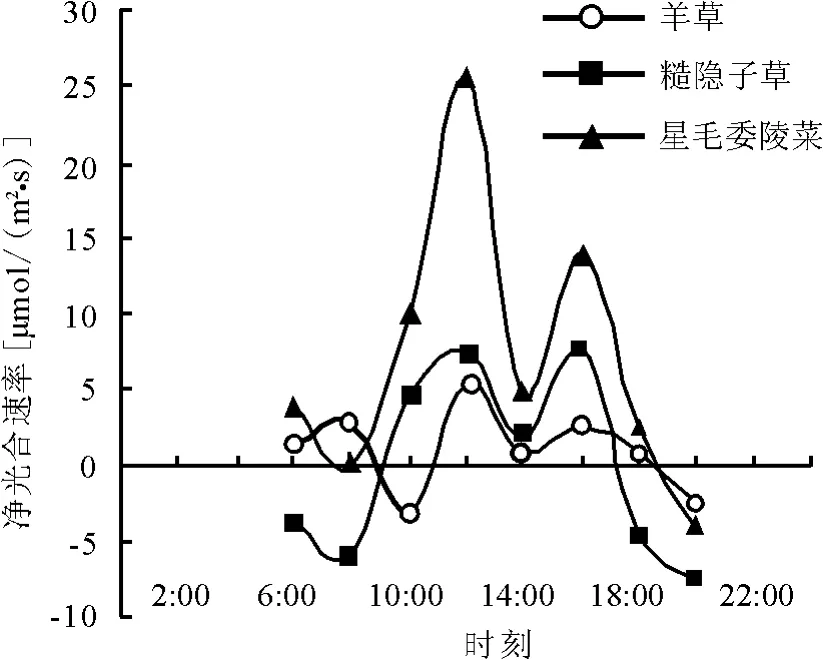
图2 不同能量功能群代表植物净光合速率日变化
2.3 各功能群代表植物气孔导度大小比较气孔导度曲线(图3),依然是星毛萎陵菜居上,糙隐子草居中,羊草居下,即气孔导度的大小,依然是低能值植物最高,中能值植物次之,高能值植物最低。气孔导度表示气孔张开的程度,植物在光下进行光合作用,经由气孔吸收CO2,且它直接与蒸腾作用成正比,与气孔阻力呈反比。说明低能值植物净光合速率大都以大的气孔导度为前提的,即吸收较多的CO2原料,自身光合作用很强。
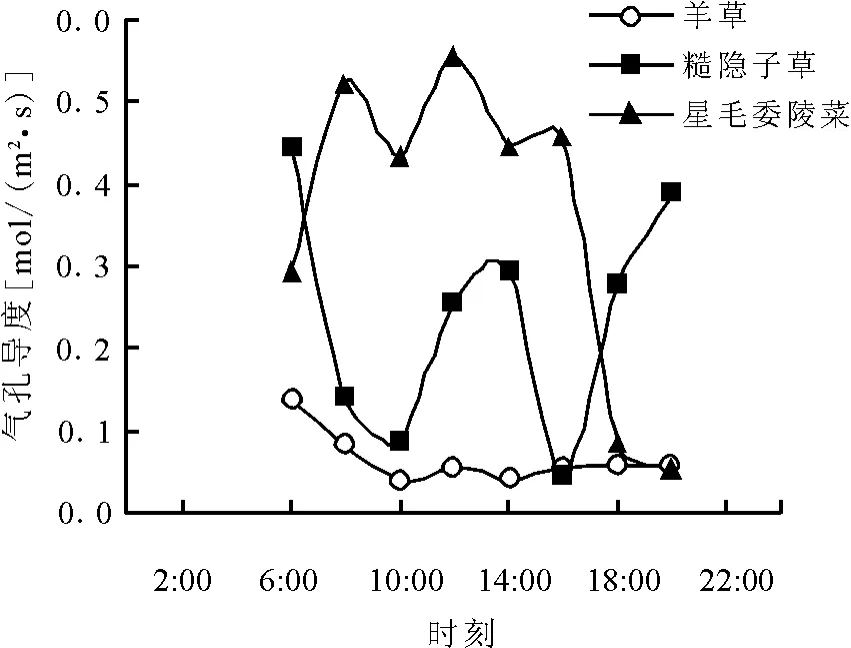
图3 不同能量功能群代表植物气孔导度日变化
2.4 各功能群代表植物蒸腾作用大小比较蒸腾速率曲线(图4),星毛萎陵菜居上,糙隐子草居中,羊草居下,即低能值植物蒸腾速率最高,其次是中、高能值植物。这与气孔导度与蒸腾速率成正比相符[5],由此可知,低能值植物生存需水量较大。

图4 不同能量功能群代表植物蒸腾速率日变化
综合分析不同功能群代表植物的净光合速率、气孔导度和蒸腾速率发现:在14:00时植物“午休”,所以净光合速率很小,但是气孔导度和蒸腾速率仍然较高。例如羊草和星毛萎陵菜的蒸腾速率在“午休”时仍很高,其气孔导度虽有下降,但仍然保持了一个相对高的值,再看糙隐子草,其“午休”时期的蒸腾速率和气孔导度也达到极大。陶汉之[6]对茶树(Melaleaac alterni folia)光合作用的研究发现,净光合速率和气孔导度的变化没有平行关系,不呈明显正相关。可以认为,在“午休”这段时间,植物的光合速率下降了,但气孔导度并没有明显下降,因为植物必须通过蒸腾作用散失水分降温来避免叶片受高温危害[7]。
3 结论与讨论
目前,对于草原植物光合作用特征已有一些研究[8-9],如杜菁昀等[8]对内蒙古草原植物光合作用进行研究时,依据所测植物光合速率的高低,可将其分为4个水平,星毛萎陵菜和糙隐子草属于第2个水平,即光合速率为 250~150[μ mol/(kg· s),CO2],而羊草属于第4个水平,即光合速率<50[μ mol/(kg·s),CO2],与本试验测得的结果相符,即代表低、中能值植物功能群的星毛委陵菜和糙隐子草的净光合速率要高于代表高能值植物功能群的羊草。而根据所测植物蒸腾速率的强弱,亦将其分为4个水平,星毛萎陵菜属于第2个水平,即蒸腾速率为(50 ×103)~(100 ×103)[μ mol/(kg·s),H2O],而糙隐子草和羊草属于第3个水平,即蒸腾速率为(25×103)~(50 ×103)[μ mol/(kg·s),H2O],这一变化规律基本符合本研究得到的不同能量功能群蒸腾速率变化的规律。
光合作用是植物十分复杂的生理过程,叶片光合效率既受自身因素的影响,又与外界因子密切相关。在自然界,植物光合作用在中午降低是一个普遍的现象,即光合午休,它是植物在长期进化过程中适应干旱环境而产生的一种生理现象,对于植物的节水与生存是有利的[10]。通常情况下,叶片光合速率日进程和太阳辐射强度的变化趋势相似,是一条单峰曲线,然而在高温低湿阳光充足的夏季晴天,许多植物的光合日变化表现为一条双峰曲线,上下午各有1个高峰,中午有1个低谷[11]。这在本研究所选植物光合日变化上得到了验证,并且关于羊草光合特性的研究显示:羊草叶片净光合速率的日变化在土壤质量含水量为(40±1)%、(20±1)%及(10±1)%的条件下呈双峰曲线,峰值分别出现在8:00和16:00,有明显的光合午休现象[12]。而本研究中羊草叶片的净光合速率除了8:00和16:00的峰值外,12:00也出现了峰值,出现了“三峰”,明显的光合午休现象则出现在14:00。说明试验过程中水分充足,同时也说明羊草本身能成为高能值植物,也因为其积极进行光合作用,同时消耗较少。也提示在实际提高植物能值或生产力的时候,除了选择本身具有较高生产力的植物外,也应该设法让植物的光合日进程变成三峰或更多峰,即如何避免植物的午休[13],提高单种植物的生产力,从而上升到功能群,提高整个草原生产力,使畜牧业发展。正是因为改善光合作用对于提高作物的产量潜力具有的重要意义,使之成为农学研究较为重视的课题之一[14]。
尺度上推是生态学研究中常用的方法,多植物种构成的群落的综合特性——类群的合并问题,即属于生态学里尺度上推问题之一。如何划分植物功能类群或功能型,才能在模型中准确地预测系统整体的行为,是生态系统模拟中基础但又十分重要的问题。这个问题的有效解决,可以为大尺度的景观动态模拟提供更加坚实的理论基础[15]。关于植物功能群的研究,朱玉杰等[16]有基于机理的气孔导度模型和光合模型参数拟合结果作为分类指标划分植物类群,评价了不同植物类群的植物生理生态特点,得出强抗旱中光合植物功能类群、中抗旱高光合植物功能类群、低抗旱低光合植物功能类群所具有的不同特点的结论。
更多的研究则是以不同的光合作用途径划分光合功能群,如C3和C4植物,在韩梅等[17]的研究中,大都发现在环境比较恶劣的时候,比如土壤盐碱化,大气中CO2浓度过低,C4植物的光合作用能力比C3植物强,在这种情况下从光合功能群的角度研究也许更适合。本研究选择的都是草原中数量多而分布极广的植物,所测量的植物中仅糙隐子草为C4植物,其余皆为C3植物[18],且本研究条件下CO2充足,所以进行C3和C4植物的区分意义不大。
如果说光合功能群反应的是植物的光合作用机理的差异,那么能量功能群反应的就是光合固能的结果,即植物具有的能量的状态的不同。本研究就是从植物能量出发探讨了3种不同能量功能群的光合作用的不同特点,并基于此探讨能量功能群一些生理和生态特征,研究结果显示不同能量功能群的植物显示出不同的光合作用特点。比较不同能量功能群的净光合速率,由低能值植物向高能值植物降低,从气孔导度和蒸腾速率看,低能值植物都较大,说明其光合生产力很强,生命活动旺盛,耗能也较大。再而从植物繁殖策略上分析,低能值植物多为一、二年生植物,选择了多繁殖后代以延续种群的方法[19],繁殖消耗较大,故而尽管其光合能力强,但自身能量水平仍较低。而高能值植物刚好相反,尽管其净光合速率相对较低,但由于其生长耗能和繁殖耗能都较少,反而自身能量积累较高。
能量功能群作为一种新的功能分类方法,在体现植物的功能特性上具有一定的特色和优势,从能量出发,也便于与草原植物生产力相连接,而草原植物生产力又是整个草原食物链的基础,直接关系到草原畜牧业的发展和可持续利用[20]。
[1]孙慧珍,国庆喜,周晓峰.植物功能型分类标准及方法[J].东北林业大学学报,2004,32(2):81-83.
[2]鲍雅静,李政海.基于能量属性的植物功能群划分方法探索——以内蒙古锡林河流域草原植物群落为例[J].生态学报,2008,28(9):4540-4541.
[3]顾德兴.普通生物学[M].北京:高等教育出版社,2000.
[4]陈佐忠.中国典型草原生态系统[M].北京:科学出版社,2000.
[5]许大全,沈允钢.光合作用的限制因素[A].余叔文,汤章城.植物生理学和分子生物学(第2版)[M].北京:科学出版社,1999:262-276.
[6]陶汉之.茶树光合日变化的研究[J].作物学报,1991(6):444-452.
[7]曾艳琼,卢欣石,杨世民.花椒树下苜蓿、百脉根和白三叶的光合生理生态特性研究[J].草业科学,2008,25(4):120-121.
[8]杜菁昀,杜占池,崔骁勇.内蒙古典型草原地区常见植物光合、蒸腾速率和水分利用效率的比较研究[J].草业科学,2003,20(6):12-13.
[9]闫瑞瑞,卫智军,运向军,等.放牧制度对短花针茅荒漠草原主要植物种光合特性日变化影响的研究[J].草业学报,2009,18(5):160-167.
[10]韩瑞宏,田华,张亚光,等.北京地区公农1号紫花苜蓿叶片光合作用日变化特征[J].草业科学,2008,25(6):37-38.
[11]杜占池,杨宗贵.羊草和大针茅光合作用午间降低与生态因子关系的研究[J].自然资源学报,1990,5(2):185-187.
[12]李林芝,张德罡,辛晓平,等.呼伦贝尔草甸草原不同土壤水分梯度下羊草的光合特性[J].生态学报,2009,29(10):5271-5273.
[13]唐微,朱名安,刘俊.C3、C4及CAM 植物的光合速率日变化及叶绿素含量的比较[J].湖北农业科学,2002(4):39-40.
[14]林茂兹,曹智,王阮萍,等.太子参光合速率变化特征初探[J].草业科学,2009,26(7):36-37.
[15]臧润国,张志东.热带森林植物功能群及其动态研究进展[J].生态学报,2010,30(12):3289-3291.
[16]朱玉洁,高琼,刘峻杉,等.基于气孔导度和光合模型的植物功能类群合并问题[J].植物生态学报,2007,31(5):880-882.
[17]韩梅,杨利民,张永刚,等.中国东北样带羊草群落C3和C4植物功能群生物量及其对环境变化的响应[J].生态学报,2006,26(6):1830-1831.
[18]Wang R Z.Photosynthetic pathway types in rangeland plant species from Inner Mongolia,North China[J].Photosynthetica,2004,42(3):339-344.
[19]尚玉昌.普通生态学[M].北京:北京大学出版社,2002.
[20]洪锐民.天然草地净初级生产力研究进展[J].草业科学,1993,10(5):31-33.
猜你喜欢
农业工程学报(2022年3期)2022-04-16
西南农业学报(2021年7期)2021-08-23
现代畜牧科技(2021年7期)2021-07-28
现代畜牧科技(2021年4期)2021-07-21
节水灌溉(2021年5期)2021-05-26
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
中国农业科技导报(2020年12期)2020-03-15
现代畜牧科技(2016年9期)2016-02-21
