
肝细胞核因子4α与糖尿病
2011-06-07 02:21润综述杨红英审校
实验与检验医学 2011年6期
马 润综述,杨红英审校
(昆明医学院第二附属医院,云南 昆明 650101)
肝细胞核因子(HNF)是一类异质的保守转录因子,参与调节肝脏内糖代谢、脂质代谢相关基因的表达。肝细胞核因子4α(HNF-4α)作为细胞核受体超家族成员之一,具有脂溶性激素受体的特性,能够直接进入胞核调节基因转录,它通常在肝脏、肾脏、肠及胰腺表达,从而参与调节胰岛β细胞和肝脏的发育、分化及功能,维持葡萄糖稳态[1]。HNF-4α是青少年发病的成年型糖尿病 (Maturity-Onset Diabetes of the Young,MODY)的致病基因,即MODY1基因。近年来国外大量的科学研究发现,HNF-4α基因多态性与2型糖尿病之间存在密切的关联。现就HNF-4α与糖尿病的关系作一综述。
1 HNF-4α的概述
HNF家族有6个成员,分别是HNF-1(包括HNF-1α 和 HNF-1β)、HNF-3(包括 HNF-3α、HNF-3β 和 HNF-3γ)、HNF-4(包括 HNF-4α 和 HNF-4γ)、CCAATP增强子结合蛋白 (CPEBP)、D2结合蛋白(DBP)和HNF-6。其家族的共同特点是(1)在肝脏中高水平表达;(2)含有进化上相当保守的DNA结合区;(3)在转录水平调节肝脏内基因的特异性表达;(4)转录因子间相互作用并形成网络系统——HNFs调节网络;(5)功能上的交叉表现为靶基因谱的部分交叉重叠,其靶基因包括载脂蛋白、脂肪酸及葡萄糖代谢相关基因等[2]。
肝细胞核因子4α(Hepatocyte nuclear factor-4α gene,HNF-4α)是核受体超家族配基激活型转录因子的高度保守成员之一,在分化成熟的肝细胞中高表达,是调控肝细胞分化和维护肝细胞生物学功能的重要转录蛋白,在肝脏的形成和成人肝脏许多基因的转录调节中起关键的作用[3],其基因结构大致
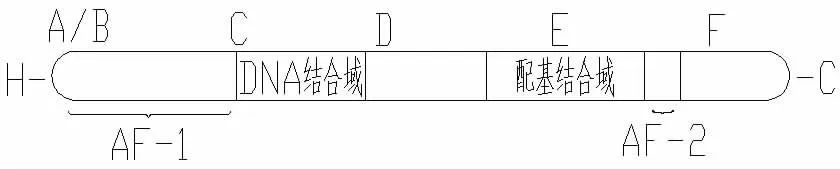
图1 HNF-4α的结构
可分为 A、B、C、D、E 和 F 区[4](见图 1)。 其中 A/B 区即氨基末端区,含有反式激活功能域AF-1,其功能与启动子特殊序列有关。C区具有高度保守的锌指结构,并可与D区形成铰链-DNA结合域(DBD)。DBD可与靶基因特殊DNA—序列激素反应元件(HREs)结合。E区是一个长疏水区,可形成配基结合域(LBD),并具有受体二聚体化和配基结合等功能。DBD和LBD均为保守区域,是核受体超家族的特征。E区还具有一个由7个氨基酸组成的核心序列即激活功能域AF-2。羧基端的F区可协助AF-2发生构相变化,从而易化AF-2与其他辅活化子或转录因子顺式反应元件的结合[5]。HNF-4α的表达受两个选择性启动子的调控,P1及P2,而在胰腺β细胞中P2启动子的转录则是主要的。
HNF-4α的功能主要通过两种方式来实现。一种是由于HNF-4α在肝、肠、肾及胰的表达,参与组织分化和能量代谢,从而调控机体的各种生理活动;另一种方式是通过与其他HNF分子共同组成在组织分化过程中所必需的转录因子调节网络——HNFs调节网络[6],调控胚胎发育过程中各种基因的表达,HNFs 调节网络系统包括 HNF-4α、HNF-1α、HNF-1β、HNF-3α 和 HNF-3β 等,它们在机体中相互依赖,形成交互调节的网络。
2 HNF-4α与MODY
MODY是一种高度异质性的单基因疾病,与β细胞功能中单基因缺陷相关。临床研究显示其发病与葡萄糖刺激的胰岛素不正常分泌有关[7],其胰岛素不足程度介于1型和2型糖尿病之间,临床表现具有两者的某些特点,构成了糖尿病疾病谱的中间过渡类型。临床特征是:(1)诊断糖尿病时年龄小于25岁,少年期往往无症状,仅在等应激状态下出现症状;(2)确诊后至少2年内不需要用胰岛素治疗;(3)一般无酮症酸中毒;(4)空腹血清C肽≥0.3nmol/L,葡萄糖刺激后≥0.6nmol/L;(5)累及3代以上的家族成员,呈常染色体显性遗传,而与HLA无关;(6)少年期MODY病人以非肥胖者居多;(7)可有大血管和微血管并发症。MODY患病率具有明显的种族差异,随着基因及候选基因克隆技术的发展,目前已发现至少有 6种 MODY致病基因,即 HNF-4α/MODY1 、葡 萄 糖激酶 (GCK)/MODY2、HNF-1α/MODY3、胰岛素启动因子1(IPF2-1)/MODY4、HNF-1β MODY5 及 (NeuroD1/BetaA2)/MODY6, 其 中HNF-4α基因突变所致的MODY1比较罕见(迄今只在13个家族中发现),患者无胰岛素抵抗(IR),但胰岛β细胞分泌功能受损[8-13]。
199 6 年发现MODY患者HNF-4α基因的第268位密码子可发生CAG(G)→TAG(Am)无意义突变,即Q268X突变型,认定HNF-4α是第一个与MODY相关的基因,即MODY1基因。近年来,在MODY1患者体内发现的突变有 13个[14,15]:无义突变2个(Q268X和R154X)、移码突变2个(F75f sdel T和K99f sdelAA)、插入突变1个(V328-329)和8个错 义 突 变 (G115S、R127W、T130I、V255M、E276Q、V393I、P441I和I454V),这些位点分别位于HNF-4α基因的反式激活域及DNA结合域,直接影响其与所调节基因的结合及转录调控。这些突变损伤HNF-4α功能的可能机制总结为:(1)无义突变使激活功能区域AF2激活作用缺失,如Q268X的突变使其失去了二聚体形成、再识别位点结合和转录激活功能,从而引起基因表达失控;(2)DNA结合区域突变使DNA结合能力丧失,如E276Q的突变,使其在体内不稳定,丧失了与DNA结合的能力,导致HNF-4α转录活性受损,影响胰腺β细胞代谢途径,从而诱发糖尿病的发生[16];(3)辅激活因子及其他转录激活因子募集受损。辅激活因子是作为核受体转录激活的桥梁,从而调节不同靶细胞的表达。腺病毒E1A相关蛋白(p300)是与HNF-4α相互作用的重要辅激活因子,而R154X、E276Q等基因的突变可使p300募集反应受损,从而影响HNF-4α的表达;(4)错义突变如G115S(Gly115--Ser)产生一个蛋白激活A(PKA)磷酸化位点[17],PKA介导的磷酸化使HNF-4α转录活性受损。HNF-4α是一种磷蛋白,其活性受磷酸化调节。酪氨酸磷酸化为HNF-4α核定位所必需。丝氨酸及苏氨酸对维持HNF-4α的正常功能发挥重要作用。cAMP依赖的PKA使位于A盒的丝氨酸磷酸化,导致HNF-4α DNA结合及转录活性丧失。
3 HNF-4α与2型糖尿病
糖尿病是一组由环境因素和遗传因素相互作用而产生的临床综合征,其中遗传因素在糖尿病,尤其在2型糖尿病的发病机制中扮演着重要角色。在双胞胎之一有2型糖尿病的情况下,若是同卵双生那么另一个患此病的机率为41%~55%;若是异卵双生,那么发病几率为10%~15%[18]。2002年5月15日《美国医学协会杂志(JAMA)》公布了一篇由凯撒医学机构调查的研究结果。调查显示,由于基因的关系,亚裔在各族裔当中,2型糖尿病的发病率高居榜首,发病率比其它族裔高出50%,首次从基因的角度解释2型糖尿病在不同族裔当中的发病率。该研究负责人卡特表示,遗传因素在2型糖尿病发病过程中起决定作用,族裔的基因差别被认为是2型糖尿病发病的重要原因。
国内外大量的科学研究发现,HNF-4α基因与2型糖尿病之间存在密切的关联。HNF-4α的变异,最先在MODY患者中被发现。早在1998年,Hani等[19]对法国糖尿病家系进行基因扫描时,发现HNF-4α基因标记位点与2型糖尿病连锁。HNF-4α基因第393位碱基发生错意突变,导致缬氨基酸变成异亮氨酸,该突变与糖尿病共分离,并与胰岛素分泌受损有关,体外表达时发现该突变物使胰岛素基因转录活性明显下降。随后Love-Gregory[20]的研究又发现,在这基因启动子区有一可变区域,这一区域与2型糖尿病有确切相关性。正是这一启动子区的多态性,导致了胰腺β细胞中的基因的剪接的变化。研究者在这一区域选取了9个SNPs(Single nucleotide polymorphisms,单核苷酸多态性)排列它们的单倍型,结果发现了在德国犹太人中4个与2型糖尿病密切相关的SNPs,即rs4810424、rsl884613、rsl884614 与 rs2144908。最近,Barroso等[21]对德系犹太人的研究也同样发现在HNF-4α启动子附近的rs1884613和rs2144908与2型糖尿病相关,Neuman等[22]的研究也验证了在这一人群中rs1884613与2型糖尿病相关。在芬兰,Silander等[23]用受累同胞对的分析方法,将2型糖尿病的易感基因定位于20q13.12-20q13.13,这一区域正好是HNF-4α所在的染色体区域,其中与2型糖尿病相关最显著的是位于P2启动子附近的rs2144908和位于外显子附近的rsl884613。而在日本,Hara等[24]的研究也证明了这一结论,Hara等发现,在日本人群中HNF-4α基因P2启动子区包含的两个SNPs(rs1884614和rs2144908)的单体型与2型糖尿病显著相关,相关程度随检测位点与P2启动子区的距离成反比。最近,Jafar-Mohammadi等[25]对1000多例英国2型糖尿病患者研究发现,T130I突变型(rs1800961)与2型糖尿病相关。在国内,LI等[26]对中国人群的研究中发现rs11574736与中国汉族人群患2型糖尿病相关。HNF-4α基因中的SNPs能增加糖尿病患病风险的机理可能是由于在胰腺中,SNPs影响了HNF-4α中的P2启动子,从而影响了HNF-4α的基因表达,进而引起了β细胞功能受损,导致糖尿病。
然而,对HNF-4α基因SNPs的研究还存在与上述截然相反的结论。Vaxillaire等[27]在法国人群中未证实 P2启动区附近的 4个 SNPs(rs2144908、rs6031552、rs2425637、rs1884614)与 2 型糖尿病的相关性。Winckler等[28]对瑞典、加拿大、芬兰等地区的研究也得出了阴性结果,未能证明HNF-4α基因的SNPs与2型糖尿病有关。最近,Muller等[29]研究了Pima印第安人HNF-4α基因启动子和编码区的19个SNPs,结果表明HNF-4α基因变异可能不是Pima 2型糖尿病的主要相关因素。在国内,Wang等[30]的研究发现rs2144908与北京汉族人群患2型糖尿病无关,Chen等[31]对近2000名中国汉族2型糖尿病病人的研究也发现rs184614和rs2425637与中国汉族人群患2型糖尿病无关。这种研究结果的不一致可能反映了2型糖尿病的异质性,在与2型糖尿病连锁的HNF-4α基因区20q12-20q13.1,不同的种群、不同的生活环境以及不同的生活习惯可能有不同的易感位点,也可能是由于研究对象来自2型糖尿病家系,导致过高评价群体发病率而造成的。
4 结束语
随着糖尿病的发病率逐年上升,遗传因素在2型糖尿病中的作用越来越受人们重视。HNF-4α自1996年被确定为MODY1的致病基因后,其在2型糖尿病中的作用也开始受到人们的关注。HNF-4α作为2型糖尿病的候选基因,现在发现的突变类型多在欧美地区分布,但由于族裔的差异可能存在尚未发现的变异类型。因此全面系统分析HNF-4α在我国人群中的遗传变异,研究在我国人群2型糖尿病发病中的作用,对预防和治疗2型糖尿病具有十分重要的临床意义。
[1]Gupta RK,Vatamaniuk MZ,Lee CS,et al.The MODYl gene HNF-4d regulates selected genes involved in insulin secretion[J].J Clin Invest,2005,115:1006-1015.
[2]Seufert J,Weir GC,Habener JF,et al.Differential expression of the insulin gene transcriptional repressor CCAAT/enhancer binding protein beta and transactivator islet duodenum homeoborl in rat pancreatic beta cells during the development of diabetes mellitus[J].J Clin Invest,1998,101:2528-2539.
[3]郑 洁,李 进,于树娜,等.肝细胞核因子的研究进展[J].医学综述,2008,14(4):491-493.
[4]Mangelsdorf DJ,Evans RM.The RXR heterodimers and orphan receptors[J].Cell,1995,83:841-850.
[5]Servitja JM,Ferrer J.Transcriptional networks controlling pancreatic development and beta cell function[J].Diabetologia,2004,47:597-613.
[6]Duncan SA,Navas MA,Dufort D,et al.Regulation of a transcription factor network required for differentiation and metabolism[J].Science,1998,281:692-695.
[7]苏 永,黄志勇,汪艳芳,等.2型糖尿病与肝细胞核因子-4α相关基因的研究[J].河南诊断与治疗杂志,2002,16(2):87-88.
[8]Yamagata K,Furuta H,Oda N,et al.Mutations in the hepatocyte nuclear factor-4α gene in maturity-onset diabetes of the young(MODYl)[J].Nature,1996,384:458-460.
[9]Froguel P,Zouali H,Vionnet N,et al.Familial hyperglycemia due to mutations in glucokinase: definition of a subtype of diabetes meltitus[J].N Engl J Med,1993,328:697-702.
[10]Yamagata K,Oda N,KaIsaki PJ,et al.Mutations in the hepatocyte nuclear factor-1α gene in maturity-onset diabetes of the young(MODY3)[J].Nature,1996,384:455-458.
[11]Stoffers DA,Ferrer J,CIarke WL,et al.Early-onset type-Ⅱdiabetes mellitus(MODY4)linked to IPFl[J].Nature Genet,1997,17:138-139.
[12]Horikawa Y,Lwasaki N,Hara M,et al.Mutation in hepatocyte nuclear factor 1B gene(TCF2)associated with MODY[J].Nature Genet,1997,17:384-385.
[13]Malecki MT,Jhala US,Antonellis A,et al.Mutation in NEUROD1 are associated with the development of type 2 diabetes mellitus[J].Nat Genet,1999,23:323-328.
[14]Navas MA,Munoz-Elias,Kim J,et al.Functional characterization of the MODY1 gene mutations HNF4(R127W),HNF4(V255M),and HNF4(E276Q)[J].Diabetes,1999,48:1459-1465.
[15]Taghavi SM,Fatemi SS,Rafatpanah H,et al.Mutations in the coding regions of the hepatocyte nuclear factor 4 alpha in Iranian families with maturity onset diabetes of the young[J].Cardiovascular Diabetology,2009,8:63.
[16]宋景春,李志超等.肝细胞核因子-4与MODY[J].国外医学内分泌学分册,2001,21(1):13-15.
[17]Oxombre B,Kouach M,Moerman E,et al.The G115S mutation associated with maturity-onset diabetes of the young impairs hepatocyte nuclear factor 4 alpha activities and introduces a PKA phosphorylation site in its DNA-binding domain [J].Biochem J,2004,383:573-580.
[18]周 莉,孙子林.2型糖尿病病因研究进展 [J].实用糖尿病杂志,2007,4(2):56-57.
[19]Hani EH,Suaud L,Boutin P,et al.A missense mutation in hepatocyte nuclear factor-4α,resulting in a reduced transactivation in human late-onset NIDDM[J].J Clin Invest,1998,101:521-526.
[20]Love-Gregory LD,Wasson J,Ma J,et al.A common polymorphism in the uptream promoter region of the hepatocyte nuclear factor-4α gene on chromosome 20q is associated with type 2 diabetes and appears to contribute to the evidence for linkage in an Ashenzai Jewish population[J].Diabetes,2004,53:1134-1140.
[21]Barroso I,Luan J,Wheeler E,et al.Population-specific risk of type 2 diabetes conferred by HNF4A P2 promoter variants[J].Diabetes,2008,57:3161-3165.
[22]Neuman RJ,Wasson J,Atzmon G,et al.Gene-Gene interactions lead to higher risk for development of type 2 diabetes in an Ashkenazi Jewish population[J].PLoS ONE,2010,5(3):9903.
[23]Silander K,Mohlke KL,Scott LJ,et al.Genetic variation near the hepatocyte nuclear factor-4α gene predicts susceptibility to type 2 diabetes[J].Diabetes,2004,53:1141-1149.
[24]Hara K,Horikoshi M,Kitazato H,et al.Hepatocyte nuclear factor-4alpha P2 promoter haplotypes are associated with type 2 diabetes in the Japanese population[J].Diabetes,2006,55:1260-1264.
[25]Jafar-Mohammadi B,Groves CJ,Gjesing AP,et al.A role for coding functional variants in HNF4A in type 2 diabetes susceptibility[J].Diabetologia,2011,54:111-119.
[26]Li T,Wu X,Zhu X,et al.Association analyses between the genetic polymorphisms of HNF4A and FOXO1 genes and Chinese Han patients with type 2 diabetes[J].Mol Cell Biochem,2011,353:259-265.
[27]Vaxillaire M,Dina C,Lobbens S,et al.Effect of common polymorphisms in the HNF4alpha promoter on susceptibility to type 2 diabetes in the French Caucasian population[J].Diabetologia,2005,48:440-444.
[28]Winckler W,Graham RR,de Bakker PI,et al.Associafion testing of variants in the hepatocyte nuclear factor 4alpha gene with risk of type 2 diabetes in 7,883 people[J].Diabetes,2005,54:886-892.
[29]Muller YL,Infante AM,Hansen RL,et al.Variants in hepatocyte nuclear factor 4alpha are modestly associated with type 2 diabetes in Pima Indians[J].Diabetes,2005,54:3035-3039.
[30]Wang F,Han X,Ren Q,et al.Effect of genetic variants in KCNJ11,ABCC8,PPARG and HNF4A loci on the susceptibility of type 2 diabetes in Chinese Han population[J].Chinese Medical Journal,2009,122(20):2477-2482.
[31]Chen Z,Zhang D,Liu Y,et al.Variants in hepatocyte nuclear factor 4α gene promoter region and type 2 diabetes risk in Chinese[J].Experimental Biology and Medicine,2010,235:857-861.
猜你喜欢
青少年科技博览(中学版)(2022年11期)2023-01-07
上海金属(2021年6期)2021-12-02
昆明医科大学学报(2021年3期)2021-07-22
汽车维修与保养(2021年8期)2021-02-16
生物学通报(2019年3期)2019-02-17
特种经济动植物(2019年9期)2019-01-08
数码世界(2018年1期)2018-12-23
天然产物研究与开发(2016年6期)2016-06-05
工业设计(2016年4期)2016-05-04
中国学术期刊文摘(2015年16期)2015-11-01
