
地理居群和地形对伊犁绢蒿遗传多样性的影响
2011-12-08 08:00侯钰荣安沙舟徐彩芹
草业科学 2011年1期
侯钰荣,安沙舟,刘 冬,王 卫,徐彩芹
(新疆农业大学草业与环境科学学院 新疆草地资源与生态重点实验室,新疆 乌鲁木齐 830052)
地理居群和地形对伊犁绢蒿遗传多样性的影响
侯钰荣,安沙舟,刘 冬,王 卫,徐彩芹
(新疆农业大学草业与环境科学学院 新疆草地资源与生态重点实验室,新疆 乌鲁木齐 830052)
对不同地理居群和不同地形的160株伊犁绢蒿(Seriphidiumtransiliense)个体进行SRAP分析,以期为伊犁绢蒿遗传资源的有效利用提供理论依据。试验结果表明,1)伊犁绢蒿种群的多态位百分率为78.43%;2)不同地理居群的伊犁绢蒿各居群间没有明显的分界点,说明地理距离差异对伊犁绢蒿遗传分化物质的影响较小;3)不同地形的伊犁绢蒿可以聚为2类,将阳坡和平原聚为一类,阴坡和丘陵聚为一类。由结果可见,地形对伊犁绢蒿遗传物质分化的影响较大。
地理居群;地形;遗传多样性;伊犁绢蒿
伊犁绢蒿(Seriphidiumtransiliense)是新疆北疆荒漠地区最主要的牧草资源,是我国野生牧草资源遗传育种和生物技术研究的重要物质基础,是生物多样性的重要组成部分。近年来,不少学者针对伊犁绢蒿的生物学特性、生育规律、生理生态特性及繁殖等进行了大量的研究,但有关伊犁绢蒿种群的分子生物学方面的研究较少。遗传多样性是指某一物种群体中的可遗传物质的差别程度[1]。目前,遗传多样性在农作物[2-5]、蔬菜[6-7]、果木[8-9]、花卉[10-11]、药材[12-13]、牧草[14-16]中的应用有大量文献报道,但对菊科植物关于遗传多样性方面的研究较少,仅在个别花卉品种和蒿属植物中有研究。蒿属植物以冷蒿(Artemisiafrigida)[17-18]、青蒿(A.carvifolia)[19]和蒌蒿(A.selengensis)[20]为例,其他植物鲜见有报道。
对植物种群的遗传分化和遗传多样性研究以往多采用ISSR方法[21],而相关序列扩增多态性(Sequence-related amplified polymorphism,SRAP)是一种新型的基于PCR扩增的标记系统,标记采用长17~18 bp的引物对开放阅读框(ORFs)进行扩增,因不同物种的内含子、启动子与间隔长度不等而产生多态性[22]。与其他分子标记相比,该标记有稳定可靠、多态性高、共显性高、产率中等;在基因组中分布均匀、易测序、引物具有通用性、操作简单、成本低廉等优点[23-24]。利用SRAP分子标记方法,对伊犁绢蒿在不同生境的遗传多样性进行了研究,初步从分子水平了解伊犁绢蒿种群对生境和地形的响应,为探讨伊犁绢蒿种群的分子水平对环境的生态适应性提供基础资料。
1 材料与方法
1.1供试材料的准备 本试验材料共计5个居群,160个个体。样地路线选择从伊犁河谷西部的察布查尔县开始,路经新源县、沙湾县、乌鲁木齐市,以奇台县为落脚点。为了取到具有代表性的样品,首先将阳坡、阴坡、平原和丘陵从地貌类型上划分开,在选择样地时,阳坡和阴坡都选择在山体明显地段,平原都选择在山前冲积扇处,而丘陵地带选择在地势起伏不大,基本分不出阴阳坡的地带上,且在每个地理居群的不同坡向(以实际地形为准,包括阳坡、阴坡、平原和丘陵),选择生长健康的植株10株,即同一生境共采样30株(除新源县为40株)。单株距离>10 m,采其绿叶及花蕾共5~10 g,记号标签,放入密封袋,保存在冰盒内(底部有冰袋,内部温度<5 ℃),带回实验室供试验分析。
1.2SDS-CTAB法DNA的提取 采用SDS-CTAB法提取叶片基因组DNA[25-28],提取的部分结果见图1。储存缓冲液配备如表1。

图1 伊犁绢蒿基因组DNA电泳图

类项 剂量及药品名溶液A50mL100mmol/LTris-HCl(pH值8.0)50mmol/LEDTA(pH值8.0)1mol/LNaCl2%SDS2%PvP40蒸馏水溶液B25mL100mmol/LTris-HCl(pH值8.0)50mmol/LEDTA(pH值8.0)1mol/LNaOH10%CTAB蒸馏水1%β-巯基乙醇20%CTAB
具体提取步骤如下:1)称取各样品0.2 g,放入研钵中,在液氮下迅速磨成粉末,将粉末转入50 mL Beckman离心管中,加入0.8 mL 65 ℃预热的CTAB缓冲液(A液),质量分数1%的β-巯基乙醇40~50 μL,用玻璃棒搅拌使其混匀。 2)在65 ℃下水浴加热50 min,期间摇匀2次。 3)水浴后,加5 mol/L KAc 300 μL,冰浴60 min平衡后12 000 r/min离心10 min,取上清液0.8 mL,加B液200 μL,均匀水浴20 min,室温冷却。 4)加入500 μL氯仿异戊醇(24∶1),轻轻倒转摇动10 min,平衡后12 000 r/mim离心5 min。重复2次。 5)取上清液500 μL(不能吸到分界面),加水冷的异丙醇500 μL,-20 ℃冰箱放置60 min以上,平衡后6 000 r/min离心5 min,弃上清。 6)沉于管底的DNA用70%乙醇洗涤2~3次,风干加ddH2O(双蒸水) 40 μL,加1/10体积3 mol/L NaCl(pH值5.2)20 μL,乙醇400 μL,-20 ℃放置10 min,6 000 r/mim离心,将DNA真空抽干。 7)将DNA沉淀溶于50~100 μL的TE缓冲液中, -20 ℃下保存备用。
从样品中提取的基因组DNA经过紫外分光光度计检测,将DNA浓度稀释至50 ng/μL,放入-20 ℃冰箱中保存备用。
1.3SRAP反应体系的建立 一个标准PCR反应体系包括: 0.2 mmol/L dNTPs, 1.5 mmol/L MgCl2,0.3 μmol/L 引物,1 U TaqDNA聚合酶,30 ng模板DNA,最后加ddH2O补充体积至25 μL。PCR 反应采用复性变温法:开始94 ℃变性5 min,之后的35个循环在94 ℃变性1 min,35 ℃退火1 min,72 ℃延伸1~2 min,最后72 ℃延伸5~7 min进行。4 ℃终止反应并保存。
1.4引物的筛选 从现有的97个引物中逐一进行筛选,最后选出21条扩增条带清晰,重复性较好且稳定的引物对伊犁绢蒿160个样品进行DNA扩增。上游和下游自由组合结果序号为18、24、27、38、41、42、49、72、79、80、81、82、84、87、89、107、108、110、117、119、120;选用的引物序列号如表2。

表2 用于SRAP分析的标准引物
1.5聚丙烯酰胺凝胶电泳检测
1.5.1溶液配制
1)30%非变性聚丙烯酰胺母液:丙烯酰胺为亚甲基双丙烯酰胺(29∶1)。丙烯酰胺290 g亚甲基双丙烯酰胺10 g 加ddH2O定容至1 000 mL (定性滤纸过滤4 ℃ 放置)放置备用。
2)5×TBE(pH值为8.0)母液: 54 g Tris-base, 27.5 g硼酸, 20 mL 0.5 mol/L EDTA,加ddH2O定容至1 000 mL放置备用。
3)质量分数10%(NH4)2S2O8液(APS,过硫酸铵):1 g(NH4)2S2O8定容至10 mL, -20 ℃放置备用。
4)质量分数1%Na2S2O3溶液100 mL:1 g Na2S2O3定容至100 mL,4 ℃放置备用。
5)固定/终止液配方1 L: 5 mL冰乙酸,100 mL无水乙醇,加ddH2O定容至1 000 mL 放置备用。
6)染色液1 L: 质量分数0.2%的 AgNO3溶液。
7)显色液1 L:质量分数1.4%的NaOH溶液使用前加入10.0 mL 37%甲醛。
8)终止显色液1 L:质量分数0.75% Na2CO3。
9)质量分数10%的非变性聚丙烯酰胺凝胶的配置:13.3 mL 30%非变性聚丙烯酰胺母液,10 mL 5×TBE,700 μL APS,50 μL TEMED(四甲基乙二胺),25.5 mL ddH2O。
1.5.2非变性聚丙烯酰胺凝胶检测 PCR产物上样量为3.0 μL;恒压电泳60~80 min。
1.5.3银染检测程序 1)电泳结束,用蒸馏水洗胶2~3 min,加固定液固定10 min,轻轻摇动,直至染料消失; 2)用蒸馏水洗胶2次,加入染色液染色20 min,过程中轻轻摇动;3)用蒸馏水洗胶1次,加入显色液中直至条带清晰出现即可;4)条带清晰后,将显色液倒出,加入0.75%Na2CO3终止显色;5)取出胶片用保鲜膜包裹,记录编号并照相,对每个引物在每个样品产生的条带进行统计记录,在相同迁移率位置上有DNA条带的记为“1”,无DNA条带的记为“0”。
1.6数据处理 试验数据用PopGene32软件完成遗传多样性供试样品多态性的检测统计分析(表3),用UPGMA进行聚类分析。

表3 SRAP引物配对明细表
注:“+”代表上游引物,“-”代表下游引物,“-”代表未配对。
2 结果与分析
2.1多态位点分析 21个引物对不同地理居群和不同地形的伊犁绢蒿的160个DNA样品进行扩增(图2),共扩增出153个条带,平均每个引物扩增出7.3个条带。其中,有120条重复性高、清晰的多态性条带,扩增片段的大小均在1 000 bp以下,多态位点的百分率为78.43%。不同的引物组合扩增的多态性带数明显不同,21对引物扩增的多态性带数范围从5条(me3+em2)到55条(me7+em6) 不等,而同一引物组合在不同群体扩增的多态性带数基本相同。

图2 引物18对样品49~96的扩增结果
2.2不同地理居群间遗传多样性聚类分析 伊犁绢蒿5个居群间的遗传聚类结果见图3,5个居群基本被聚为2组,一组为察布查尔县和奇台县,另一组为沙湾县、乌鲁木齐市和新源县,但从遗传距离来看,没有明显的分界点;说明地理距离差异对伊犁绢蒿遗传分化物质的影响较小。同时也说明,荒漠区降水量的细微差别对伊犁绢蒿种群的遗传特性影响不大。

图3 不同生境间伊犁绢蒿居群间聚类分析
2.3不同地形居群间遗传多样性聚类分析 不同地形的伊犁绢蒿聚为2类,将阳坡和平原聚为一类,阴坡和丘陵聚为一类(图4)。这表明,接受光照时间长、光照强度大、地表温度较高的阳坡和平原分化明显,而阴坡和丘陵地带接受光照和地表温度较相似,聚为一类。说明地形对伊犁绢蒿遗传物质的分化有重要影响。
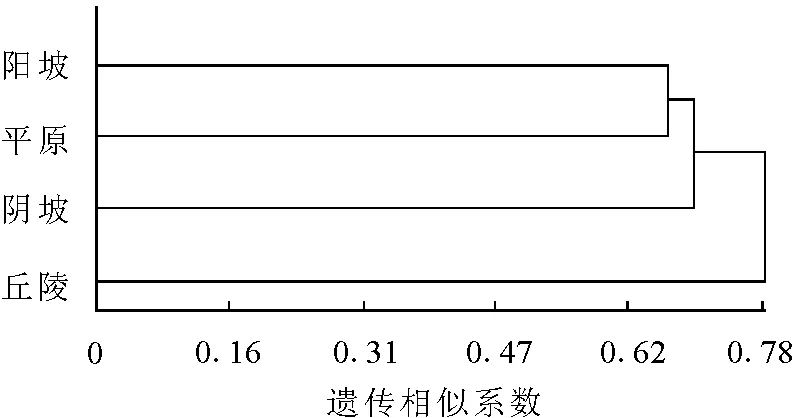
图4 不同地形间伊犁绢蒿居群间聚类分析
3 讨论与结论
伊犁绢蒿在分类上属于菊科植物,在新疆草地中主要出现在干旱和半干旱的荒漠区,生活型是小半灌木,实生苗是其种群更新的主要繁育系统,在群落的演替系列上,存在于演替的各个阶段,特别是在草地重度放牧下,由荒漠草地的建群种演替为伴生种或偶见种。伊犁绢蒿在时空变化上广泛分布,面临的环境差异也很大,个体和种群数目多,适应幅度大,基因交流频繁,因此维持遗传变异的能力强,产生新的遗传变异的机会也多,这都有利于其增加遗传多样性水平。Grant[29]、Millar和Libby[30]都认为一个物种的进化潜力和抵御不良环境的能力取决于遗传多样性水平的高低。因此,伊犁绢蒿较高的遗传多样性水平是其耐啃食、耐践踏、耐干旱、耐贫瘠、再生生长能力强的遗传基础。
通过SRAP分子标记对不同地理居群和不同地形的伊犁绢蒿天然居群的研究表明,伊犁绢蒿种群的多态位百分率为78.43%,随着生境的改变,伊犁绢蒿遗传多态位点数变化不大,地形的改变对伊犁绢蒿遗传多态位点数有一定影响,与其他采用SRAP标记进行检测的植物多态性百分率有差异,如新疆野苹果(Malussieversii)(P=98.56%)[21]、多花黑麦草(Loliummultiflorum)(P=77.09%)[31]、不结球白菜(Brassicapestrisssp.chinensis)(P=41.86%)[32]、苎麻(Boehmerianivea)(P=85.54%)[33]等。与之相比,伊犁绢蒿的遗传多样性水平属于中等。但与牧草相比,伊犁绢蒿遗传多样性具有相似性,这与牧草的生活特性也有一定的相关性。本研究表明,遗传距离和地理距离两者之间的关系不明显。这与Cui等[34]对松嫩草原羊草(Leymuschinensis)、穆立蔷和刘赢男[35]对紫椴(Tiliaamurensis)种群的研究结果基本一致,均显示遗传距离与地理距离间不存在显著的相关性。遗传距离受地形因素影响鲜见有报道。
目前,由于在不同的种、不同个体间具有一定得多态性SPAP技术已经成功用于种质资源的鉴定与评价工作[36]。对同一植物材料在分子标记方法上进行遗传多样性和形态学对比已有大量报道,尤其在蔬菜方面更是成熟,结果均表明SRAP标记比AFLP、RAPD、SSR等方法更能提供优良信息。但关于牧草方面的研究还远远不够,有待于深入研究。
[1] 丁玲,陈发棣,房伟民.菊属8个种27份材料遗传多样性的同工酶分析[J].西北植物学报,2007,27(2):249-256.
[2] 胡根海,胡海燕.用SSR标记分析高油栽培大豆品种的遗传多样性[J].江西农业大学学报,2008,30(4):638-642.
[3] 姜伟,朱宏波,何觉民.不同来源棉花种质资源遗传多样性的ISSR分析[J].棉花学报,2008,20(5):348-353.
[4] 陈志石,陈夕军,熊如意,等.江苏水稻品种抗稻瘟病遗传多样性研究[J].南京农业大学学报,2008,31(3):77-80.
[5] 李武,倪薇,林忠旭,等.海岛棉遗传多样性的SRAP标记分析[J].作物学报,2008,34(5):893-898.
[6] 黄如葵,孙德利,张曼,等.苦瓜遗传多样性的形态学性状聚类分析[J].广西农业科学,2008,39(3):351-356.
[7] 洪雨顺,杨德.辣椒种质资源遗传多样性保护和利用研究进展[J].中国农学通报,2006,22(2):358-360.
[8] 郭传友,黄坚钦,王正加,等.山核桃种质资源遗传多样性的RAPD分析[J].激光生物学报,2007,16(4):405-410.
[9] Chen X S,Feng T,Zhang Y M.Genetic diversity of volatile components in Xinjiang wild apple (Malussieversii)[J].Journal of Genetics and Genomics,2007,34(2):171-179.
[10] 韩洁,胡楠,李玉阁,等.菊花品种资源遗传多样性的AFLP分析[J].园艺学报,2007,34(4):1041-1046.
[11] 缪恒彬,陈发棣,赵宏波,等.应用ISSR对25个小菊品种进行遗传多样性分析及指纹图谱构建[J].中国农业科学,2008,41(11):3735-3740.
[12] 徐文斌,郭巧生,王长林.药用菊花遗传多样性的RAPD分析[J].中国中药杂志,2006,31(1):18-21.
[13] 葛淑俊,李广敏,马峙英,等.甘草野生种群遗传多样性的AFLP分析[J].中国农业科学,2009,42(1):47-54.
[14] 曾兵,张新全,范彦,等.鸭茅种质资源遗传多样性的ISSR研究[J].遗传,2006,28(9):1093-1100.
[15] 卢萍,赵萌莉,韩国栋,等.内蒙古西部地区砂珍棘豆遗传多样性分析[J].内蒙古农业大学学报,2007,38(2):160-165.
[16] 郭海林,刘建秀,周志芳,等.结缕草属植物种间关系和遗传多样性的SSR标记分析[J].草地学报,2008,16(6):552-558.
[17] 王静,杨持,尹俊,等.冷蒿种群在放牧干扰下遗传多样性的变化[J].生态学报,2004,24(11):2464-2471.
[18] 宛涛,蔡萍,张辰波,等.内蒙古不同生态地区冷蒿居群遗传多样性的RAPD分析[J].西北农林科技大学学报,2008,36(2):165-174.
[19] 陈大霞,李隆云,瞿显友,等.青蒿种质资源遗传多样性的SRAP分析[J].分子植物育种,2007,5(6S):52-56.
[20] 李双梅,郭宏波,黄新芳,等.蒌蒿DNA提取、RAPD优化及引物筛选初报[J].农业生物技术科学,2006,22(4):78-80.
[21] 张春雨,陈学森,林群,等.新疆野苹果群体遗传结构和遗传多样性的SRAP分析[J].园艺学报,200936(1):7-14.
[22] 路娟,张绍铃,刘庆忠,等.樱桃SRAP-PCR 体系优化及其遗传多样性分析[J].果树学报,2009,26(2):163-169.
[23] Li G,Quiros C F.Sequence-related amplified polymorphism (SRAP),A new marker system based on a simple PCR reaction:its application to mapping and gene tagging inBrassica[J].Theory Application Genetic,2003,107(1):168-180.
[24] 谭碧,王源秀,徐立安.杨树基因组SRAP扩增体系的建立与优化[J].应用研究,2009,23(2):25-29.
[25] Nei M,Li W H.Mathematical model for studying genetic variation in terms of restriction endonucleases[J].Proceeding of National Academy of Science of the USA,1985,76:5269-5273.
[26] Keim P,Shoemaker R C.A rapid protocol for isolating soybean DNA[J].Soybean Genetics Newsletter,1988,15(1):150-152.
[27] 陈旭辉,高玉葆,朱敏杰,等.小叶锦鸡儿基因组DNA的提取及AFLP反应体系的建立[J].植物研究,2009,29(5):529-533.
[28] 徐秋华,张献龙,聂以春,等.我国棉花抗枯萎病品种的遗传多样性分析[J].中国农业科学,2002,35(3):272-276.
[29] Grant V.The Evolutionary Process:A Critical Study of Evolutionary Theory[M].New York:Columbia University Press,1991:102-111.
[30] Millar C I,W J Libby.Strategies for conserving clinal,ecotypic,and disjunct population diversity in widespread species[A].In:Falk D A,Holsinger K E.Genetics and Conservation of Rare Plants[C].New York: Oxford University Press,1991:149-170.
[31] 季杨,张新全,马啸,等.多花黑麦草品种( 系) 间杂交及其杂种后代SRAP遗传分析[J].草业学报,2009,18(4):260-265.
[32] 单晓政,侯喜林,李英,等.不结球白菜SRAP体系优化与品种聚类分析[J].江苏农业学报,2009,25(3):610-615.
[33] Liu L J,Peng D X,Wang B.Genetic relation analysis on RamieBoehmerianivea(L.)Gaud.inbred lines by SRAP markers[J].Agricultural Science in China,2008,7(8):944-949.
[34] Cui J Z,Zu Y G,Nie J L,etal.Genetic differentiation ofLeymuschinensispopulations in Songnen grassland[J].Bulletin of Botanical Research,2001,21:116-215.
[35] 穆立蔷,刘赢男.不同地理分布区紫椴种群的遗传多样性变化[J].植物生态学报,2007,31(6):1190-1198.
[36] 陈福.新型标记SRAP及其在草业研究中的应用前景[J].草业与畜牧,2007(7):1-4.
InfluenceofdifferentlygeographicalpopulationsandterrainsongeneticdiversityofSeriphidiumtransiliense
HOU Yu-rong, AN Sha-zhou, LIU Dong, WANG Wei, XU Cai-qing
(College of Grassland and Environment Science, Xinjiang Agricultural University; Key Laboratory of Grassland Resources and Ecology of Xinjiang, Xinjiang Urumqi 830052, China)
The 160 plants ofSeriphidiumtransiliensewith the different geographical population growing at different terrains was used to screen the genetic diversity by SRAP technical analysis for providing the useful information for utilization this plant resource. The results of this study showed the percentage of polymorphism loci ofS.transiliensepopulationwas 78.43. This study also indicated that the obvious demarcation point was not found among the different geographical population ofS.transiliense, implying that the difference in geographical distance betweenS.transiliensepopulations did not affect the hereditary substance ofS.transiliense. However, 160 plants were clustered into two groups, and one group consisted of sunny slope and plain populations, the other group consisted of shady slope and hilly populations, indicating that terrain showed a greatly effect on the hereditary substance ofS.transiliense.
geographical population; terrains; genetic diversity;Seriphidiumtransiliense
S540.3;Q948
A
1001-0629(2011)01-0094-06
2010-02-23 接受日期:2010-04-27
教育部高等学校博士学科点专项科研基金项目(20096504110002);新疆草地资源与生态重点实验室开放课题(XJDX0209-2007-05)
侯钰荣(1982-),女,新疆玛纳斯人,在读博士生,从事草地资源与生态研究。
E-mail:houyurong0994@sina.com
安沙舟 E-mail:xjasz@126.com
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26
世界科学技术-中医药现代化(2022年3期)2022-08-22
浙江中医药大学学报(2021年6期)2021-07-12
中国粮油学报(2020年12期)2021-01-09
草地学报(2018年5期)2018-11-07
散文诗(2017年15期)2018-01-19
连环画报(2017年1期)2017-07-13
西南农业学报(2016年6期)2016-04-16
大众考古(2015年1期)2015-06-26
中央民族大学学报(自然科学版)(2015年1期)2015-06-11
