
珍稀濒危植物堇叶紫金牛生存群落结构特征及物种多样性
2012-05-30 03:29夏国华闫道良谢文远严彩霞吴家森李根有
浙江农林大学学报 2012年4期
马 凯,夏国华,闫道良,谢文远,严彩霞,吴家森,李根有
(1.浙江农林大学 风景园林与建筑学院,浙江 临安 311300;2.浙江农林大学 林业与生物技术学院,浙江 临安 311300;3.浙江省森林资源监测中心,浙江 杭州 310020)
群落结构特征及物种多样性研究一直是生态学研究的热点问题[1-2]。群落在组成和结构上表现出的多样性是考察群落中各种生物在空间和时间上的配置状况,甚至是结构、功能状态的基础,也是生物多样性研究中至关重要的方面[3],对研究群落原始生产力和环境条件资源都有重要的价值[4-5]。群落结构包括形态方面和生态方面的结构,它体现群落中不同个体的组配情况和更新机制,并能反映与环境之间的相互适应关系。种群的径级结构和高度级是群落结构的重要指标,对阐明群落的形成及其稳定性乃至动态变化与演替趋势都有重要的意义。开展对濒危植物群落结构等研究,有助于了解群落物种组成的生态特性、更新对策和动态,也可以为濒危物种保育的制定提供依据[6-8]。堇叶紫金牛Ardisia violacea,又名裹堇紫金牛,属紫金牛科Myrsinaceae紫金牛属Ardisia植物,特产于浙江和台湾,对研究中国大陆和台湾植物区系演化具有重要的科研价值。世界自然保护联盟将它列为稀有种,浙江省和台湾省则列为珍稀濒危植物[9]。堇叶紫金牛兼具紫金牛属锯齿组Sect.Bladhia和圆齿组Sect.Crispardisia的特征,可能是这2个类群之间的过渡类型,对紫金牛属植物的分组以及系统发育具有较高的研究价值。该种以其株型紧凑、叶绿果艳,姿态优美,具有较高观赏价值,是一种优良的盆栽植物,仅分布在浙江省杭州云栖、台湾北部乌来、桃宜公路巴棱至四棱山区[10]。最近在浙江舟山白泉乡、建德绿荷塘(最近调查资料,未发表)以及台湾巴福越、尖石、北横乡均有少量发现。堇叶紫金牛现有的研究仅局限于分类学研究,对其生境特点、种群结构、群落特征、自然更新机制等方面均尚未有报道。本研究通过对浙江省建德市绿荷塘森林公园中堇叶紫金牛生存群落的区系组成、外貌、结构和演替趋势等特征的研究,有助于探讨堇叶紫金牛的适生环境,从而采取适当的保护措施达到保护的目的,同时也为其他濒危植物的保护提供新的途径,为其群落和种群的合理保护提供理论依据。
1 研究地点及其自然概况
研究地位于浙江省建德市绿荷塘森林公园, 29°25′45″N,119°28′24″E,年平均气温为 16.7℃,最冷月(1月)平均气温为4.9℃,极端最低气温-13.9℃,最热月(7月)平均气温为28.1℃,极端最高气温42.9℃,年降水量为1604mm,平均相对湿度为69.6%,年日照时数为1762 h,全年无霜期为261 d,属北亚热带季风性气候。灌草植被丰富,森林覆盖率达76.7%,分布有典型的亚热带常绿阔叶林和针阔混交林。土壤为中亚热带山地红黄壤,土壤成土时间长,土层深厚,黏质粗松,酸碱度为pH 5.0~5.5,有机质和氮、磷、钾等元素含量丰富。
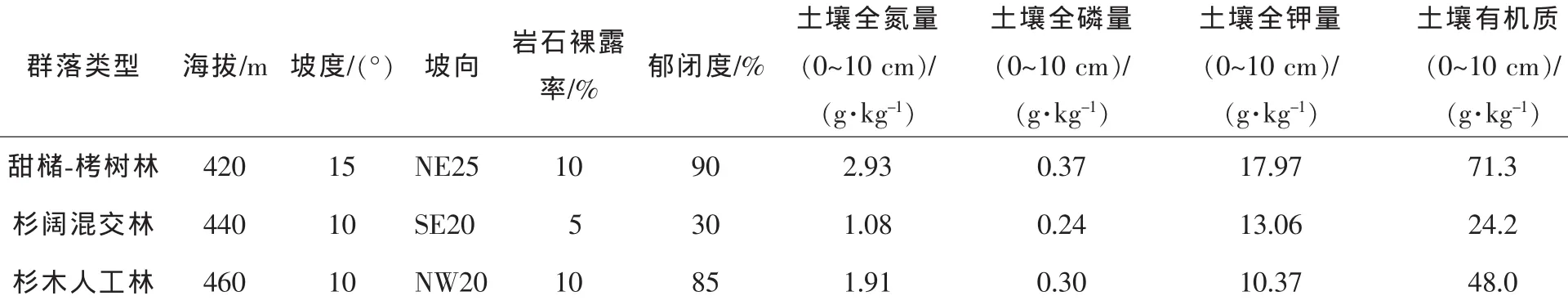
表1 堇叶紫金牛生存群落基本情况Table1 Environmental conditions of survival community of Ardisia violacea
2 研究方法
2.1 样地的设置和调查
根据堇叶紫金牛分布特点和资源总量等确定调查样地,对其分布地点浙江省建德市绿荷塘森林公园进行样方调查。根据堇叶紫金牛在不同斑块面积的大小,采用永久样方法,确定样方面积。前人研究证明:永久样方法在群落动态方面能够得到最直接和准确的数据[11],共设置甜槠-栲树林(20 m×20 m),杉阔混交林(80 m×5 m),杉木人工林(80 m×5 m)3个样地,分别记录样地的海拔、坡度、坡向、岩石裸露率、郁闭度,再将每个样地划分成5 m×5 m的相邻小格子进行统计分析,对样地内所有植物种类进行调查,记录乔木层(高5 m以上,胸径2.5 cm以上)个体的胸径、高度、冠幅,以及灌木、草本的高度、盖度等指标,并记录群落的生态学特征和周围生境。
2.3 数据处理分析
2.3.1 重要值的计算[13]乔木重要值=(相对密度+相对频度+相对优势度)×100。灌木重要值=(相对多度+相对频度+相对盖度)×100。其中,相对密度和相对多度为某种植物的个体数目与样方中全部植物的个体数目的比值;相对频度为某种植物出现的样方数占总样方数的比例,它是反映某种植物分布均匀程度的一个指标;相对优势度为某种植物的胸高断面积总和与样方内所有物种的胸高断面积总和的比值,能反映物种在群落中的优势程度。
2.3.2 多样性测度方法 采用下述方法测定堇叶紫金牛生存群落物种多样性[12-13]。公式分别为:①Pielou均匀度指数 J=-∑PilnPi/lnS。 ②Shannon-Wiener指数 H′=-∑PilnPi。 ③Simpson 指数 D=N(N-1)/∑ni(ni-1)。其中:N为物种总个体数;ni是第i种的个体数;Pi是第i种的个体数占总个体数的比例;S为样地中物种的总数。
2.4 高度结构和径级结构划分
根据堇叶紫金牛生存群落内各植物种群的高度特点,划分为5个等级,第1高度级为h≤5 m,每个等级间隔5 m,第2高度级为5 m<h≤10 m,群落内最大高度级h>20 m。在径级结构划分时,即以立木径级代替年龄进行分析[14-15],划分为 10个径级,第1径级胸径(DBH)≤5 cm,每个径级间隔5 cm,即5 cm<DBH≤10 cm为第 2径级,10 cm<DBH≤ 15 cm为第 3径级,…,群落内最大高度级为45<DBH≤50 cm。
3 结果与分析
3.1 群落种类组成
分析群落种属组成和地理成分是认识群落区系特征和生物多样性的首要基础。在对堇叶紫金牛生存群落样地调查中,根据调查资料统计,共有维管束植物116种(含变种),隶属于47科79属(表2)。其中蕨类植物6科7属9种,裸子植物2科2属2种,被子植物39科70属105种。被子植物中单子叶植物4科9属12种,双子叶植物35科61属93种。其中含有较多的属为禾本科(Gramineae,6属),山茶科(Theaceae,4属), 茜草科(Rubiaceae,4属), 豆科(Leguminosae, 4属), 樟科(Lauraceae,3属), 蔷薇科(Rosaceae,3属),壳斗科(Fagaceae,3属),马鞭草科(Verbenaceae,3属);含有种最多的是杜鹃花科(Ericaceae, 7种),壳斗科(Fagaceae, 7种)和樟科(Lauraceae, 7种), 山茶科(Theaceae, 6种),禾本科(Gramineae, 6种)。
从生活型上区分来看,堇叶紫金牛生存群落中共出现木本植物83种。其中针叶树2种,常绿阔叶树48种,落叶阔叶树33种。针叶树有马尾松Pinus massoniana和杉木Cunninghamia lanceolata,常绿阔叶树主要有山茶科、壳斗科、樟科、杜鹃花科和冬青科,占总数的57.83%。山茶科主要由隔药柃Eurya muricata,厚皮香Ternstroemia gymnanthera,木荷Schima superba,毛花连蕊茶Camellia fraterna,窄基红褐柃E.rubiginosa var.attenuata,油茶Camellia oleifera等组成。壳斗科主要由苦槠Castanopsis sclerophylla,米槠Castanopsis carlesii,甜槠Castanopsis eyrei,栲树Castanopsis fargesii,乌楣栲Castanopsis jucunda,石栎Lithocarpus glaber,青冈Cyclobalanopsis glauca等组成。樟科主要由乌药Lindera aggregata,刨花楠Machilus pauhoi,浙江樟Cinnamomum chekiangense,樟树Cinnamomum camphora等组成。杜鹃花科主要由马银花Rhododendron ovatum,乌饭树Vaccinium trichocladum,江南越橘Vaccinium mandarinorum,黄背越橘Vaccinium iteophyllum,短尾越橘Vaccinium carlesii等组成。冬青科主要由冬青Ilex purpurea,大果冬青Ilex macrocarpa,尾叶冬青Ilex wilsonii,厚叶冬青Ilex lmerrilliana等组成。落叶阔叶树种多为阳性树种,占到了总数的39.76%。如樟科的山胡椒Lindera glauca和山鸡椒Litsea cubeba,安息香科的拟赤杨Alniphyllum fortunei,郁香安息香Styrax odoratissimus,茜草科的白马骨Serissa japonica,金缕梅科的枫香Liquidambar formosana,五加科的棘茎楤木Aralia echinocaulis,忍冬科的宜昌荚蒾Viburnum erosum,马鞭草科的老鸦糊Callicarpa giraldii等。

表2 堇叶紫金牛生存群落维管束植物区系统计Table2 Satistics of the vascular plants of survival community of Ardisia violacea
根据Raunkiaer生活型分类系统划分(表3)[16-17],堇叶紫金牛生存群落中不含有大高位芽植物,中高位芽植物9种,小高位芽植物30种,矮高位芽植物31种,地上芽植物21种,地面芽植物4种,地下芽植物5种,1年生植物6种。该类群落以小高位芽植物和矮高位芽植物的种类最多,共61种,占总数的66.04%,说明此群落中高位芽植物占据绝对优势,其中地上芽植物、地面芽植物、隐芽植物分别占19.81%,3.77%,4.72%,而1年生植物占5.66%。由此可见大高位芽植物、1年生植物、隐芽植物和地面芽植物都比较少,反映了堇叶紫金牛样地环境为温暖多湿的特点,这一生活型组成与中国亚热带地区多数常绿阔叶林存在着普遍相似性[18]。
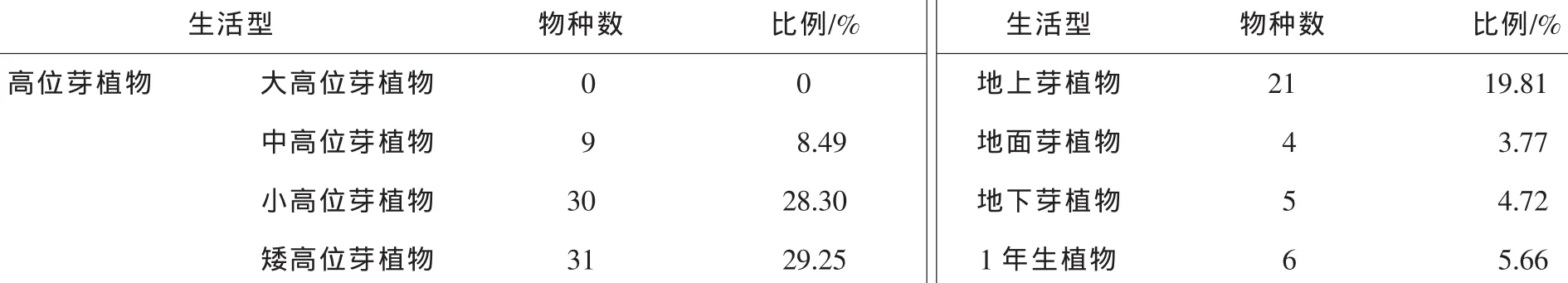
表3 堇叶紫金牛生存群落生活型谱Table3 Life-form spectrum of survival community of Ardisia violacea
3.2 群落类型特征
堇叶紫金牛生存群落生境存在一定的异质性,因此各样地的物种多样性有一定的差异,但变化幅度较小。根据其分布生境、群落结构和种类组成,可将堇叶紫金牛生存群落分为3个类型,即甜槠-栲树林、杉阔混交林和杉木人工林。
3.2.1 甜槠-栲树林 该类型主要分布在海拔420 m的山坡常绿阔叶林中,土层疏松肥厚。群落的特征和优势度明显。群落外貌暗绿色,林冠浑圆,郁闭度可达0.9,因此光照条件差,有机质含量丰富。堇叶紫金牛仅能够在林下林窗、林缘形成群聚。群落垂直结构可以分为草本层、灌木层、乔木下层、乔木上层,另外还有一些层间植物。乔木上层高14 m以上,主要树种有乌楣栲,栲树,野漆Taxicodendron succedaneum,青冈,拟赤杨,黄连木Pistacia chinensis,山合欢Albizia kalkora,无患子Sapindus mukorossi等,盖度为60%~80%,它们组成了群落的林冠层。乔木下层高度在8~15 m,主要组成种除了乔木上层的优势种类外,还有甜槠,苦槠,米槠,山矾Symplocos sumuntia,郁香安息香,大青Clerodendrum cyrtophyllum,冬青,大果冬青,石栎,黄檀Dalbergia hupeana,豹皮樟Litsea coreana var.sinensis和山鸡椒等,该层盖度为50%~70%。灌木层主要由乔木幼苗幼树和灌木组成,盖度为30%~50%。主要组成种有赤楠Syzygium buxifolium,乌药,窄基红褐柃,毛花连蕊茶,檵木Loropetalum chinense,宜昌荚蒾,八角枫Alangium chinense,石斑木Raphiolepis indica,乌饭树,朱砂根Ardisia crenata等。林下阴湿,光线弱,草本层发育不良,数量较为匮乏,物种比较单一,主要分布在林窗、林冠边缘,盖度在15%~30%左右,主要由阔鳞鳞毛蕨Dryopteris championii,石韦Pyrrosia lingua和薹草类Carex spp.等组成。层间植物不甚明显,主要由络石Trachelospermum jasminoides,香花崖豆藤Millettia dielsiana,缘脉菝葜Smilax nervo-marginata,土茯苓Smilax glabra和大叶白纸扇Mussaenda shikokiana等组成。
3.2.2 杉阔混交林 该群落主要分布在海拔440 m左右的山脊线上,地处防火带。群落外貌黄绿色,人为经营活动影响较大,乔木层和灌木层均不发达,林下环境相对较为宽松,仅在杉阔混交林与杉木人工林和常绿阔叶林带交界处略见乔、灌木分布,郁闭度在30%左右,因此有足够的阳光照射到该林下,满足了堇叶紫金牛生长对光照、水分、温度和养料等微环境的需求,因此堇叶紫金牛在该群落分布量十分丰富。该类型分层不明显,但草本层非常丰富。主要组成种有淡竹叶Lophatherum gracile,荩草Arthraxon hispidus,阔鳞鳞毛蕨Dryopteris championii,黑足鳞毛蕨Dryopteris fuscipes,青绿薹草Carex leucochlora,星宿菜Lysimachia fortunei,麦冬Ophiopogon japonicus,狗脊Woodwardia japonica,三脉紫菀Aster ageratoides,华泽兰Eupatorium chinense,早落通泉草Mazus caducifer,博落回Macleaya cordata,狭叶香港远志Polygala hongkongensis var.stenophylla,日本金星蕨Parathelypteris japonica等。
3.2.3 杉木人工林 该类型分布在海拔460 m左右的山坡上,生境较为潮湿,土壤较深厚肥沃,为杉木的适生环境,杉木与常绿阔叶树组成混交林,凋落物层深厚。群落外貌深绿色与褐绿色相间,种类组成较复杂。群落郁闭度可达0.8,有机质适中,但杉木凋落物分解慢,枯枝覆盖层厚,严重影响了堇叶紫金牛种子的萌发与扩展,因此堇叶紫金牛只能在杉木人工林和杉阔混交林交界处(约1 m处)形成大量群聚。群落垂直结构可以分为草本层、灌木层、乔木下层、乔木上层,另外还有一些层间植物。乔木上层高14 m以上,组成树种以杉木占优势,其他物种有乌楣栲,栲树,枫香Liquidambar formosana,拟赤杨和野漆等中高位芽树种,盖度可达50%~70%。乔木下层在4~6 m,主要树种除了上层主要优势树种外,还包括甜槠,木荷Schima superba,光叶石楠Photinia glabra,石栎,白栎Quercus fabri,青冈Cyclobalanopsis glauca,刨花楠Machilus pauhoi,山鸡椒,棘茎楤木和郁香安息香等物种。灌木层种类较多,主要有乌药,赤楠,隔药柃,山矾,毛花连蕊茶,窄基红褐柃,老鸦糊,朱砂根,紫金牛Ardisia japonica,杜茎山Maesa japonica,山莓Rubus corchorifolius,老鼠矢Symplocos stellaris,江南越橘,白马骨和卵叶帚菊Pertya scandens等组成。该层盖度为30%~50%。草本层优势种有柔枝莠竹Microstegium vimineum,芒萁 Dicranopteris pedata,芒 Miscanthus sinensis,求米草 Oplismenus undulatifolius,荩草,星宿菜,青绿薹草,华泽兰,早落通泉草,棕叶狗尾草Setaria plicata,狭叶香港远志,淡竹叶,阔鳞鳞毛蕨,黑足鳞毛蕨,狗脊,日本金星蕨,三脉紫菀和博落回等组成。层间植物包括络石,香花崖豆藤,菝葜Smilax china,藤葡蟠Broussonetia kaempferi和广东蛇葡萄Ampelopsis cantoniensis等。
3.3 群落垂直结构特征
垂直结构是反映群落结构重要的因素之一,群落的垂直结构决定于各种植物种的高度及在不同高度植物的数量[3]。植株的高度级概率分布反映了树种在森林中的各种特征(如耐荫性、更新类型等)以及群落的分层结构。
由图1得:3个群落类型的高度(h)结构大致相同,h≤5 m,5 m<h≤10 m,10 m <h≤15 m,h>20 m高度级的个体数目依次减少。h≤5 m高度级的个体数目在3个群落类型中都占有很大比例,尤其是在甜槠-栲树林和杉阔混交林中均达到70%以上,5 m<h≤10 m,10 m<h≤15 m,h>20 m只占到20%以上。可以看出,相对于甜槠-栲树林和杉阔混交林,杉木人工林的平均高度更高。

图1 群落高度结构Figure1 Height structure of the community
3.4 群落数量特征
3.4.1 甜槠-栲树林乔木种群数量特征 对甜槠-栲树林乔木种群特征进行分析,重要值处于前5位的分别是石栎、乌楣栲、青冈、甜槠和郁香安息香,重要值分别为75.8653,65.5884,57.8072,39.7549和25.0715,其重要值之和高达264.0873(表4),占到该群落乔木树种的88.0%。说明这5个种群在甜槠-栲树林乔木种群中占有绝对优势地位。
3.4.2 杉阔混交林群落乔木种群数量特征 对杉阔混交林乔木树种进行统计,重要值前3位的分别是杉木、乌楣栲和甜槠,其重要值分别为80.5389,62.7415和47.7535,且3个树种的重要值之和达到191.0339,说明杉阔混交林群落乔木种群的优势树种为杉木、乌楣栲和甜槠。而栲树、石栎、枫香和木荷的重要值也分别达到了23.1727,20.8728,20.8410和14.9537(表5),说明这4个树种在杉阔混交林乔木种群中也占有一定的地位。
3.4.3 杉木人工林乔木种群数量特征 对杉木人工林群落内的乔木种群数量特征进行分析,乔木种群中,杉木的重要值最高,达到83.2397,远远高于第2位甜槠的46.1250,在乔木种群中处于绝对优势地位。处于第3~6位的分别是枫香、乌楣栲、石栎和栲树。它们的重要值分别为37.0305,34.5711,27.2728和26.6793,这4个种群的重要值之和达到125.5537,与前2个种群的重要值之和仅相差3.8110(表6),说明这4个种群在杉木人工林群落内也占有一定的地位。
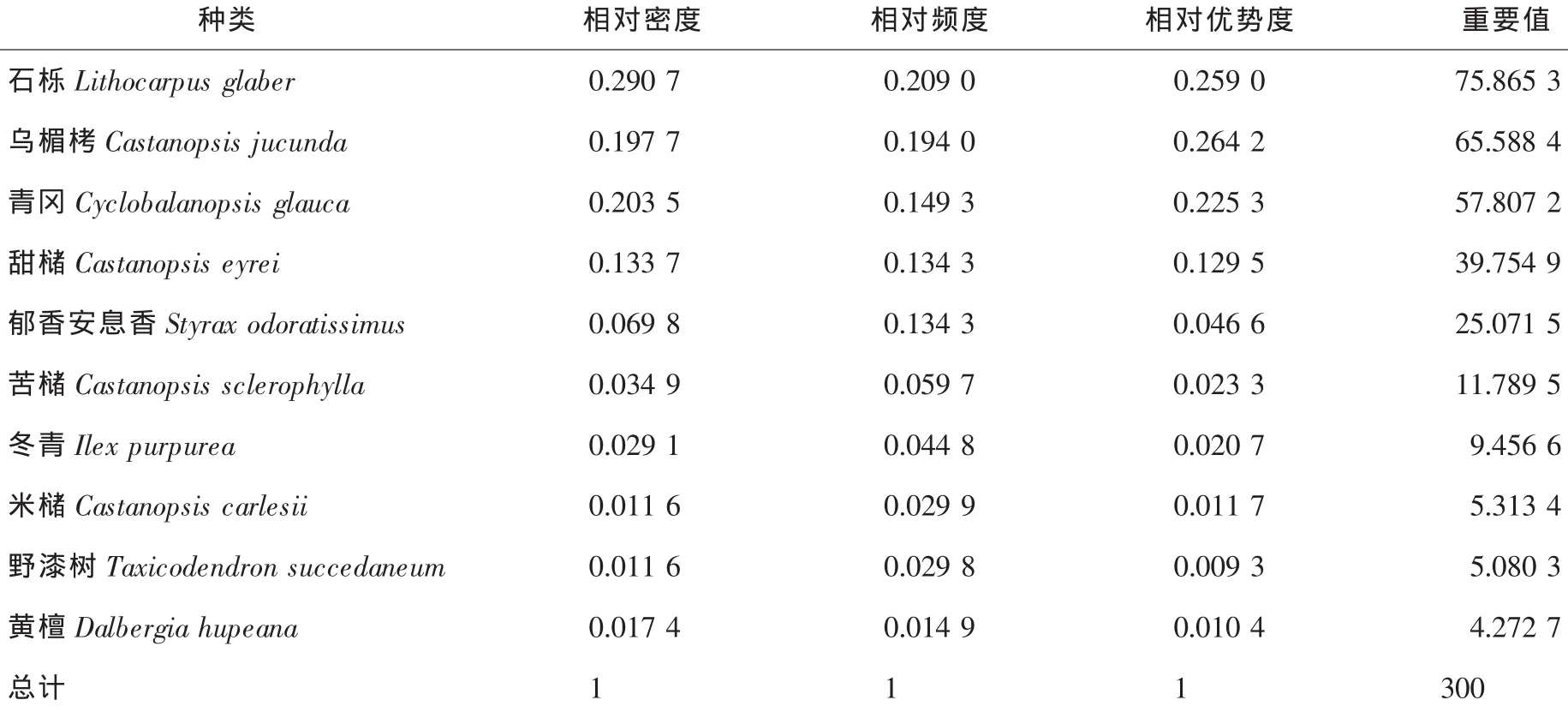
表4 甜槠-栲树林群落内主要乔木种群的重要值Table4 Importance values of tree populations in the Castanopsis eyrei-C.fargesii community
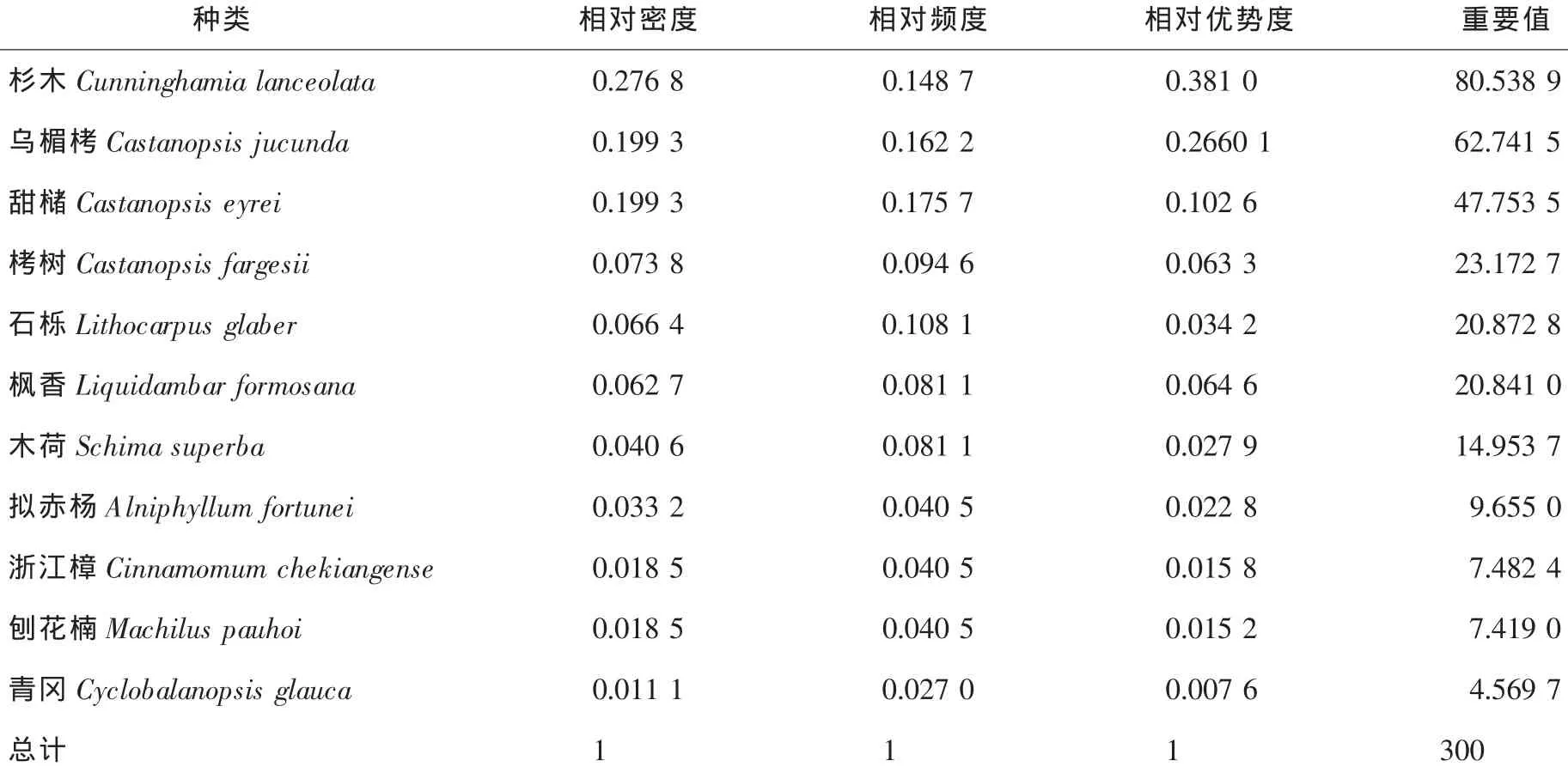
表5 杉阔混交林群落内主要乔木种群的重要值Table5 Importance values of tree populations in the Cunninghamia lanceolata and broadleaved mixed community
3.5 群落的物种多样性
物种多样性是指某一区域的物种数目和各物种分布的均匀程度,是衡量一定地区生物资源丰富程度的客观指标。它是根据一定空间范围物种的遗传多样性可以表现在多个层次上数量和分布特征来衡量的。由于环境因子对群落的物种组成有着深刻的影响,因此,这种影响必然要反映到群落物种多样性上来。由表7可见:总体来说,除杉阔混交林样地的Shannon-Weiner指数表现为草本层>灌木层>乔木层之外甜槠-栲树林、杉木人工林样地均表现出灌木层>乔木层>草本层。这是因为甜槠-栲树林和杉木人工林样地乔灌木层都很发达,郁闭度都较高,导致林下光照严重不足,因此草本层得不到充足的阳光而生长不良。而灌木层数量相对乔木层来说更为丰富,并且灌木层除了包括真正的灌木树种外,还包括了乔木层树种的幼苗、幼树等更新树种,如杉木、乌楣栲、栲树等。而杉阔混交林样地由于地处杉阔混交林,乔木层和灌木层都很稀疏,郁闭度很低,因此,没有森林凋落物影响草本层种子发育,良好的光照、水热条件和无凋落物层覆盖为草本层的生存与发展创造了最适宜的环境条件,使其种子有更多的萌发机会,同时充足的光照也大大促进了草本层的生长。
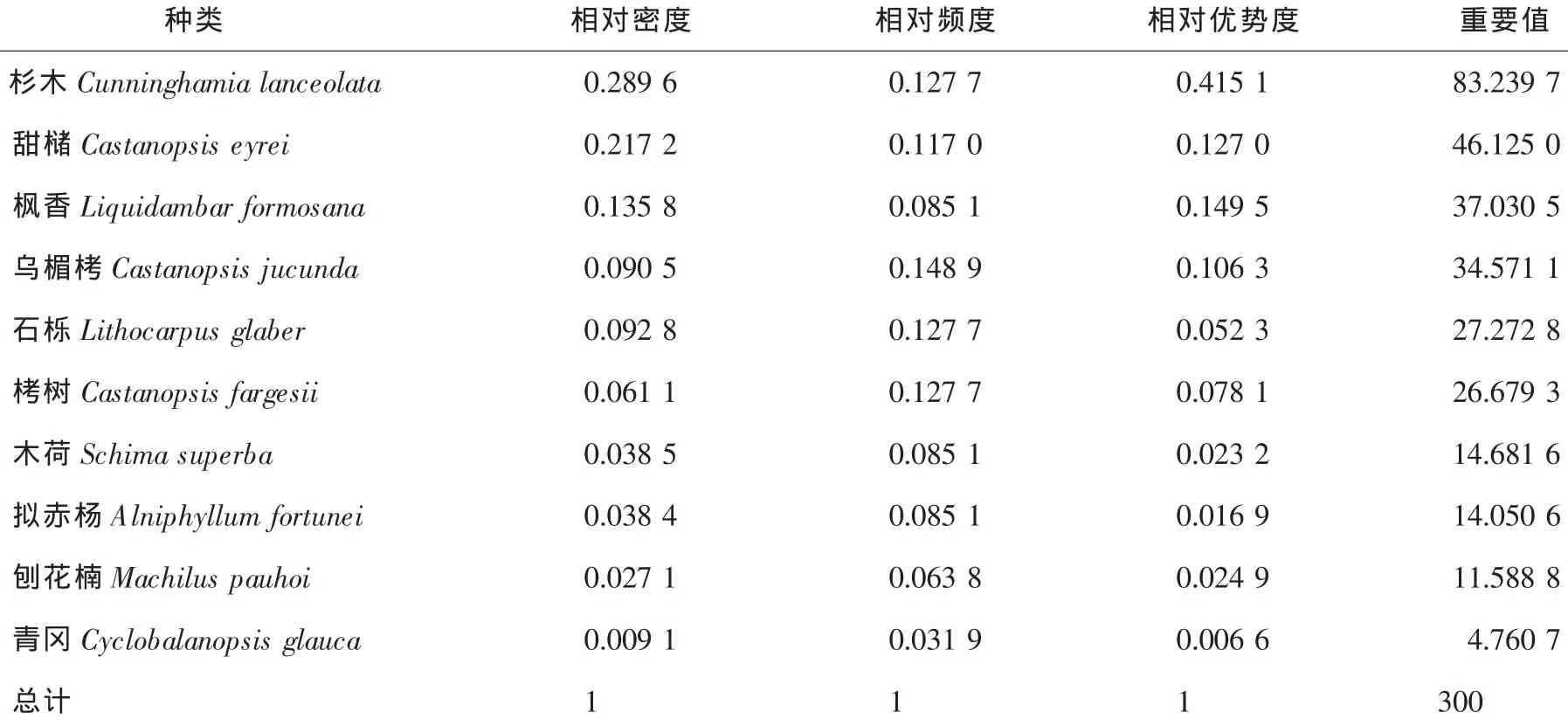
表6 杉木人工林群落内主要乔木种群的重要值Table6 Importance values of tree populations in the Cunninghamia lanceolata community

表7 群落乔木、灌木和草本层物种多样性Table7 Community species diversity indices of tree,shrub and herb layers
群落的优势度指数是衡量群落中优势种聚集程度的指标,其值越大则群落中一个或几个种的优势度就越高[19]。Simpson指数在甜槠-栲树林中表现为乔木层>灌木层>草本层。这是因为甜槠-栲树林样地中,石栎、甜槠、栲树、乌楣栲等乔木树种占的比例非常大,重要值很高,因此优势度十分明显;而在杉阔混交林样地中则表现为灌木层>草本层>乔木层,这是因为在此样地中堇叶紫金牛数量非常丰富,优势度很大,而草本层由于没有了乔灌木层的遮挡,有了充裕的阳光条件,生长也较为良好,尤其是荩草、淡竹叶等重要值也很高。在杉木人工林样地,Simpson指数表现为乔木层>草本层>灌木层,杉木人工栽培已有近30 a的历史,因此该样地中杉木占绝对优势地位,重要值也很大。由于杉木人工林郁闭度大,树冠浓密,冠幅大,林下的幼树、灌木生长受压制,导致其余树种生长势都不强,因此其灌木层的优势度也不高。Pielou均匀度指数也是变化度指数,物种数量越多,分布越均匀,其值就越大,变化趋势与Simpson指数基本一致。另外,乔木层盖度对灌木层物种多样性也有一定的影响,如杉木人工林样地,大量分布的杉木等乔木树种在对阳光、水分等竞争中处于优势地位,造成林下郁闭度较高,林下光环境恶化,物种数及其个体数量就会大幅度下降,因此灌木层物种丰富度较低。
3.6 主要乔木层组成种的径级结构
根据径级频率分布的形状,主要乔木层组成种可归纳为3个种群结构类型,即单峰型(unimodal type), 间歇性(sporadic type), 逆 J-字型(inverse-J type)[20]。 各类型代表树种和主要特征如图 2。
3.6.1 单峰型 只有少量大胸径个体出现,径级频率分布呈现1至数个不连续的单峰状径级,但缺乏小径级的个体更新,从种群的发展趋势看属于衰退类型。代表种有落叶阔叶乔木枫香,常绿针叶乔木马尾松和常绿阔叶乔木刨花楠。
3.6.2 间歇型 也称多峰型,群落内个体数较多,但由于亚乔木层中的中径个体受压枯死,使得径级频率分布呈不连续状,有多个单峰出现,同时具有较多的小径级后续更新个体,从种群的发展趋势看属于间歇型发展型。代表树种有常绿乔木树种青冈、木荷、乌药等。
3.6.3 逆J-字型 径级频率分布的形状类似倒逆放置的J字,个体的径级分布为连续分布,并呈现出2个峰,即大径级的小峰和小径级的大峰,小径级的后继个体较多。从种群的发展趋势来看为成熟稳定类型。代表种为常绿针叶乔木杉木,常绿阔叶乔木甜槠、栲树等。
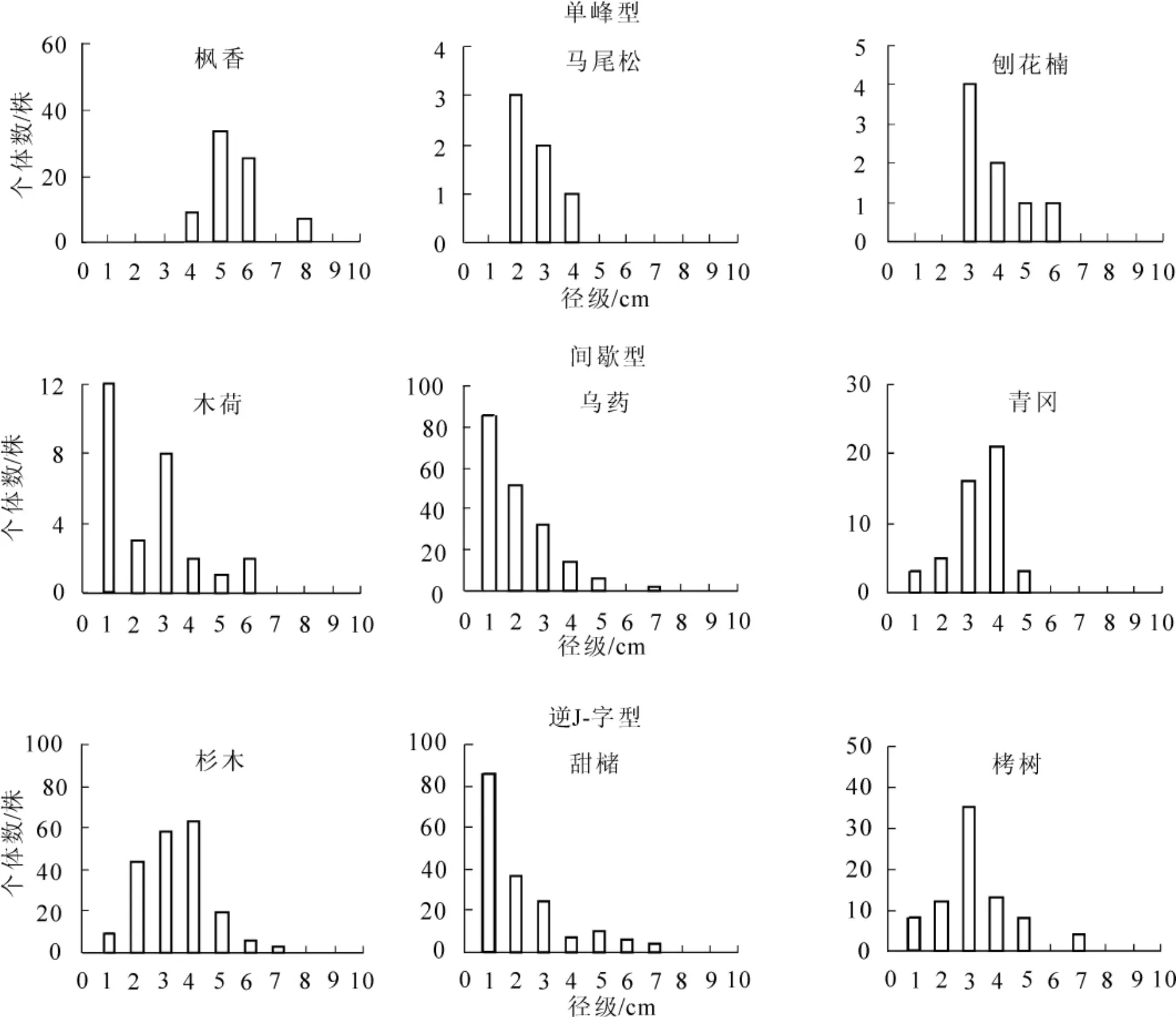
图2 堇叶紫金牛生存群落主要乔木层组成种的径级频率分布图Figure2 Diameter class frequency for tree layer species of survival community of Ardisia violacea
4 结论与讨论
堇叶紫金牛生存群落共有维管束植物116种(含变种),隶属于47科79属。根据其分布生境、群落结构和种类组成,可将堇叶紫金牛生存群落分为3个类型,即甜槠-栲树林、杉阔混交林、杉木人工林。堇叶紫金牛主要分布在光照、土壤、水分、养分等相对充足的林窗、林斑和岩石缝隙,尤其是在乔、灌木层均不发达,林下环境相对较为宽松的杉阔混交林中形成大量群聚。该类型分层不明显,高位芽植物稀少,但草本层非常丰富,占据绝对优势。
成熟稳定的群落是由一系列物种从原始生境一步步演化更替而来的[21]。单峰型的马尾松、枫香、刨花楠虽然分别为常绿针叶树、落叶树、常绿阔叶树,生活型有所不同,但却有相似的生态特性,都为阳性树种。王传华等[22]对枫香的研究认为:枫香幼苗的耐荫性会随着幼苗的个体发育发生变化,但目前尚没有发现幼苗耐荫性由 “需光型”剧烈地转变为 “耐荫型”的实例,并认为弱光环境是导致枫香幼苗更新不良的一大限制因子。程煜等[23]对马尾松的研究发现:马尾松在荫蔽状态下难以自我更新并有大量个体的枯死,必将是逐步被其他树种所替代。何贵平等[24]对刨花楠的研究中认为:刨花楠幼时喜荫、湿润环境,成年则喜光,在郁闭度高的群落中也会生长不良。
间歇型常绿阔叶树种青冈、木荷、乌药和檵木Loropefalum chinense,生态特性比较相似,都是具有中等程度的耐荫或者耐荫的特性,是主要的林冠构成种。姚贻强等[25]对青冈的研究认为,青冈在幼苗期由于灌木层的遮盖、郁闭度较大而生长受抑制,但当到了成熟期之后由于光照条件的增强,自然更新较好。逆J-字型的甜槠、栲树、乌楣栲、石栎、山矾等都为常绿阔叶乔木树种,群落中除了有大量的幼年个体外,还有一定的后续更新个体存在。胡正华等[26]和李志辉等[27]分别对古田山青冈林优势种群生态位特征和湘中丘陵区石栎群落研究中,也都证明了甜槠和石栎在亚热带常绿阔叶林中具有较大的生态位宽度,与林内其他树种的生态位重叠也很大。说明甜槠和石栎是亚热带常绿阔叶林顶极群落中的优势种和主要组成种。
物种丰富度指数与多样性指数是群落稳定性的重要指标,一定程度上决定于群落上层物种组成及其配比与空间配置 (配置方式、盖度等),而群落的稳定性与其密切相关,一般情况下,群落结构越复杂,群落越稳定,生态服务功能和生产力水平越高,抵御不良环境或外部干扰的能力愈强[28]。在3个样地中,灌木层中的物种丰富度指数均为最大。杉阔混交林中草本层的Shannon-Weiner指数最大,而甜槠-栲树林和杉木人工林中灌木层Shannon-Weiner指数最大。Simpson指数在甜槠-栲树林和杉木人工林中乔木层最大,而在杉阔混交林中灌木层最大。Pielou均匀度指数的变化趋势与Simpson指数基本一致。
通过分析各层土壤因子与群落结构特征关系,发现土壤因子与群落种群结构特征关系密切。各样地土壤相比较有如下特征:①由于甜槠-栲树林群落结构较复杂,石栎、甜槠、栲树等阔叶植物所形成凋落物分解后都会产生大量养分,因此,土壤有机质、全氮、全钾、全磷较高。②土壤含水量与群落郁闭度呈正比。群落郁闭度越大,土壤含水量越高,堇叶紫金牛数量越少;群落郁闭度越小,土壤含水量越低,堇叶紫金牛数量越多。
在调查中发现,堇叶紫金牛幼龄级数量比较丰富,生长较好。如果不加强人工抚育,幼苗幼树将很容易在演替过程中受到光照水分等环境因素的制约以及种内、种间竞争而数量减少。堇叶紫金牛种群的空间分布格局受群落特征影响主要体现在群落的组成和结构上。堇叶紫金牛作为共优种,其分布受制于其他共优种,如朱砂根、紫金牛,而常绿树种,如石栎、杉木、栲树等对它影响也比较大。例如在石栎、栲树等常绿阔叶树树冠下,堇叶紫金牛的数量极少,尤其是在杉木等常绿树种下,堇叶紫金牛的株数接近于0。群落内灌木层和藤本层的结构对堇叶紫金牛的分布影响较大,如在甜槠栲树林中,赤楠和络石的盖度在70%以上,地下根系盘结、错落有致,地上枝繁叶茂、隐蔽潮湿,阻碍种子的萌发和扩展,使种子的更新极其困难,与元宝山冷杉Abies yuanbaoshanensis很相似[29]。因此,在未来的保护中,首先应加强对原生境及植株的保护与监测,禁止采挖。应对堇叶紫金牛甜槠栲树林和杉木林进行适度的人为干预措施,注意间伐乔木层和灌木层过密树种、草本植物和非目标树种,适当扩大林窗、林斑面积,以降低林地郁闭度,扩大光照面积,加强群落通风条件,以减少与其他植物之间的生存竞争。同时应及时对其凋落物进行处理,以保证堇叶紫金牛种子顺利着土,促进种子的萌发。还要加强对堇叶紫金牛种群幼龄林的抚育管理,促进幼年个体的正常成长,加快幼苗更新和减少竞争压力,保持种群的稳定持续发展。另一方面应积极开展快繁技术研究,扩大资源量,并加强观赏应用研究,提高资源利用价值,以利野生资源的保护。
[1]达良俊,杨永川,宋永昌.浙江天童国家森林公园常绿阔叶林主要组成种的种群结构及更新类型[J].植物生态学报, 2004, 28 (3): 376-384.DA Liangjun, YANG Yongchuan, SONG Yongchang.Population structure and regeneration types of dominant species in an evergreen broadleaved forest in Tiantong National Forest Park, Zhejiang Province, eastern China [J].Acta Phytoecol Sin, 2004, 28 (3): 376-384.
[2]沈泽昊,金义兴,赵子恩,等.亚热带山地森林珍稀植物群落的结构与动态[J].生态学报,2000,20(5):800-807.SHEN Zehao, JIN Yixing, ZHAO Zien, et al.The structure and dynamics of the rare plant communities in subtropical mountian of China [J].Acta Ecol Sin, 2000, 20 (5): 800-807.
[3]刘振学,任广鑫,王得祥,等.通天河国家森林公园锐齿栎群落物种组成及群落结构研究[J].水土保持研究,2011, 18 (2): 86-90.LIU Zhenxue, REN Guangxin, WANG Dexiang, et al.Species compositon and structural characteristics of Quercus aliena community on National Forest Park of Tongtianhe River [J].Res Soil&Water Conserv, 2011, 18 (2): 86-90.
[4]SHIRO T, AKIRA H.Plant community dynamics on the volcand mount Koma, Northern Japan, after the 1996 eruption [J].Folia Geobot, 2005, 40: 319-330.
[5]ROBIN L M, ROGER F.Plant community, primary productivity, and environmental conditions following wetland reestablishment in the Sacramento-San Joaquin Delta, California [J].Wetlands Ecol Manage, 2010, 18: 1-6.
[6]杨沅志,张璐,陈北光,等.珍稀濒危植物广东松林的群落特征[J].华南农业大学学报,2006,27(2):70-73.YANG Yuanzhi, ZHANG Lu, CHEN Beiguang, et al.Characteristics of rare and endangered Pinus kwangtungensis forest[J].J South China Agric Univ, 2006, 27 (2): 70-73.
[7]红雨,邹林林,朱清芳.珍稀濒危植物蒙古扁桃群落结构特征[J].生态学杂志,2010,29(10):1907-1911.HONG Yu, ZOU Linlin, ZHU Qingfang.Community structure characteristics of endangered plant Prunus mongolica[J].Chin J Ecol, 2010, 29 (10): 1907-1911.
[8]苏文华,张光飞,欧阳志勤.稀有植物云南金钱槭生长群落特征与保护对策[J].云南植物研究,2006,28(1): 54-58.SU Wenhua, ZHANG Guangfei, OUYANG Zhiqin.Community characteristics and conservation strategies of a rare species, Dipteronia dyeriana [J].Acta Bot Yunnan, 2006, 28 (1): 54-58.
[9]张若蕙.浙江珍稀濒危植物[M].杭州:浙江科学技术出版社,1994:315-318.
[10]方云亿.浙江植物志:第5卷[M].杭州:浙江科学技术出版社,1989:31.
[11]BAKKER J P, OLEF H J, WILLEMS J H, et al.Why do we need permanent plots in the study of long-term vegetation dynamics? [J].J Veg Sei, 1996, 7: 147-156.
[12]朱圣潮.浙江丽水太山褐叶青冈林群落结构及其多样性[J].中南林学院学报,2006,26(4):56-60.ZHU Shengchao.Community structure and species diversity of form Cyclobalanopsis stewardiana of Mt.Taishan in Lishui, Zhejiang [J].J Central South For Univ, 2006, 26 (4): 56-60.
[13]张志祥,刘鹏,刘春生,等.浙江九龙山南方铁杉群落结构及优势种群更新类型[J].生态学报,2008,28(9):4547-4558.ZHANG Zhixiang, LIU Peng, LIU Chunsheng, et al.The structure characteristics and dominant population regeneration types of Tsuga tchekiangensis communities in the Jiulongshan National Natural Reserve of Zhejiang Province[J].Acta Ecol Sin, 2008, 28 (9): 4547-4558.
[14]李先琨,苏宗明,向悟生,等.濒危植物元宝山冷杉种群结构与分布格局[J].生态学报,2002,22(12):2245-2253.LI Xiankun, SU Zongming, XIANG Wusheng, et al.Study on the structure and spatial pattern of the endangered plant population of Abies yuanbaoshanensis [J].Acta Ecol Sin, 2002, 22 (12): 2245-2253.
[15]申仕康,马海英,王跃华,等.濒危植物猪血木自然种群结构及动态[J].生态学报,2008,28(5):2404-2412.SHEN Shikang, MA Haiying, WANG Yuehua, et al.The structure and dynamics of natural population of the endangered plant Euryodendron excelsum H.T.Chang [J].Acta Ecol Sin, 2008, 28 (5): 2404-2412.
[16]胡正华,于明坚.古田山青冈林优势种群生态位特征[J].生态学杂志,2005,24(10):1159-1162.HU Zhenghua,YU Mingjian.Niche characteristics of dominant populations in Cyclobalanopsis glauca forest in Gutian Mountain [J].Chin J Ecol, 2005, 24 (10): 1159-1162.
[17]兰国玉,陈伟,周小飞.海南霸王岭青梅林群落特征研究[J].西北植物学报,2007,27(9):1861-1868.LAN Guoyu, CHEN Wei, ZHOU Xiaofei.Community characteristics of Vatica mangachapoi forest of Bawangling in Hainan, South China [J].Acta Bot Boreali-Occident Sin, 2007, 27 (9): 1861-1868.
[18]魏琦,楼炉焕,冷建红,等.毛枝连蕊茶群落结构与物种多样性[J].浙江农林大学学报,2011,28(4):634-639.WEI Qi, LOU Luhuan, LENG Jianhong, et al.Community structure and species diversity of Camellia trichoclada[J].J Zhejiang A&F Univ, 2011, 28 (4): 634-639.
[19]杨旭.凤阳山自然保护区白豆杉种群生态学的研究[D].杭州:浙江大学,2005.YANG Xu.Study on the Population Ecological of Pseudotaxus chienii in Fengyangshan National Natural Reserve[D].Hangzhou: Zhejiang University, 2005.
[20]杨同辉,王良衍.浙江天童舟山新木姜子群落结构及其更新类型[J].浙江林学院学报,2010,27(6):877-883.YANG Tonghui, WANG Liangyan.Community structure and regeneration types of Neolitsea sericea in Tiantong,Zhejiang Province [J].J Zhejiang For Coll, 2010, 27 (6): 877-883.
[21]BOSSUYT B, HONNAY O, HERMY M.Evidence for community assembly constraints during succession in dune slack plant communities [J].Plant Ecol, 2005, 178: 201-209.
[22]王传华,李俊清,陈芳清,等.鄂东南低丘地区枫香林下枫香幼苗更新限制因子[J].植物生态学报,2011,35(2): 187-194.WANG Chuanhua, LI Junqing, CHEN Fangqing, et al.Factors affecting seedling regeneration of Liquidambar formosana in the L.formosana forests in hilly regions of Southeast Hubei, China [J].Chin J Plant Ecol, 2011, 35 (2):187-194.
[23]程煜,胡喜生,洪伟,等.木荷马尾松林更新层种群生态位分析[J].福建林学院学报,2009,29(3):220-225.CHENG Yu, HU Xisheng, HONG Wei, et al.Study on species niche of Schima superba and Pinus massoniana artificial forest shrub layer [J].J Fujian Coll For, 2009, 29 (3): 220-225.
[24]何贵平,骆文坚,冯建民,等.不同立地条件刨花楠天然群落物种多样性比较研究[J].浙江林业科技,2003,23(5): 1-3.HE Guiping, LUO Wenjian, FENG Jianmin, et al.Comparison study on diversity of natural Machilus pauhoi community under different site conditions [J].J Zhejiang For Sci Technol, 2003, 23 (5): 1-3.
[25]姚贻强,张忠华,梁士楚,等.桂林岩溶石山青冈栎种群的结构特征[J].浙江林业科技,2008,28(4):8-11.YAO Yiqiang, ZHANG Zhonghua, LIANG Shichu, et al.Structure of Cyclobalanopsis glauca population on karst hills of Guilin [J].J Zhejiang For Sci Technol, 2008, 28 (4): 8-11.
[26]胡正华,于明坚,徐学红,等.浙江古田山自然保护区甜槠群落特征研究[J].生态学杂志,2004,23(2):15-18.HU Zhenghua, YU Mingjian, XU Xuehong, et al.Castanopsis eyrei community characteristics in Gutianshan Natural Reserve, Zhejiang Province [J].Chin J Ecol, 2004, 23 (2): 15-18.
[27]李志辉,朱日光.湘中丘陵区石栎群落研究[J].广西植物,2000,20(1):11-17.LI Zhihui, ZHU Riguang.Studies on Lithocarpus glaber community Central Hunan’s hilly country [J].Guihaia, 2000, 20 (1): 11-17.
[28]康永祥,康博文,刘建军,等.陕北黄土高原文冠果群落结构及物种多样性[J].生态学报,2010,30(16):4328-4339.KANG Yongxiang, KANG Bowen, LIU Jianjun, et al.Structure and species diversity of Xanthoceras sorbifolia community in the Loess Plateau of North Shaanxi[J].Acta Ecol Sin, 2010, 30 (16): 4328-4339.
[29]李先琨,苏宗明,向悟生,等.濒危植物元宝山冷杉种群结构与分布格局[J].生态学报,2002,22(12):2245-2253.LI Xiankun, SU Zongming, XANG Wusheng, et al.Study on the structure and spatial pattern of the endangered plant population of Abies yuanbaoshanensis [J].Acta Ecol Sin, 2002, 22 (12): 2245-2253.
猜你喜欢
广西林业科学(2022年4期)2022-11-07
林业调查规划(2022年3期)2022-06-13
园艺与种苗(2021年3期)2021-05-26
农村实用技术(2020年1期)2020-12-16
农民致富之友(2020年8期)2020-05-11
现代园艺(2018年20期)2018-01-19
现代园艺(2018年14期)2018-01-18
中央民族大学学报(自然科学版)(2017年4期)2017-06-11
绿色科技(2016年15期)2016-10-11
安徽农业科学(2015年9期)2015-01-12
