
小佛肚竹生氰糖苷合成关键酶CYP79家族同源基因的克隆和鉴定
2012-05-30 03:29王月圆刘向敏周明兵汤定钦
浙江农林大学学报 2012年4期
王月圆,刘向敏,周明兵,汤定钦
(浙江农林大学 亚热带森林培育国家重点实验室培育基地,浙江 临安 311300)
生氰糖苷(cyanogenic glycosides)亦称氰苷、氰醇苷,是由氰醇衍生物的羟基和D-葡萄糖缩合形成的糖苷,已在2650多种植物中发现,分属于蕨类植物(蕨类)、裸子植物和被子植物(开花植物)的130个家族[1-2]。生氰糖苷的主要生物学功能在于阻止食草动物和病原体的对植物侵害。当生氰植物(本身含有生氰糖苷的植物)外部受损时,生氰糖苷将会产生有毒的氰化氢(HCN),从而阻止食草动物和病原体的进一步伤害[3]。生氰糖苷的生物合成过程比较简单,一般包括3个步骤,催化生氰糖苷合成的酶主要是2种细胞色素P450,属于CYP79家族和CYP71家族,另外还有一种葡萄糖转移酶UGT85B1[4]。本研究首次在竹子中发现和鉴定到生氰糖苷合成的关键酶CYP79家族同源基因(DQ409175),表明在竹亚科Bambusoideae也可能存在生氰糖苷途径,为研究竹类植物生氰糖苷的生物合成,人为控制生氰糖苷生物合成,从而干扰其毒性的产生,保证食笋的安全性奠定了基础。
1 材料与方法
1.1 实验材料
TaqDNA聚合酶,三磷酸碱碱基脱氧核苷酸(dNTPs),Agarose Gel DNA Extraction Kit,PCR Product Extraction Kit等均为上海生工公司产品,大肠埃希菌Escherichia coli转化的受体为大肠埃希菌DH5α菌株,克隆质粒载体为pMD18-T。在浙江省临安市太湖源观赏竹种园采集小佛肚竹Bambusa ventricosa幼嫩的笋作为基因克隆的起始材料。
1.2 总RNA提取
采用Invitrogen公司的Trizol reagent提取小佛肚竹幼笋的核糖核酸(RNA),用Invitrogen公司的反转录试剂盒Su-perScriptTMFirst Strand Synthesis System for RT-PCR合成cDNA。
1.3 基因的克隆与序列分析
根据已知CYP79家族同源基因序列设计上游引物PF(5′-CCG TGG AAG GAG ACG CTG TCG-3′)和下游引物PR (5′-GGG GTC GCA GTG CGG ACC GGT GCC-3′),由上海生工生物工程有限公司合成。以小佛肚竹cDNA为模板,片段扩增温度反应条件为:94℃ 3 min;94℃ 45 s,55℃ 45 s,72℃ 1 min 40 s,35个循环;72℃延伸10 min。聚合酶链式反应(PCR)产物克隆到pMD18-T载体,转化大肠埃希菌DH5α菌株,经蓝白斑筛选,提取阳性克隆质粒并酶切图谱分析后,再将单克隆送上海生工生物工程有限公司测序。
基因序列的3′端延伸和5′端延伸分别采用Invitrogen公司的3’RACE kit和TaKaRa公司的BD SMARTTMRACE cDNA Amplification Kit。 3′-RACE 的 2 条特异引物分别为 P3-1:5′-GGC GAC CAG TGG AAG AAG ATG-3′和 P3-2: GCG GCT ACA AGA ACG CAG TT, 精简的通用扩增引物(AUAP): 5′-GGC CAC GCG TCG ACT AGT AC-3′。 5′-RACE 的特异引物分别为 P5-1: 5′-TTC ACC ATA GCC TCC TAC AGT CCA ACC AAA-3′, 通用套式引物(NUP):AAG CAG TGG TAT CAA CGC AGA GT。
3′末端扩增反应条件同中间序列扩增一致,5′末端扩增反应条件为:94℃30 s,72℃ 3 min,5个循环;70℃30 s,72℃ 3 min,5个循环;94℃30 s,68℃ 30 s,72℃3 min,27个循环。聚合酶链式反应(PCR)产物克隆到pMD18-T载体,送上海生工生物工程有限公司测序。根据中间扩增片段、3′末端扩增片段和5′末端扩增片段的序列利用Vector NTI Advance 9软件拼接成全长序列。设计一对引物(CICR-5: CTGAAGGACTAGGAGACATCAGC和CICR3:GGGCACCATAGCC-TCCTACAG)一次性将全长序列扩增出来,克隆到pMD18-T载体后,测序验证。利用DNASTAR和Clustl W等生物软件分析测定cDNA序列及其编码蛋白质的结构特点,并与美国国家生物技术信息中心(NCBI)蛋白质数据库进行Blast比较分析。
1.4 系统树的构建
采用邻位相连法(neighbor-joining,N-J法)绘制系统树,并对其进行自展(bootstrapvalue 1000)性评估;最后用TreeView软件观看、输出图形化的系统树,每个类群出现的概率用百分数表示。
2 结果与分析
2.1 小佛肚竹BvCYP79基因的克隆
以小佛肚竹cDNA为模板,上游引物(PF)和下游引物(PR)扩增出 400 bp左右有1条片段(图1A),与预测的基因片段长度基本相符,初步确定为目的基因片段。
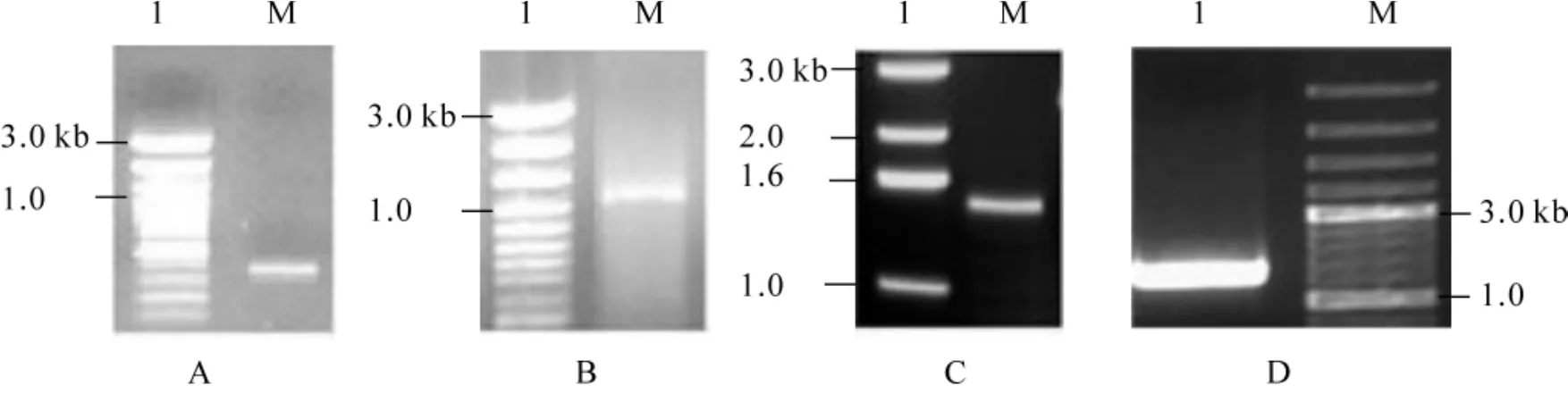
图1 聚合酶链式反应(PCR)产物电泳检测Figure1 Gel electrophoresis of amplification fragments of BvCYP79
测序结果表明:插入片段为379 bp。应用NCBI Blast在线软件,将测序的序列与已报道的基因进行同源比较,发现编码的氨基酸编码序列含有CYP79蛋白家族的保守域,初步确认为扩增的部分基因序列来源于CYP79家族基因。
再次以小佛肚竹cDNA为模板,用特异引物P3-1,P3-2和精简的通用扩增引物(AUAP)扩增出1411 bp左右的条带,特异引物P5-1和通用套式引物(NUP)扩增出1102 bp左右的条带(图1B,C),经测序后,与中间片段拼接得小佛肚竹CYP79家族基因的的全长cDNA,根据两端序列设计相应的引物,进行全长序列扩增(图1D),测序结果与拼接结果一致,遂命名为BvCYP79。该全长cDNA序列为1913 bp(图2),翻译起始点为193碱基处,终止子位于1752 bp,包含1个1559 bp的完整编码区,开放阅读框共编码519个氨基酸(图2)。
2.2 小佛肚竹BvCYP79氨基酸序列分析
应用ProtParam(http://us.expasy.org/tools/protparam.html)对上述推测的氨基酸序列进行分析。BvCYP79编码蛋白质的分子量(MW)58149.8D, 理论等电点(theoreticalPI)为 6.00, 分子式(formula)为C2594H4101N741O738S21。用PBIL Network Protein Sequence Analysis对基因编码蛋白质产物的二级结构进行了分析,BvCYP79的二级结构中41.43%为螺旋结构(helices),5.78%为线状结构(strands),52.79%为卷曲结构(coils),卷曲结构和螺旋结构是BvCYP79蛋白质二级结构的骨架。
应用Blast进行氨基酸序列相似性比较,BvCYP79与大麦Hordeum vulgare的HvCYP79A8(ACJ70085),琴叶拟南芥 Arabidopsis lyrata的 AlCYP79B2(XP-002866896)以及拟南芥 A.thaliana的AtCYP79B2(NP-195705)的相似性最高,均达99%,其次为甘蓝型油菜Brassica napus的BnCYP79B5(AF453287-1),相似性为98%,和甘蓝B.oleracea的BoCyp79B1(ADW54459)相似性为97%,和杨毛果Populus trichocarpa的PtCYP79D8(XP-002305081)相似性为96%。上述蛋白质序列都聚具有CYP79家族特有的PERH区域和与处在血红素结合区的保守序列(S/T)F(S/T)TGRRGCXG。
2.3 小佛肚竹BvCYP79聚类分析
植物细胞色素P450家族(the cytochrome P450 superfamily)又称之为CYP家族,其基因总数尚不清楚,至今已鉴定90多个植物P450基因,分属于27个基因(CYP51,CYP71,CYP72,…,CYP96)[5]。选取34个与BvCYP79同源关系较近的P450基因的氨基酸与BvCYP79氨基酸序列做聚类分析,结果表明:BvCYP79与高粱Sorghum bicolor的SbCYP79A1进化关系最近,喻示着它们之间有类似的生物学功能(图3)。高粱的SbCYP79A1是在生氰糖苷合成中首先被分离和鉴定的酶,是从高粱黄化苗的微粒体中分离出来[6]。在高粱黍蜀氰苷代谢过程中,CYP79A1,CYP71E1和UGT85B1作为一个联合体共同起作用,在NADPH(细胞色素P450还原酶)的参与下,由氰醇衍生物的羟基和D-葡萄糖催化缩合形成的糖苷。BvCYP79基因在小佛肚竹的克隆和鉴定表明生氰糖苷途径也可能存在于竹类植物。

图2 小佛肚竹BvCYP79碱基序列和推测的氨基酸序列Figure2 Sequence of BvCYP79

图3 小佛肚竹BvCYP79与34个植物P450基因的氨基酸序列采用邻位相连法构建系统树Figure3 Phylogenetic relationships of the translated BvCYP79 and 34 plant cytochrome P450 protein sequences collected in databanks
3 讨论
生氰糖苷在2000多种植物中发现,分布于蕨类植物、裸子植物和被子植物。生氰的蕨类植物和裸子植物含有芳香族生氰糖苷,源自酪氨酸或苯丙氨酸,而被子植物含有脂肪族以及芳香族生氰糖苷分别来自于异亮氨酸、缬氨酸、亮氨酸或酪氨酸、苯丙氨酸。另外,一些植物如西番莲Passiflora caerulea含有生氰糖苷来自蛋白氨基酸环戊烯甘氨酸。当含有生氰糖苷的植物组织受损害时,生氰糖苷将被β-糖苷酶和α-羟氰酶降解,释放出有毒的氰化氢以及葡萄糖和醛或酮,使植物对草食动物和病原体造成的组织损伤进行即时的化学防御反应[7-8];转基因植物也可以容纳其他植物的生氰糖苷合成途径,积累产生生氰糖苷,达到阻止特殊昆虫危害的目的。如在转基因拟南芥异源表达3个高粱基因的研究中,发现专食十字花科Cruciferae植物的跳蚤甲虫不吃含有黍蜀氰苷的拟南芥。这清楚地显示代谢工程在生产所需特性的农作物方面的潜力[9]。小佛肚竹是优良的观赏竹类,在园林观赏上具有广泛的应用前景,调查BvCYP79功能,为增强小佛肚竹的病害防御功能及通过基因工程手段调控小佛肚竹生氰糖苷生物合成奠定了基础。
生氰糖苷在分子水平上的进化信息,目前在植物中只限于CYP79家族。从植物中选定71个CYP79家族成员进行系统发育重建工作,表明CYP79家族在被子植物和裸子植物分化前(3亿年前)起源于共同祖先,反映了生氰糖苷的古老性[10]。本研究利用小佛肚竹BvCYP79和34个植物P450基因的氨基酸的聚类分析结果也表明P450基因应该起源的共同祖先,并在单子叶植物和双子植物分化之前就开始了功能的歧化。
与其他家族的植物色素P450相比,CYP79家族的特点是一般具有保守的“PERF”和血红素结合域[11]。在单子叶植物CYP79家族的PERF区域,保守的非极性的和芳香族的苯丙氨酸残基被带正电荷的氨基酸残基组氨酸所取代而使单子叶植物CYP79家族保持着PERH区域。同样,在血红素结合区保守序列PFGXGRRXCXG,疏水性氨基酸用极性的丝氨酸或苏氨酸残基进行3种替换,提供了单子叶植物CYP79家族相应保守序列为(S/T)F(S/T)TGRRGCIA[6,12], 小佛肚竹 BvCYP71 的编码氨基酸在上述 2 个保守域分别为PERF和SFSTGRRGCIA,属于典型的单子叶植物CYP79家族基因(图3)。本课题计划通过原核和真核表达分析BvCYP71催化功能,探讨生氰糖苷对小佛肚竹生长发育的影响。
[1]CONN E E.Cyanogenic glycosides [J].Biochem Plants, 1981, 7: 479-500.
[2]SIEGLER D S, BRINKER A M.Characterisation of cyanogenic glycosides, cyanolipids, nitroglycosides, organic nitro compounds and nitrile glycosides from plants [G]//DEY P M, HARBORNE J B.Methods of Plant Biochemistry,Alkaloids and Sulfur Compounds.New York: Academic Press, 1993: 51-93.
[3]WINKEL B S J.Metabolic channeling in plants [J].Annu Rev Plant Biol, 2004, 55: 85-107.
[4]HANSEN K S, KRISTENSEN C, TATTERSALL D B, et al.The in vitro substrate regiospecificity of recombinant UGT85B1, the cyano-hydrin glucosyltransferase from Sorghum bicolor[J].Phytochemistry, 2003, 64: 143-151.
[5]MORANT M, BAK S, MØLLER B L, et al.Plant cytochromes P450: tools for pharmacology, plant protection and phytoremediation [J].Curr Opin Biotechnol, 2003, 14: 151-162.
[6]BAK S, KAHN R A, NIELSEN H L, et al.Cloning of three A-type cytochromes P450, CYP71E1, CYP98, and CYP99 from Sorghum bicolor (L.) Moench by a PCR approach and identification by expression in Escherichia coli of CYP71E1 as a multifunctional cytochrome P450 in the biosynthesis of the cyanogenic glucoside dhurrin [J].Plant Mol Biol,1998, 36: 393-405.
[7]JONES P R, ANDERSEN M D, NIELSEN J S, et al.The biosynthesis, degradation, transport and possible function of cyanogenic glucosides [G]//ROMEO J T, IBRAHIM R, VARIN L, et al.Evolution of Metabolic Pathways.New York: Elsevier Science, 2000: 191-247.
[8]TATTERSALL D B, BAK S, JONES P R, et al.Resistance to an herbivore through engineered cyanogenic glucoside synthesis [J].Science, 2001, 293: 1826-1828.
[9]MORANT A V, JØRGENSEN K, JØRGENSEN B, et al.Lessons learned from metabolic engineering of cyanogenic glucosides [J].Metabolomics, 2007, 3: 383-398.
[10]BAK S, PAQUETTE S, MORANT M, et al.Cyanogenic glycosides: a case study for evolution and application of cytochromes P450 [J].Phytochem Rev, 2006, 5: 309-329.
[11]WERCK-REICHHART D, BAK S, PAQUETTE S.Cytochrome P450 [R]//SOMERVILLE C R, MEYEROWITZ E M.The Arabidopsis Book.Rockville: American Society of Plant Biologists, 2002.
[12]ANDERSEN M D, BUSK P K, SVENDSEN I, et al.Cytochromes P-450 from cassava (Manihot esculenta Crantz)catalyzing the first steps in the biosynthesis of the cyanogenic glucosides linamarin and lotaustralin: cloning, functional expression in Pichia pastoris, and substrate specificity of the isolated recombinant enzymes [J].J Biol Chem, 2000,275:1966-1975.
猜你喜欢
中国中医药现代远程教育(2022年17期)2022-09-26
环球时报(2022-09-20)2022-09-20
中国马铃薯(2022年2期)2022-07-05
中草药(2022年11期)2022-05-31
中草药(2022年9期)2022-05-06
今日农业(2020年24期)2020-12-15
中国洗涤用品工业(2019年4期)2019-05-11
中成药(2017年3期)2017-05-17
小资CHIC!ELEGANCE(2015年14期)2015-09-23
小资CHIC!ELEGANCE(2015年15期)2015-09-01
