
小菜蛾鱼尼丁受体基因克隆及序列分析
2012-06-12 00:58孙丽娜芮昌辉袁会珠
植物保护 2012年4期
孙丽娜, 芮昌辉, 袁会珠
(中国农业科学院植物保护研究所,农业部作物有害生物综合治理综合性重点实验室,北京 100193)
鱼尼丁受体(RyR)是目前所知最大的钙离子通道,由4个相同的亚基组成,每个亚基的分子量约为560ku。在脊椎动物中存在3种亚型RyRs,而在昆虫中仅存在一种鱼尼丁受体[1]。二酰胺类化合物氟虫双酰胺和氯虫苯甲酰胺对鳞翅目害虫具有较高的活性,其作用机制与其他类杀虫剂不同,它们可以结合昆虫体内的鱼尼丁受体,抑制昆虫取食,引起虫体收缩,使虫体变粗变短,最终导致死亡[2-3]。自两个化合物创制成功以来,不仅作用于鱼尼丁受体化合物的合成备受关注,昆虫鱼尼丁受体的基因克隆及分子特性也因此成为焦点[4-5]。
小菜蛾[Plutella xylostella(L.)]是一种为害十字花科植物的世界性害虫,也是对农药产生抗性的最严重害虫之一。据不完全统计,小菜蛾已经对70多种杀虫剂产生了抗药性,包括有机氯、有机磷、氨基甲酸酯、拟除虫菊酯类,以及昆虫生长调节剂和苏云金杆菌(Bt)等杀虫剂[6]。尽管二酰胺类化合物对小菜蛾具有较高的活性,但是有研究称小菜蛾对其抗性产生较快。田间监测表明,在我国华南地区田间小菜蛾对氯虫苯甲酰胺的抗性与室内敏感小菜蛾相比已达到100多倍[7];而在泰国,2008年至2011年间,小菜蛾对氟虫酰胺的抗性从1.5倍增至4 817倍,对氯虫苯甲酰胺的抗性从35.4倍增至152倍[8]。因此克隆小菜蛾鱼尼丁受体基因可以为今后小菜蛾抗性机制研究提供一定的理论基础。
1 材料与方法
1.1 供试昆虫
小菜蛾于室内采用无药剂接触甘蓝苗连续饲养的技术饲养[9]。
1.2 总RNA的提取和cDNA第一链的合成
取3龄小菜蛾幼虫,用液氮研磨后,参照Trizol(Invitrogen)试剂说明书步骤提取总RNA。以反转录酶(PrimeScript®1st Strand cDNA Synthesis Kit,TaKaRa,大连,中国)合成cDNA第一链;利用TaKaRa 3′和5′RACE 试剂盒合成3′和5′RACE cDNA第一链。
1.3 引物设计
如图1所示,小菜蛾RyR序列可以由18个片段重叠组合得到。本试验引物设计根据美国国家生物工程信息中心(NCBI)数据库中家蚕B.m.(Bombyx mori)、烟芽夜蛾 H.v.(Heliothis virescens)、黑腹果蝇D.m.(Drosophila melanogaster)、桃蚜 M.p.(Myzus persicae)、棉蚜A.g.(Aphis gossypii)、玉米飞虱P.m.(Peregrinus maidis)、埃及伊蚊A.a(Aedes aegypti)这个7个物种鱼尼丁受体基因氨基酸中保守序列来设计简并引物。根据简并引物的PCR测序结果,设计特异性引物,引物见表1。所有引物的合成由北京六合华大基因科技有限公司完成。
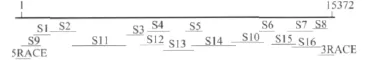
图1 PCR扩增和克隆小菜蛾鱼尼丁受体(Px-RyR)cDNA片段示意图

表1 克隆小菜蛾鱼尼丁受体基因的引物1)
1.4 PCR扩增
片段S1~S19的扩增,按照TaKaRa Ex Taq试剂盒说明进行,按照不同的引物及扩增片段的长度设置不同的PCR反应程序。
cDNA 3′末端快速扩增(3′RACE),outer PCR和inner PCR的反应条件均为:94℃3min;94℃30s,65℃→50℃(-0.5℃/循环)30s,72℃1min,30个循环;94℃30s,50℃30s,72℃1min,15个循环,72℃10min。
cDNA 5′末端快速扩增(5′RACE),outer PCR反应条件为:94℃3min;94℃30s,55℃30s,72℃1min,25个循环;72℃10min。Inner PCR反应条件为:94℃3min;94℃30s,68℃1min,30个循环;72℃10min。
将目的片段克隆于pMD®19-T Simple Vector载体内,选取阳性克隆测序。序列测定由北京六合华大基因科技股份有限公司完成。
1.5 序列测定及分析
利用DNAstar和Mega5.0等软件进行序列分析和比较。利用美国国家生物工程信息中心(NCBI,http:∥www.ncbi.nlm.nih.gov/BLAST/)的BLAST网络工具与GenBank中的其他物种序列进行比较,确定目标产物的真实可靠性。分子量和等电点(molecular weight/isoelectric point)MW/pI在ExPaSy website(http:∥us.expasy.org/tools/pitool.htm)预测计算。跨膜区域预测可以于www.cbs.dtu.dk/services/TMHMM 进行分析,蛋白质二级 结 构 于 http:∥www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi和http:∥www.expasy.org/tools/scanprosite/分析。
2 结果与分析
根据NCBI中昆虫鱼尼丁受体基因序列设计简并引物,通过RT-PCR扩增得到了目标片段,而后根据目的片段序列,再设计特异性引物,继续进行RT-PCR扩增。进一步进行5′RACE和3′RACE扩增,得到两端序列后,通过小菜蛾所有片段的序列拼接得到小菜蛾鱼尼丁受体基因cDNA序列全长。小菜蛾鱼尼丁受体基因全长为15 748bp,其中5′端非阅读区267bp,3′端非阅读区109bp,开放阅读区15 372bp。开放阅读区编码5 123个氨基酸残基(GenBank登录号为JF927788),预测其分子量约为579.39ku,等电点为5.45。
对预测的Px-RyR氨基酸序列与其他物种RyR基因氨基酸序列进行多重比对,结果表明Px-RyR与其他物种RyR存在很高的同源性如表2。与烟芽夜蛾(H.v.)、家蚕(B.m.)、桃蚜(M.p.)、棉蚜(A.g.)、玉米飞虱(P.m.)、果蝇(D.m.)、鼠(M.m.)骨骼肌RyR1、鼠心肌RyR2及鼠RyR3的相似性分别为92%、92%,77%、77%、80%、78%、45%、47%、46%。

表2 Px-RyR与其他物种RyRs序列相似性 %
通过MEGA 5.0利用邻接法对22个物种RyR蛋白质进行进化树分析(Bootsrap为1 000个重复),从而通过RyR氨基酸多态性来揭示这些物种间RyR蛋白质的进化关系。由图2看出,22个物种间的RyR基本分两大类,即脊椎动物中一类,无脊椎动物中一类。图2可以明显看出,小菜蛾RyR与烟芽夜蛾和家蚕RyR关系最近。
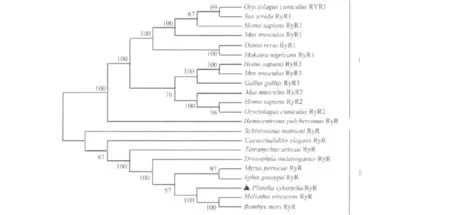
图2 Px-RyR与其他21个物种间RyR蛋白质进化树分析
根据氨基酸二级结构预测小菜蛾鱼尼丁受体基因存在6个跨膜区域,位于第4 474~5 022氨基酸之间。这6个氨基酸序列分别为(如表3):Phe4474-Leu4496;Asp4655-Tyr4676;Val4734-Leu4756;Phe4875-Ala4897;Leu4923-Phe4945;Ile5003-Ile5022。除了跨膜区域外,小菜蛾鱼尼丁受体蛋白还存在其他结构域。如RyR结构域(重复序列)、细胞内钙离子释放通道结构域、钙离子结合的EF-hand结构域、MIR结构域(因该结构域同时存在于甘露糖转移酶、三磷酸肌醇受体和鱼尼丁受体中而命名)、Spla/RYanodine receptor SPRY结构域、离子通道结构域、鱼尼丁受体和三磷酸肌醇受体相似结构域、似SPRY结构域及一些未命名的结构域[10]。

表3 小菜蛾RyR跨膜区域序列
3 结论与讨论
鱼尼丁受体是最大的钙离子释放通道之一,在肌肉兴奋收缩偶联中具有重要作用,在很多细胞中具有第二信使的作用。鱼尼丁受体因其所具有的特性,近30年来在医学界备受关注,而近10年来在杀虫剂研究领域也深受广大研究者关注[1]。因此克隆昆虫鱼尼丁受体基因对作用于鱼尼丁受体新化合物的设计及害虫抗药性研究至关重要。在本研究中,笔者克隆了小菜蛾鱼尼丁受体。但其具有较大分子量,不易表达。
Puente等[11]克隆了编码烟芽夜蛾RyR C-末端1 172个氨基酸,通过比对发现其与哺乳动物3种亚型的RyRs具有45%~47%的同源性,与桃蚜(M.p.)、棉蚜(A.g.)、玉米飞虱(P.m.)、果蝇(D.m.)的同源性分别为76.8%、77.5%、79.6%、78.2%。而本文中小菜蛾RyR基因与其他物种同源性关系分析表明,小菜蛾RyR与烟芽夜蛾RyR和蚕相似性高达92%,这与3种昆虫同属鳞翅目有关。
根据预测的氨基酸二级结构分析,小菜蛾鱼尼丁受体基因C端存在6个跨膜区域,这也是鱼尼丁受体基因的特性之一。有研究认为鱼尼丁受体基因存在6~8个跨膜区域,也有研究认为存在4~12个跨膜区域[12-14],然而鱼尼丁受体基因跨膜区域的特定结构尚未明确。Puente等[11]研究表明烟芽夜蛾C-末端存在5个较高的疏水性区域,这5个区域在氨基酸序列的位置为:516~532(M′)、704~720(M1)、782~798(M2)、972~988(M3)和1 054~1 070(M4),即为5个跨膜区域。烟芽夜蛾RyR与小菜蛾RyR跨膜区域的不同,可能是由于计算方式不同引起的。小菜蛾RyR基因6个跨膜区的亲水性指数为1.5~2.6,然而TM4的亲水性指数仅0.6,这与Zorzato等[14]推测的12个跨膜区域的亲水性指数在0.8~2.9之间基本一致。
Puente等[11]研究还表明在烟芽夜蛾RyR跨膜区M3和M4之间存在一个钙离子释放通道的成孔结构域GVRAGGGIGD,同样的结构域也存在于小菜蛾RyR基因C-末端的TM5和TM6之间。然而在兔心肌RyR中也存在这个结构,但是GXRXGGG-XGD第8个氨基酸残基I在兔心肌RyR中为第4897个氨基酸 T[15]。
此外,一个重复序列出现4次不仅体现在小菜蛾鱼尼丁受体基因中,也存在于脊椎动物的骨骼肌和心肌鱼尼丁受体中[14-15]。第一个重复区域有94个残基,位于859~952氨基酸之间;第二和第三个重复区域均有95个氨基酸残基,位于972~1 066和2 838~2 932氨基酸之间;第四个重复区域有89个氨基酸,位于2 964~3 052氨基酸之间。比对重复序列(如图3),根据二级结构预测,小菜蛾鱼尼丁受体基因的重复序列是α-α型,而在RyR1和RyR2的重复序列呈现的是β-α-β-α型。在小菜蛾鱼尼丁受体基因中4个重复序列的相似性有33%,在哺乳动物RyR1和RyR2中重复序列的相似性为28%,然而这一区域的功能尚未得到验证。
微摩尔浓度的细胞质Ca2+可以激活RyRs,毫摩尔的Ca2+可以抑制RyRs。这些双向作用可能是由不同亲和性的Ca2+结合位点的相互作用引起。在鳞翅目昆虫的RyRs中存在PPPEPTEEE氨基酸束,而在兔RyR1这也存在一个脯氨酸和谷氨酸丰富的氨基酸束PEPEPEPEPEPE(4 488~4 499)[16]。45Ca2+的平衡结合试验表明龙虾 RyR与哺乳动物RyR1和RyR2的相应此结构域包含了Ca2+钝化RyR的结合位点即位于EF-hand结构域钙离子结合位点[17]。

图3 重复序列比对结果图
ATP与RyR1存在3个可能的结合位点,结合位点的氨基酸序列为GXGXXG[18]。在烟芽夜蛾的RyR中3个位点分别为GVGLEG、GEGGEG、GSGESG。而小菜蛾中仅有1个相似位点位于RyR的4 005~4 010(GVGLEG)。此外与烟芽夜蛾其余两个位点的位置,小菜蛾的氨基酸序列分别为4 683~4 688(AEPGEG),4 710~4 715(GSGEE-)。在后两个位点区域出现了一个突变位点,一个缺失位点,可能是由于物种间的差异造成,或者是ATP与小菜蛾RyR就存在一个结合位点。
另一个对哺乳动物RyRs具有重要调节作用的是具有双向功能的钙调蛋白。脱钙钙调蛋白(Ca2+游离钙调蛋白)激活的同时,钙调蛋白(Ca2+结合钙调蛋白)抑制RyR钙离子通道[19]。根据有关研究推断4124HS RLWDAVGGFLFLFSHMQDKLSKH4148可能是钙调蛋白与RyR的结合位点[11]。
Ulich等[20]认为氟虫双酰胺与RyR的结合不受鱼尼丁的影响,Kenta等[21]研究表明,氟虫双酰胺与鳞翅目昆虫RyR的结合位点在C-末端4 111~5 084氨基酸之间,因此推测氟虫双酰胺与RyR的结合位点不同于鱼尼丁的结合位点。可见,有关二酰胺类杀虫剂的作用位点仍需进一步研究。因此小菜蛾鱼尼丁受体基因的克隆为此提供了有利的研究基础,也为今后小菜蛾对二酰胺类化合物产生的抗性研究提供理论支持。
[1]David B S,Daniel C,Timothy R C.Insect ryanodine receptors:molecular targets for novel pest control chemicals[J].In-vertebrate Neuroscience,2008,8(3):107-119.
[2]Greg T H,Melissa Z,Paula G M.Feeding cessation effects of chlorantraniliprole,a new anthranilic diamide insecticide,in comparison with several insecticides in distinct chemical classes and mode-of-action groups[J].Pest Management Science,2009,65:969-974.
[3]Hirooka T,Nishimatsu T,Kodama H,et al.The biological profile of flubendiamide,a new benzenedicarboxamide insecticide[J].Pflanzenschutz-Nachrichten Bayer,2007,60:183-202.
[4]Ralf N.Insecticide mode of action:return of the ryanodine receptor[J].Pest Management Science,2006,62:690-692.
[5]Timothy C,Daniel C,Steven G,et al.Isolation and use of ryanodine receptor:US 7655395[P].2010-02-02.
[6]Sun Jingyan,Liang Pei,Gao Xiwu.Cross-resistance patterns and fitness in fufenozide-resistant diamondback moth,Plutella xylostella(Lepidoptera:Plutellidae)[J].Pest Management Science,2012,68(2):285-289.
[7]Wang Xinling,Li Xiangyong,Shen Aidong,et al.Baseline susceptibility of the diamondback moth(Lepidoptera:Plutellidae)to chlorantraniliprole in China[J].Journal of Economic Entomology,2010,103:843-848.
[8]Sukonthabhirom S,Dumrongsak D,Jumroon S,et al.Update on DBM diamide resistance from Thailand:causal factors and learnings[DB/OL].(2011-3-25)[2011-08-10].http:∥www.irac-online.org/resources-2/document-library/#Posters.
[9]杨峰山.小菜蛾Bt蛋白受体基因克隆及序列分析[M].哈尔滨:黑龙江大学出版社,2009:37-40.
[10]Hamilton S L M,Serysheva I I.Ryanodine receptor structure:progress and challenges[J].The Journal of Biological Chemistry,2009,284(7):4047-4051.
[11]Puente E,Suner M,Evans A D,et al.Identification of a polymorphic ryanodine receptor gene from Heliothis virescens(Lepidoptera:Noctuidae)[J].Insect Biochemistry and Molec-ular Biology,2000,30:335-347.
[12]Guoguang D,Bimal S,Vijay K K,et al.Topology of the Ca2+release channel of skeletal muscle sarcoplasmic reticulum(RyR1)[J].Proceedings of the National Academy of Sciences,2002,99:16725-16730.
[13]Hiroshi T,Seiichiro N,Takeshi M,et al.Primary structure and expression from complementary DNA of skeletal muscle ryanodine receptor[J].Nature,1989,339:439-445.
[14]Francesco Z,Junichi F,Kinaya O,et al.Molecular cloning of cDNA encoding human and rabbit forms of the Ca2+release channel(ryanodine receptor)of skeletal muscle sarcoplasmic reticulum[J].The Journal of Biological Chemistry,1990,265:2244-2256.
[15]Kinya O,Huntington F W,Vijiay K K,et al.Molecular cloning of cDNA encoding the Ca2+release channel(ryanodine receptor)of rabbit cardiac muscle sarcoplasmic reticulum[J].The Journal of Biological Chemistry,1990,265:13472-13483.
[16]Wayne S R C,Lin Z,David H M.Antibodies as probes for calcium activation sites in the calcium release channel ryanodine receptor of rabbit skeletal muscle sarcoplasmic reticulum[J].The Journal of Biological Chemistry,1993,268:13414-13421.
[17]Xiong Hui,Feng Xiaoyong,Gao Ling,et al.Identification of a two EF-hand Ca2+binding domain in lobster skeletal muscle ryanodine receptor/Ca2+release channel[J].Biochemistry,1998,37:4804-4814.
[18]Rik K W,Wim G J H.Predicted nucleotide-binding properties of p21protein and its cancer-associated variant[J].Nature,1983,302:842-844.
[19]Ashutoshe T,Le X,Geoffrey M,et al.Calmodulin activation and inhibition of skeletal muscle Ca2+release channel(ryanodine receptor)[J].Biophysical Journal,1995,69:106-119.
[20]Ulich E K,Peter L,Nicole L,et al.Phthalic acid diamides activate ryanodine sensitive Ca2+release channels in insects[J].Cell Calcium,2006,39:21-33.
[21]Kenta K,Shigeki K,Yuichi S,et al.Molecular characterization of flubendiamide sensitivity in the lepidopterous ryanodine receptor Ca2+release channel[J].Biochemistry,2009,48:10342-10352.
猜你喜欢
工业水处理(2022年12期)2022-12-29
中华实验眼科杂志(2022年8期)2022-11-15
湖北农业科学(2022年11期)2022-07-18
植物保护(2022年1期)2022-02-10
农业技术与装备(2022年11期)2022-01-01
实用肿瘤学杂志(2020年4期)2020-12-08
医药前沿(2020年28期)2020-12-02
上海农业科技(2020年5期)2020-10-24
蔬菜(2018年9期)2018-09-21
中国老年学杂志(2015年9期)2015-01-31
