
海南新纪录种东方蜈蚣藻(红藻门)的形态学观察和rbcL基因序列分析
2012-10-23 03:02王宏伟盛英文栾日孝
海洋科学 2012年7期
张 雯, 王宏伟, 赵 丹, 盛英文, 栾日孝
(1. 辽宁师范大学 生命科学学院, 辽宁 大连 116029; 2. 大连自然博物馆, 辽宁 大连 116023)
海南新纪录种东方蜈蚣藻(红藻门)的形态学观察和rbcL基因序列分析
张 雯1, 王宏伟1, 赵 丹1, 盛英文1, 栾日孝2
(1. 辽宁师范大学 生命科学学院, 辽宁 大连 116029; 2. 大连自然博物馆, 辽宁 大连 116023)
通过形态结构观察、生殖器官的发育和rbcL序列分析相结合的方法, 发现了分布于海南省陵水新村的东方蜈蚣藻(Grateloupia orientalis S.-M. Lin et H.-Y.Liang)为海南新纪录种, 该物种具有以下特征: 藻体质地凝胶状或软骨质; 皮层外部由5~6层念珠状的细胞构成, 内部由1~2层星形细胞构成; 生殖枝丛由两个分枝组成; 果孢子囊分布在除藻体基部外的皮层内。
海膜科(Halymeniaceae); 东方蜈蚣藻(Grateloupia orientalis S.-M. Lin et H.-Y.Liang); 新纪录种;形态学观察; rbcL
蜈蚣藻属(Grateloupia C.Agardh)隶属于红藻门(Rhodophyta)隐丝藻目(Cryptonemiales)海膜科(Halymeniaceae), 由 C.Agardh在 1822年基于Grateloupia ornata C.Agardh, G. hystrix C.Agardh和G. filicina (Lamouroux) C.Agardh这3种建立的,是海膜科最大的一个属[1]。属模为 G.filicina(Lamouroux) C.Ag., 主要特征是藻体直立,淡红黄色或紫红色, 黏滑或软骨质, 多轴构造。分为皮层和髓部, 中央髓部由纵走髓丝错综交织着,外围皮层由圆形、椭圆形或不规则细胞组成, 背斜排列。有性藻体雌雄同体或异体, 精子囊群由雄配子体的外皮层细胞形成, 圆柱状, 无色。果孢枝与辅助细胞分别由雌配子体的内皮层细胞产生的枝丝中形成。成熟囊果埋于皮层中, 成颗粒状, 突出于藻体表面。四分孢子囊由孢子体的外皮层细胞形成, 十字形分裂[2]。蜈蚣藻属广泛分布于世界各地的沿海, 全世界报道有约80种[3-10], 中国共报道32种[2]。
作者对海南沿海蜈蚣藻属资源调查时, 对采自海南陵水新村的编号为 LNU092102和 LNU092103的标本进行了形态结构观察和 rbcL基因序列分析,发现与2008年林绣美等[11]报道的产自台湾林园的新种东方蜈蚣藻(G. orientalis S.-M. Lin et H.-Y.Liang)完全一致, 由于在海南省为首次发现, 因此确认该种为海南新纪录种。
1 材料与方法
1.1 形态学观察
材料采自海南省陵水新村。分别制作成腊叶标本、10%福尔马林溶液液浸标本和硅胶干燥标本, 用于形态观察、制作冰冻切片和DNA提取。其中, 冰冻切片用 0.5% (w/v)的甲基蓝染色, 并用 Olympus BH2数字显微镜观察, 用Nikon HFX-ⅡA照相机拍照记录结果。标本保存于辽宁师范大学生命科学学院植物标本室中(LNU)。
1.2 分子序列分析
DNA的提取、PCR扩增以及引物设计参照Wang等[4]并改进的方法。引物组合方式为: (1)F8—R1381-ii, (2)F1—R1150, F735—R1381-ii(表 1)。
从 GenBank 网站(http://www.ncbi.nlm.nih.gov)下载并选取了16个物种的基因序列进行rbcL序列分析(表 2), 包括 14个蜈蚣藻属(Gratelouopia)的物种, 2个外群物种(Halymenia floresia, Polyopes constrictus)。MEGA 4.0软件用于进行碱基差异度的分析以及系统树的构建, Bootstrap值为 1000。采用Kimura的双参数法计算距离矩阵[12], 采用NJ(Neighbor-Joining)法和 MP(Maximum Parsimony)法构建系统树。

表1 rbcL序列扩增所需引物Tab. 1 Primers used for amplifying rbcL sequences

表2 rbcL序列分析所用海藻种类和基因库编号列表Tab. 2 The species used for rbcL sequence analysis and gene number list
2 结果
2.1 藻体外部形态
藻体深红色或棕色, 凝胶状或软骨质, 由 1~15个圆柱状或亚圆柱状的主枝组成, 高达16 cm, 基部具圆饼状的固着器, 直径2~7 mm。单生或数个集生在一起, 集生藻体的固着器紧密连接在一起。主枝长5~12 cm, 常为亚圆柱状, 直径 0.7~1.5 mm×2~3 mm,具有不规则的羽状分枝, 分枝长 8~20 mm, 直径0.2~0.3 mm×0.3~1.5 mm, 配子体和四分孢子体是同型的(图1a和图1b)。

图1 东方蜈蚣藻Fig. 1 External morphology of G.orientalisa. 雌配子体(LNU092102); b. 四分孢子体(LNU092103)a. Female gametophyte (LNU092102); b. Tetrasporic plant (LNU092103)
2.2 内部结构
幼体和成熟藻体的内部结构有细微的变化。幼枝内部中空, 髓丝松散排列, 外皮层由四到五层念珠状的细胞构成(图2a)。随着分枝直径的增加, 外皮层细胞经纵向和横向的分裂, 成为六到八层细胞, 髓丝伸长并紧密缠绕在一起, 由次生纹孔联系相连接。

图2 东方蜈蚣藻的横切面结构和生殖结构(ac代表辅助细胞)Fig. 2 Transverse section and reproductive structures of G. orientalis.(The ac is auxiliary cell )a. 雌配子体横切面观; b~c. 辅助细胞生殖枝丛切面观; d~e. 囊果的发育; f~g. 正在分裂的四分孢子; h. 成熟的四分孢子囊a. Cross-section through a branchlet female gametophyte; b~c. Lateral view of mature auxiliary cell ampulla; d~e. Developing cystocarp;f~g. Dividing tetrasporangium; h. Fully developed tetrasporangium
2.3 生殖结构
果孢枝和辅助细胞生殖枝丛分别始于基部和近基部的内皮层(图 2b)。辅助细胞原始生殖枝丛横向分裂形成一个长达三个细胞的长度的枝丝, 果孢枝和辅助细胞生殖枝丛继续分裂并向外皮层延伸。当原始生殖枝丛达到 5~6个细胞的长度, 次生生殖枝丛的第一个细胞扩大, 行使辅助细胞的功能。受精后, 受精卵核通过联络丝从果孢枝生殖枝丛发出,到辅助细胞。果孢枝和辅助细胞生殖枝丛在外形上相同。一个完整发育的辅助细胞生殖枝丛有两个分枝, 每个分枝可达 7~10个细胞的长度(图 2b和图2c)。
辅助细胞接受了受精卵核后, 经横分裂, 产生一原产孢丝细胞, 由它连续斜分裂形成产孢丝基部细胞。基部细胞继续横分裂, 形成产孢丝。当产孢丝聚集到 5~6个细胞的时候, 在辅助细胞和产孢丝之间的纹孔联系扩大。接着, 产孢丝基部细胞与第一次分裂后的辅助细胞融合, 形成一个小的融合胞。髓丝和融合胞之间形成次级纹孔联系。产孢丝附近的髓丝逐渐缠绕, 并通过次级纹孔联系相互连接形成一个丝状囊果被, 顶端有时可以看到一囊孔。产孢丝从末端开始成熟, 最后形成为梨形或椭圆形的果孢子囊。成熟囊果中, 产孢丝除基部细胞外全部形成果孢子囊, 宽 9~10 μm, 长 11~12 μm(图 2d-e)。
四分孢子始于内皮层细胞, 十字形分裂(图2f-h)。成熟的四分孢子囊长 34~38 μm, 宽 12~15 μm。精子囊母细胞由表皮细胞形成, 第一次纵向分裂, 伸长,再横分裂, 产生精子囊。
2.4 东方蜈蚣藻与其相似种的形态学比较
通过东方蜈蚣藻与其相似种进行形态学特征比较(表 3), 可以看出东方蜈蚣藻与繁枝蜈蚣藻较容易分辨: 东方蜈蚣藻具不规则羽状分枝; 繁枝蜈蚣藻具不规则叉状分枝或互生分枝。东方蜈蚣藻和蜈蚣藻(G. filicina)在外形上较难分辨, 且东方蜈蚣藻曾经被当作G. filicina[13-14]研究, 但是, 可以通过其藻体直径和皮层数分辨: 东方蜈蚣藻藻体直径为0.2~0.3 mm×0.3~1.5 mm, 皮层为 6~9 层;G. filicina藻体直径为350~450 μm, 皮层为5~8层。东方蜈蚣藻与链状蜈蚣藻在外形上极为相似, 很难分辨。
习性: 生长在中、低潮带岩石上或潮下带0.5~2 m的岩石上。产地: 中国海南省陵水新村、中国台湾林园模式标本产地: 中国台湾林园

表3 东方蜈蚣藻及其近似种的形态学比较Tab. 3 Comparison of morphological features between Grateloupia orientalis and its similar species
2.5rbcL分析
全序列比对结果如表 4所示。编号为LNU092102和 LNU092103的标本与东方蜈蚣藻三者之间无碱基差异, 与属模蜈蚣藻(G. filicina)的碱基差异为 8.2%(72 bp), 与繁枝蜈蚣藻(G. ramossissima)的碱基差异为 8.3%(73 bp), 与链状蜈蚣藻(G.catenata)的碱基差异为 8.1%(67 bp), 与蜈蚣藻属内其他各种的碱基差异为 8.65%~9.3%(79~91 bp), 与外群(Polyopes lancifolius,Halymenia floresia)的碱基差异为 10.0%~10.4%(118~127 bp)。

图3 NJ法构建的系统发育树Fig. 3 The phylogeny tree based on Neighbor-Joining比例尺为0.01, 表示遗传距离; 分支上的数字表示支持率(1000次重复), 支持率小于50%的没有给出Scale bar=0.01 substitutions per site. Numbers at nodes are bootstrap values; the numbers lower than 50% are not shown
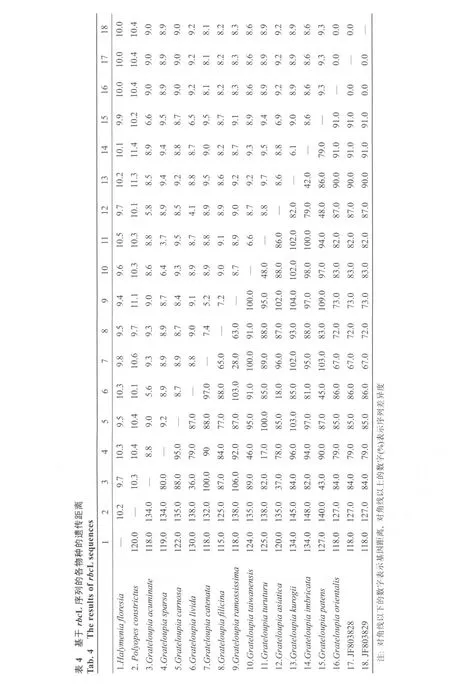
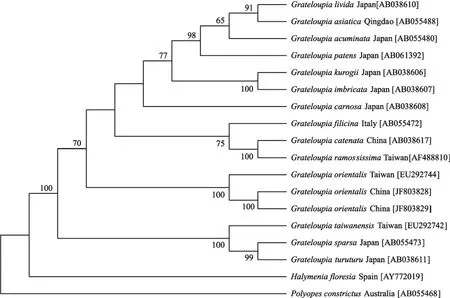
图4 MP法构建的系统发育树Fig. 4 The phylogeny tree based on Maximum Parsimony分支上的数字表示支持率, 支持率小于50%的没有给出Numbers at nodes are bootstrap values; the numbers lower than 50% are not shown
从NJ系统树(图3)和MP系统树(图4)中可以看出, 18个海膜科的物种聚集为3个大的进化支, 其中,编号为LNU092102, LNU092103的标本与蜈蚣藻属内的其他各种聚集为一个大的进化支, 与蜈蚣藻属中的属模蜈蚣藻(G. filicina)、东方蜈蚣藻(G. orientalis)、繁枝蜈蚣藻(G. ramossissima)、链状蜈蚣藻(G.catenata)形成一个小分支, 并与东方蜈蚣藻(G. orientalis)组成一个单独的分支, 区别于其他物种, 其支持率达100%。
3 讨论
蜈蚣藻属(GrateloupiaC.Agardh)是海膜科(Halymeniaceae)中最大的一个属, 其外部形态变化很大。许多分类学家都不同程度地对蜈蚣藻属做了分类学研究。
2008年, 林绣美等[11]首次报道了分布在台湾林园的新种东方蜈蚣藻(G. orientalisS.-M. Lin et H.-Y.Liang), 并描述了该种的形态结构特征。本文采用形态结构观察和rbcL基因序列分析相结合的方法,对海南沿海蜈蚣藻资源进行了调查, 发现编号为LNU092102和 LNU092103的海藻样本的形态结构特征与林绣美报道过的东方蜈蚣藻相同, 经rbcL基因序列分析, 二者无碱基差异, 并且在系统树上也聚为一个进化支, 因此断定该种即为东方蜈蚣藻。由于该种只在中国台湾有过报道, 在海南省为首次发现, 故将其确定为海南新纪录种。
对该藻进行的内部结构观察的结果显示, 其果孢枝细胞和辅助细胞生殖枝丛是典型的蜈蚣藻型,并与东方蜈蚣藻具有相同的结构特点: 藻体质地凝胶状或软骨质; 皮层外部由 5~6层念珠状的细胞构成, 内部由1~2层星形细胞构成; 生殖枝丛由两个分枝组成; 果孢子囊分布在除藻体基部外的皮层内;受精后, 产孢丝细胞邻近的内皮层细胞, 产生次生髓丝, 最后发育为囊果被。
通过东方蜈蚣藻与其相似种的形态学特征比较,可以看出东方蜈蚣藻与繁枝蜈蚣藻较容易分辨, 与蜈蚣藻(G. filicina)和链状蜈蚣藻在外形上较难分辨,但与蜈蚣藻(G. filicina)可通过藻体直径和皮层数分辨开来。
rbcL序列分析的结果显示, 编号为LNU092102和LNU092103的标本与东方蜈蚣藻的序列相似度高达100%, 与其外形相似的繁枝蜈蚣藻、蜈蚣藻以及链状蜈蚣藻的差异为种间差异, 与海膜科内海膜属(Halymenia)和海柏属(Polyopes)的差异较大, 超过了种间差异的范围。在基于rbcL基因(1381 bp)序列的系统发育树(图3和图4)中可以看出, 该藻与东方蜈蚣藻聚成了一个单独的分支, 区别于其他种类, 并且, 东方蜈蚣藻与蜈蚣藻属内的其他种形成了一个大的进化支。这不仅证明了该藻与东方蜈蚣藻是同一物种, 也进一步验证了东方蜈蚣藻 是蜈蚣藻属内的一个独立的物种。
[1] Agardh C A. Species Algarum[M]. Lundae: ex officina Berlingiana, 1822: 169-398.
[2] 夏邦美.中国海藻志. 第二卷. 红藻门. 第三册[M].北京: 科学出版社, 2004: 59-81.
[3] Wang H W, Kawaguchi S, Horiguchi T,et al. A morphological and molecular assessment of the genus Prionitis J. Agardh (Halymeniaceae, Rhodophyta)[J].Phycol Res, 2001, 49:251-262.
[4] Wang, H W, Kawaguchi S, Horiguchi T, et al. Reinstatement of Grateloupia catenata (Rhodophyta, Halymeniaceae) on the basis of morphology and rbcL sequences[J]. Phycologia 2000, 39: 228-237.
[5] Kawaguchi S, Wang H W, Horiguchi T, et al. A comparative study of the red alga Grateloupia filicina (Halymeniaceae) from the northwestern Pacific and Mediterranean with the description of Grateloupia asiatica sp. nov. [J]. J Phycol, 2001, 37:433-442.
[6] Gavio B, Fredericq S. Grateloupia turuturu (Halymeniaceae, Rhodophyta) is the correct name of the non-native species in the Atlantic known as Grateloupia doryphora[J]. Phycol, 2002, 37: 349-360.
[7] Faye E T, Wang H W, Kawaguchi S, et al.Reinstatement of Grateloupia subpectinata (Rhodophyta, Halymeniaceae) based on morphology and rbcL sequences[J]. Phycol Res, 2004, 52: 59-68.
[8] De Clerck O, Gavio B, Fredericq S, et al. Systematics of Grateloupia filicina (Halymeniaceae, Rhodophyta),based on rbcL sequence analyses and morphological evidence, including the reinstatement of G. minima and the description of G. capensis sp. nov.[J]. J Phycol,2005a, 41: 391-410.
[9] De Clerck O, Gavio B, Fredericq S, et al. Systematic reassessment of the red algal genus Phyllymenia(Halymeniaceae, Rhodophyta)[J]. Eur J Phycol, 2005b,40: 169-178.
[10] Wilkes R J, McIvor L M, Guiry M D. Using rbcL sequence data to reassess the taxonomic position of some Grateloupia and Dermocorynus species(Halymeniaceae, Rhodophyta) from the north-eastern Atlantic[J]. Eur J Phycol, 2005,40:53-60.
[11] Lin S M, Liang H Y, Hommersand M H. Two types of auxiliary cell ampullae in Grateloupia (Halymeniaceae,Rhodophyta ), including G. taiwanensis sp. nov. and G.orientalis sp. nov. from Taiwan based on rbcL gene sequence analysis and cystocarp development[J].Journal of Phycology, 2008, 44(19):196-214.
[12] Kimura M. A simple method for estimating rate of base substitutions through comparative studies of nucleotide sequences[J]. Journal of Molecular Evolution, 1980, 16:111-120.
[13] Chiang Y M. Marine algae of northern Taiwan(Rhodophyta)[J]. Taiwania, 1962, 8: 167-180.
[14] Chiang Y M. Notes on marine algae of Taiwan[J]. Taiwania, 1973, 18:13-17.
Morphological observation and rbcL gene sequence analysis of Grateloupia orientalis (Rhodophyta), a new record of the Hainan province
ZHANG Wen1, WANG Hong-wei1, ZHAO Dan1, SHENG Ying-wen1,LUAN Ri-xiao2
(1. College of Life Sciences, Liaoning Normal University, Dalian 116029, China; 2. Dalian Natural History Museum, Dalian 116023, China)
Nov.,12,2011
halymeniaceae; Grateloupia orientalis; new record; morphological observations; rbcL
The Grateloupia orientalis S.-M. Lin et H.-Y.Liang collected from Lingshui, Hainan province has been confirmed to be a new record in the Hainan province through morphological observations, development of reproductive structures and molecular phylogenetic studies. This species has the following morphological features:Thallus are gelatinous to cartilaginous in texture; cortex is composed of five to six layers of outer moniliform cells and one to two layers of stellate-shape inner cells; ampullae is composed of only two orders of unbranched filament;carposporangia distribute in all the cortex except the bottom of thallus.
Q943
A
1000-3096(2012)07-0109-08
2011-11-12;
2012-03-23
国家自然科学基金项目(30870161)
张雯(1986-) 女, 山东莱芜人, 硕士研究生, 从事海藻分类学研究, E-mail: zhangwen111213@126.com; 王宏伟, 通信作者, 教授,博士, 从事海藻分类、多样性及其分子系统学研究, E-mail: kitamiwang@yahoo.com.cn
(本文编辑:梁德海)
猜你喜欢
中国海洋大学学报(自然科学版)(2022年8期)2022-08-15
广西科学(2022年1期)2022-04-12
意林原创版(2021年9期)2021-09-28
教学考试(高考生物)(2020年6期)2020-11-23
意林·少年版(2020年23期)2020-01-15
食品与生物技术学报(2020年8期)2020-01-06
水产科学(2019年4期)2019-07-23
学苑创造·B版(2019年5期)2019-06-14
科学24小时(2019年5期)2019-06-11
科技创新与应用(2017年20期)2017-07-15
