
朽木中白腐真菌的选育及对木质纤维素降解性能研究
2013-01-29 03:47李海红李红艳
陕西科技大学学报 2013年4期
李海红, 李红艳
(西安工程大学 环境与化学工程学院, 陕西 西安 710048)
0 引言
农作物秸秆作为一种宝贵的可再生资源,由于非水溶性的秸秆木质素与半纤维素以共价键结合形成的紧密结构,将纤维素分子包埋在其中,导致了秸秆的难生物降解特性[1,2],限制了秸秆的资源化利用.因此,缩短秸秆降解周期的关键是木质素的高效降解.目前生物预处理被证明是木质素降解的有效途径,自然界中可以腐蚀和分解秸秆、树叶及树木等高纤维物质的微生物很多,其中白腐真菌作为一个庞大的微生物家族,以其特殊的生理生化机制和强大的降解能力,越来越引起人们的关注.
章燕芳等人[3]研究发现,白腐真菌与其它的微生物相比,在对木质素的降解方面表现出很强的优势.白腐菌是一类丝状担子真菌[4,5],喜好温暖湿润的环境,多见于热带雨林的阔叶树或针叶树的枯木上群生,能够分泌胞外氧化酶降解木质素,被认为是最主要的木质素降解微生物.而优良菌株是分解或转化木质素的关键.
为获得可用于秸秆还田条件下接种、促进秸秆快速腐解的白腐真菌,本实验从众多朽木上大量取样,通过富集培养、划线初筛以及复筛鉴定选育出2株具有木质素氧化酶的菌株,利用滤纸条和稻草秸秆对其木质纤维素降解能力进行研究,得到了具有降解秸秆纤维素和木质素的候选白腐真菌,为秸秆的综合利用提供了工作基础和理论参考.
1 材料与方法
1.1 菌源
选择秦岭南骊宁陕县未开发森林为样品采集地点,菌种的采集对象主要锁定在多年的朽木上,用刀片将含菌的朽木从表层割下,并用标签标记,保存于冰箱中.
1.2 培养基
1.2.1 选择培养基
主要成分:KH2PO4·3H2O 1 g/L;NaH2PO4·7H2O 0.2 g/L;MgSO4·7H2O 0.5 g/L;CaCl20.1 mg/L;FeSO4·7H2O 0.4 mg/L;CuSO4·7H2O 0.2 mg/L;ZnSO4·7H2O 1.4 mg/L;(NH4)C4H4O60.1 g/L;葡萄糖1 g/L;VB10.1 mg/L;pH 5.5,此培养基又叫专项培养基.
1.2.2 土豆培养基(PDA)
主要成分:马铃薯 200 g;葡萄糖 20 g;K2HPO43 g;MgSO4·7H2O 1.5 g;蒸馏水 1 000 mL;pH 5.5;VB1微量.用于菌类的增殖扩大培养.
1.2.3 滤纸条鉴定培养基
主要成分:(NH4)2SO41 g/L;KH2PO4·3H2O 1 g/L;MgSO40.5 g/L;K2HPO42 g/L;酵母膏 0.1 g/L;灭菌后的滤纸条(1*7 cm)一条;蒸馏水1 000 mL;自然 pH.经恒温振荡培养,计算滤纸失重率.
1.2.4 固态发酵培养基
主要成分:稻草粉(40目)3 g;CaSO40.02 g;MnSO40.06 g;接种量10%;12 mL自来水;pH 5.5.
配制好的液体培养基加入一定量的琼脂(1.5~2.0 %),即1 000 mL加入15~20 g后加热搅拌,即可得到固体培养基,接种前于121 ℃下高压灭菌20~30 min.
1.3 实验与方法
1.3.1 白腐真菌的筛选
(1)初筛:用75 %的乙醇将采集的样品浸泡3~5 min进行消毒处理,阴干后对腐木的不同部位剪切分离,把得到的腐木小块混均后,接种到选择固体平板培养基上,一部分腐木块与培养基混合培养;一部分均匀撒到培养基表层,形成对照试验,于30 ℃培养箱培养7 d.
按照白腐菌菌落外形特征、颜色、透明度等,选择疑似白腐真菌的菌落于PDA固体平板培养基上和选择固体平板培养基上经4次交叉划线分离.
(2)复筛:对初筛得到的单菌落进行光学显微镜观察,根据生长曲线的变化观察生物量的增减情况,对木质素氧化酶进行显色反应确定酶存在情况,从而选出具有一定酶活力的白腐真菌.
1.3.2 生长曲线的测定
将镜检得到的符合白腐真菌特征的菌种,用打孔器以点状的形式接种到PDA平板培养基上,置于生化培养箱中30 ℃恒温培养,每12 h测量菌种的直径.按照真菌菌落的观察法,初划分为某几种菌,并以培养时间为横坐标,以菌落直径为纵坐标,绘制白腐真菌生长曲线[6].
1.3.3 锰过氧化酶(MnP)的定性测定
将分离划线筛选出的菌株接种于加入0.1 g/L MnCl2·4H2O的PDA培养基中,于30 ℃培养箱培养7 d,平板上形成黑棕色斑点[7],指示有锰过氧化酶的合成.
1.3.4 木质素过氧化酶(LiP)的定性测定
本实验采用的是亚甲基蓝(MB)法[8],通过指示剂颜色的变化测定LiP的存在.LiP的定性反应混合液2.7 mL:2.2 mL酶液,0.1 mL的1 mmol/L的MB,0.3 mL的0.5 mol/L的酒石酸钠缓冲成分( pH 4.0),加入0.1 mL的4.5 mmol/L的H2O2启动反应.将有酶液体系的颜色变化,与以蒸馏水取代酶液的体系的空白进行比较,对LiP进行定性测定.在 LiP 的催化下,亚甲基蓝MB发生脱甲基反应,转化为天青C,同时,MB由略显绿的蓝色,变为紫蓝色的天青C.
粗酶液的制备:取出2.0 g发酵有菌的培养基,加入25 mL的蒸馏水,振荡浸提2 h.过滤去除残渣,离心后(3 000 r/min,10 min)取上清液用于测酶活[9].
1.3.5 滤纸条崩解实验
称取剪成1 cm 宽的滤纸条0.50 g 作为白腐真菌生长的唯一碳源,将其放置于100 mL 除去葡萄糖的滤纸条培养基中,接种5 mL 筛选菌液放在30 ℃条件下静置培养6 d,进行4 000 r.min-1离心,去除上清液后,用盐酸和硝酸混合液(1∶1)冲洗而消除菌体,离心,清水洗涤,离心后105 ℃烘干并称重,计算失重量和失重率[10],并观察滤纸降解情况,从中挑选出滤纸降解快、失重率高的菌种做进一步研究.
滤纸失重率(%)=(A-B)/A×100%
(1)
其中:A—降解前滤纸质量,g;B—降解后滤纸质量,g.
1.3.6 固态发酵实验
菌液培养: 将筛选出的菌株在斜面固体PDA培养基中进行种子活化,培养7 d,将活化的斜面接种入液体土豆培养基中,培养5~7 d,即得种子液(培养温度为30 ℃).
稻草基质发酵培养: 配制固态发酵培养基并灭菌,按10%的接种量接入上述培养好的种子液,30 ℃静置培养.每5 d取固体发酵样品,用150 mL pH 5.0的醋酸-醋酸钠缓冲液浸泡3 h,过滤,滤液待用,滤渣于60 ℃烘干,用于测定其木质素的含量.
木质素含量的测定方法:采用文献[11]中的差重法计算,并计算其降解率.
木质素降解率(%)=(1-A/B)×100 %
(2)
其中:A—降解后木质素含量,g;B—降解前木质素含量,g.
2 结果与讨论
2.1 菌株的初筛
实验对朽木样品进行乙醇消毒预处理,可以初步杀死一些杂菌,采用葡萄糖为碳源,主要用于担子菌以外的其他菌种,尤其是对细菌和霉菌具有抑制作用,但其营养成分却能满足白腐真菌生长发育的选择平板培养基进行富集培养.对5种菌源A~E对照培养,结果发现将朽木小块均匀洒到培养基表层更有利于白腐真菌的生长,这正符合其好氧的特性.

表1 菌落特征表
在10个培养基上,根据白腐真菌的菌落特征(菌体呈绒毛状或粉状,菌落扁平、生长均匀,有同心圆或放射线)选出长势较好,疑似白腐菌的11株菌落以及1株未知菌落,依次编号(未知菌落的编号为L12),如表1所述.挑取每株菌落于PDA平板培养基上和选择平板培养基上经过4次交叉划线分离,在培养基中的第三条线和第四条线上均有一个一个的单独菌落生成.实验结果发现,相比较选择培养基里的菌落的生长缓慢、稀疏、颜色淡,PDA培养基里的菌种则生长尤为茂盛、密实且洁白有光泽,不足处是因为其适合很多菌的生长,所以培养过程会被杂菌感染,出现绿色的菌落.不同的培养基有不同的功效,所以结合起来使用更方便菌种的纯化,而且缩短了分离周期.
2.2 菌株的复筛
2.2.1 镜检
对初筛选出的12组菌种在光学显微镜下进行观察,进一步确定菌株的个体形态,结果发现编号为B2、C3、E5、I9、K11的培养基里的白色菌落经染色后,在100倍的目镜下观察到大量的发达菌丝,丝内含有大量的细胞核,且菌体存在一定量的分生孢子和孢子囊,如图1所示.

图1 菌落镜检形态图
从图1可以看出,除C3菌和K11菌有少量的锁状结构,其余菌株几乎无隔膜菌丝,无锁状联合,这符合相关资料对白腐真菌的形态描述,而其他7种菌落的镜检结果多为点状.所以选择上述5种菌株进行下一步鉴定.
2.2.2 生长曲线的测定
实验每隔12 h测定一次菌落的生长曲线,实验结果如图2所示.
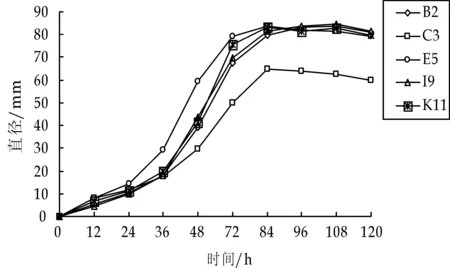
图2 生长曲线
从图2中可以看出,C3菌生长速度较缓慢,而E5菌的生长速度最快,其他3种菌生长速度基本一致,即开始随天数的增加,其生物量呈线性增加;在一定天数后,其生物量增加变缓,有些菌株生物量开始减少,出现这种情况的原因是随着天数的增加,液体培养基的营养成分消耗而使菌体进入次生代谢,或是菌株出现了自溶现象等.整体而言,白腐真菌的生长周期大致如下:调整期为0~1 d,营养期为2~3 d,停滞期自第3天左右开始,并延续1~2 d,随后进入繁殖期,生长基本停止,甚至发生生物量的下降.
2.2.3 木质素氧化酶的测定
白腐真菌在适宜的条件下,其菌丝首先用其分泌的超纤维氧化酶溶解秸秆表面的蜡质,然后菌丝进入秸秆内部,应答合成多种酶,并分泌到细胞外,构成降解系统的主要成分.其中关键的两类过氧化物酶——锰过氧化酶和木质素过氧化酶,在分子氧的参与下,依靠自身形成的H2O2,触发启动一系列自由基链反应,实现对秸秆中木质素无特异性的彻底氧化.因此,在选菌过程中,对这两类酶进行测定很重要,测定结果如下.
(1)锰过氧化酶(MnP)的测定:将5种菌接种于加入0.1 g/L MnCl2·4H2O于PDA培养基中,经过24 h的培养,发现B2菌和E5菌的培养基上出现黑色斑点,说明这两种菌产生了 MnP ,如图3所示,而其余的培养基上无此情况.
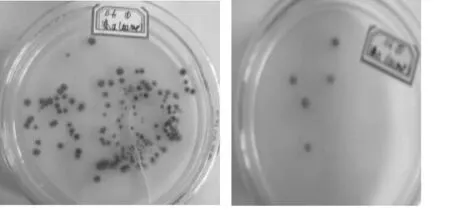
图3 B2菌(左)和E5菌(右)的MnP测定结果
对比图3两图中的斑点数,发现B2菌所产生黑棕色斑点明显多于E5菌.分析原因有两方面:一方面与初始接种量有关,另一方面可能是由于菌株源于不同朽木,由于各种自然原因而导致锰过氧化酶的产量差距.
(2)木质素过氧化酶(LiP)的测定:应用亚甲基蓝法,取6个试管,依次编号,加入2.2 mL木质素过氧化酶,试管6加入2.2 mL的蒸馏水作为对照,其余试剂加入量相同,当加入H2O2启动反应后,发现试管1和试管3由略显绿的蓝色变成紫蓝色,表明B2菌和E5菌存在LiP.
2.3 滤纸条崩解实验
此实验主要是分析菌株的纤维素降解能力,选用滤纸条失重率这一指标对B2、C3、E5、I9、K11号菌株进行进一步的筛选.经过为期6天的观察培养,5个菌株对滤纸条的降解结果如表2所示.
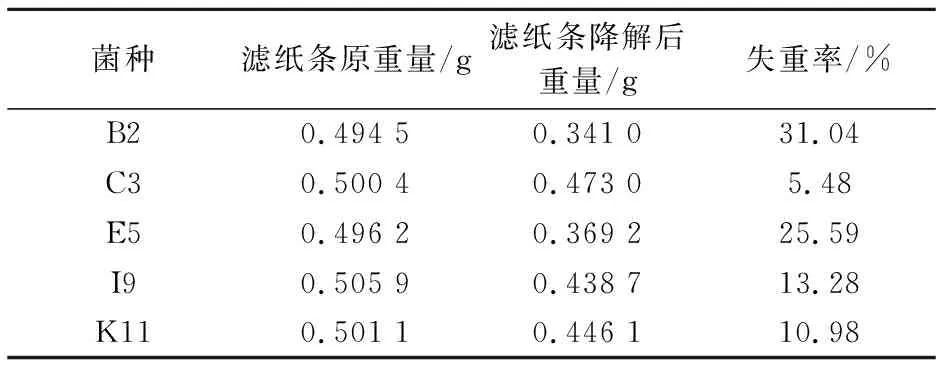
表2 滤纸条降解结果
以失重率>25%为界限,筛选出B2菌及E5菌,其中B2菌的滤纸失重率最大,为31.04%,其次是E5菌,滤纸失重率为25.59%,说明B2菌和E5菌能有效降解滤纸,具有较强的纤维素降解能力,这与菌株的木质素氧化酶存在情况相符合.其余的菌株虽具有一定的降解能力,生物特性也符合白腐真菌的相关描述,但不具有木质素氧化酶,所以降解效率较低,这表明这些菌株可能不是白腐真菌.故选择B2和E5白腐菌菌株,来研究其木质素降解能力.
2.4 固体发酵培养实验
通过差重法测定,两株白腐真菌对玉米秸秆中木质素20天内的降解率如图4所示.

图4 两种菌株固体发酵的木质素降解效率
从图4图形走势可以看出,B2号菌和E5号菌在0~10天的时间内降解速率较快,以后成缓慢减速状态,这与白腐真菌的生长曲线相符合,在降解20天后,B2和E5菌的降解效率分别约为33.7%、30.7%,B2菌的降解速率要高于E5菌,这与MnP的测定结果以及滤纸条失重率的结果相符合.
结果表明,菌株B2和E5对木质素的降解效率和酶存在情况有很大的联系,而这两株菌体能在20 d内对木质素高聚物保持稳定降解,说明菌株B2和E5具有较强的木质素降解能力,可扩大培养,为后续的秸秆综合利用试验提供优良的微生物源.
3 结论
(1)通过分离筛选及鉴定,从来自秦岭山的多年朽木中获得5株形态特征符合白腐真菌的生物特性的菌株,但只有B2菌与E5菌存在木质素氧化酶(LiP和MnP),表明这两种菌株具有一定的木质纤维素降解酶活,属于白腐真菌.
(2)利用滤纸条崩解实验分析菌株的纤维素降解能力,B2菌的滤纸失重率最大为31.04%,E5菌的失重率次之,为25.59%,表明B2菌和E5菌能够有效降解滤纸,具有较强的纤维素降解能力.
(3)通过差重法测定固体发酵实验中木质素降解效率,在25 ℃发酵20 d时, B2菌和E5菌的降解效率分别约为33.7%、30.7%,说明两菌株具有较强的木质素降解能力,且B2菌的降解效果优于E5菌.因此,这两菌株可作为秸秆木质纤维素降解菌进行研究,但是对于木质素酶和降解效率的关系仍需进一步探究.
[1] 黄 茜,黄凤洪,江木兰,等.木质素降解菌的筛选及混合菌发酵降解秸秆的研究[J].中国生物工程,2008,28(2):66-70.
[2] 郁红艳,曾光明,黄国和,等. 木质素降解真菌的筛选及产酶特性[J].应用与环境生物学报,2004,10(5):639-642.
[3] 章燕芳,李华钟,华兆哲,等.木质素过氧化物酶和锰过氧化物酶对染料脱色性能比较[J].环境科学研究,2002,15(5):17-22.
[4] 李慧蓉.白腐真菌生物学和生物技术[M].北京:化学工业出版社,2005:12-49.
[5] 李海红,常华,袁月祥,等.复合菌剂预处理对秸秆厌氧发酵的影响[J].西北大学学报(自然科学版),2012,42(6):949-952.
[6] 张红丹,杜 茜,张正坤,等.放线菌769抑菌谱及液体培养生长曲线的测定[J].中国植保导刊,2010,30(3):4-8.
[7] Yu G C,Wen X H,Qian Y.Production of the ligninolytic enzymes by immobilized phanerochaete chrysosporium in an air atmosphere[J].World Journal of Microbiology and Biotechnology,2005,21(3):323-327.
[8] Jeffries T W,Choi S,Kirk T K.Nutritional regulation of lignin degradation by phanerochaete chrysosporium[J].Appl.Environ.Microbiol.,1981,42(2):290-296.
[9] 陈 合,张 强.菌酶共降解玉米秸秆的工艺研究[J].农业工程学报,2008,24(3):270-271.
[10] Keyser P,Kirk T K,Zeikus J G.Ligninolytic enzyme system of phanerochaete chrysosporium synthesized in absence of lignin in response to nitrogen starvation[J].Journal of Bacteriology,1978,135:790-797.
[11] 薛惠琴,杭怡琼,陈 谊,等.稻草秸秆中木质素、纤维素测定方法的研讨[J].上海畜牧兽医通讯,2001,(2):15.
猜你喜欢
昆明医科大学学报(2020年12期)2021-01-26
上海包装(2019年8期)2019-11-11
上海农业学报(2017年4期)2017-04-10
天津造纸(2016年1期)2017-01-15
中国科技信息(2016年19期)2016-10-25
中国粮油学报(2016年5期)2016-01-23
中国造纸学报(2015年1期)2015-12-16
中国塑料(2014年4期)2014-10-17
应用化工(2014年12期)2014-08-16
汽车零部件(2014年2期)2014-03-11
