
毛竹林地上生物量与胸径的分形关系
2013-11-24 07:08商珍珍周国模杜华强
浙江农林大学学报 2013年3期
商珍珍,周国模,杜华强
(1.浙江农林大学 浙江省森林生态系统碳循环与固碳减排重点实验室,浙江 临安 311300;2.浙江农林大学 环境与资源学院,浙江 临安311300)
毛竹 Phyllostachys edulis 林乔木层年固碳量为 5.097 t°hm-2°a-1[1],通过与同类文献[2-3]对比发现,其单位面积固碳量是热带山地雨林的1.4倍左右,是苏南27年生杉木Cunninghamia lanceolata林的2.0倍多;李惠敏等[4]对杭州市余杭区1985年和1998年森林碳库的动态研究结果表明:1985年和1998年分别占全区森林面积46%和49%的竹林,却占该区当年森林碳储量总量的66%和65%,而且竹林的碳密度是松林、杉木林和经济林的2.0倍左右;李正才等[5]初步估算出中国竹林生态系统的碳储量为整个森林生态系统碳储量的4.05%。因此,竹林在森林生态系统二氧化碳减量方面,乃至对全球碳平衡的贡献开始受到关注。目前,生态学家越来越关注空间格局与生态过程的关系,并采用多尺度方法,从不同层次上分析生态属性在空间上的变化。分形是解释复杂结构和尺度问题的一个优秀的数学工具,能够用来分析数据的各种空间现象,描述现实世界详细的等级层次[6-7],以分形理论为基础的时空格局变化及对复杂性的刻画,也逐渐受到生态学家的关注[8],并在生物多样性、群落结构、空间异质性等研究方面取得显著成果[9-12]。在生物量方面,学者研究了植株地上生物量与株高的分形关系,以及生物空间分布格局的分形特征,为揭示植被生长过程、生物量累积规律及空间格局提供有价值参考[13-16]。Ricotta等[17]研究采用分形理论描述大尺度上归一化植被指数(NDVI,normalized difference vegetation index)年度累积的自相似格局,并估算了净初级生产力(NPP,net primary productivity)的空间分布。本研究将应用分形相关原理与方法,研究毛竹地上生物量与胸径的分形关系,分析不同年龄毛竹林地上部分生物量分形特征,为竹林碳汇研究及经营实践提供参考。
1 研究区域及数据
研究区位于浙江省安吉县境内,地处 30°23′~30°53′N,119°14′~119°53′E。地势自西南山区向东北丘陵平原成喇叭形倾斜展开,海拔为500~1 000 m。气候属亚热带海洋性季风气候,光照充足,气候温和,雨水充沛,四季分明,年均降水量为1 400 mm,年平均气温为15.6℃。全县森林覆盖率为71.1%,拥有山林13.80万hm2,竹林面积为6.97万hm2,其中毛竹林约5.53万hm2,广泛分布于全县。
2 研究方法
2.1 毛竹样地调查与地上生物量估算
样地调查时间段在2008年8月19日到2008年9月3日。采用典型抽样和随机抽样相结合的方法设置了具有不同立地条件、不同立竹度及不同经营状况的55个样地,大小为30 m×30 m,样地基本覆盖整个安吉县不同乡镇毛竹林。样地调查时,利用罗盘仪确定样地边界,闭合差控制在样地周长的1/200,调查内容包括样地经纬度、海拔、坡度、郁闭度、毛竹胸径和年龄(度)。
55个样地毛竹总株数为16 033株,毛竹株数最多的样地有500株,而最少的仅有153株。为快速估算毛竹地上生物量,用毛竹单株生物量估算模型(式1)[18],代入样地调查因子,估算样地每株毛竹生物量并汇总得样地总生物量。模型(1)的决定系数R2=0.937,在0.05置信水平下,模型的预估精度为96.43%,总系统误差为-0.021%,满足生物量估算精度要求。
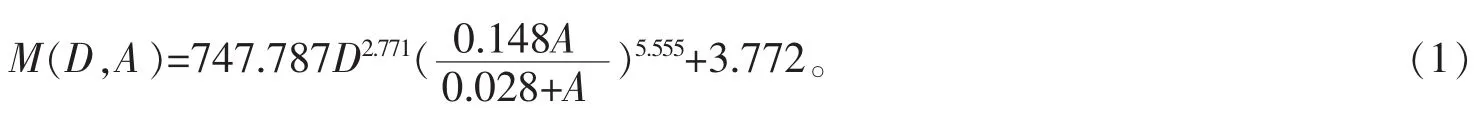
式(1)中:M为生物量(干质量,kg),D为胸径(cm),A表示竹龄(度)。
2.2 分形理论及分形维数计算方法
在不同的尺度上,自然现象常常有相似的特征,即自然界许多目标的大小分布都可以用式(2)表示:

式中:σ为尺度,M(σ)为某尺度 下分形目标的特征数,d为分形维数。由式(2)可知:

现将尺度σ换成λσ,则:

可见:其特征尺度没有发生变化,只是分形对象原来的形状被放大或缩小[19],而分形维数d用它来描述该分形对象的空间分布格局或内在的等级层次、结构。
竹类植物是单子叶植物,没有形成层,竹笋有多少节和多粗,成竹就有多少节和多粗。因此,不同胸径的毛竹,可看成胸径的放大或缩小。结合(1)式,D是影响毛竹单株生物量的主要因素。这样,不同胸径毛竹生物量应该满足式(4),用毛竹胸径D代替尺度σ得:

即,D为尺度,M(D)为尺度D下的分形特征数即生物量。因此,样地内毛竹生物量的变化规律将由其分形维数d控制和描述,成为联系不同胸径毛竹的桥梁。以上就是采用分形研究毛竹地上生物量的理论基础。
3 结果与分析
3.1 毛竹地上生物量与分形维数间的关系
为了保证数据的可比性,随机抽取毛竹150株°样地-1,按上述方法计算各样地地上生物量与胸径的分形关系,并按图1所示计算样地分形维数。研究发现:lg(M(D))和lg(D)之间存在无标度区,无标度区内线性回归曲线相关指数为0.970 9~0.997 1(表1),显著水平很高。表明毛竹地上生物量和胸径之间存在分形特征,即小胸径毛竹与大胸径毛竹地上生物量在一定尺度范围内的生物量空间分布格局具有相似的特征。这一特征通过分形维数定量表达出来,成为不同尺度毛竹生物量特征的桥梁,为研究毛竹生物量及其空间分布提供了重要参考。
为了说明这一点,进一步根据表1计算结果,构建了毛竹地上部分生物量(对数,下同)与分形维数之间的关系模型,如图2。模型相关指数为0.847 4,显著水平为0.01,说明地上部分生物量与分形维数具有很好的线性关系,即随着分形维数增加,生物量随之增加。对于单株毛竹,胸径大小是生物量的关键因素,样地总生物量是不同胸径毛竹累加的结果;而图2的关系表明分形维数成为控制毛竹生物量的因素,也就是说,分形维数将不同胸径毛竹的生物量特征联系起来,反映生物量的累积规律。
分形维数是分形对象的重要参数,它可以定量反映空间数据的复杂性、形状的变化等[6]。分维数越大,分形对象越复杂,其占有空间的能力越强;反之亦然。另外,分形对象在不同尺度 “形”的自相似,使得可以通过分形维数这一重要的参数,模拟分形生长过程。因此,相对仅具有统计意义的胸径分布而言,分形维数更具有空间意义,即分形维数越大,毛竹生物量在空间上积累就越多[15]。就本研究而言,毛竹林生物量对数的空间积累符合图2所示的线性关系,即毛竹林生物量在空间上按指数关系累积,其增长率为1.047 9(图3)。

图1 分形维数计算示意图Figure 1 Algorithm of fractal dimension
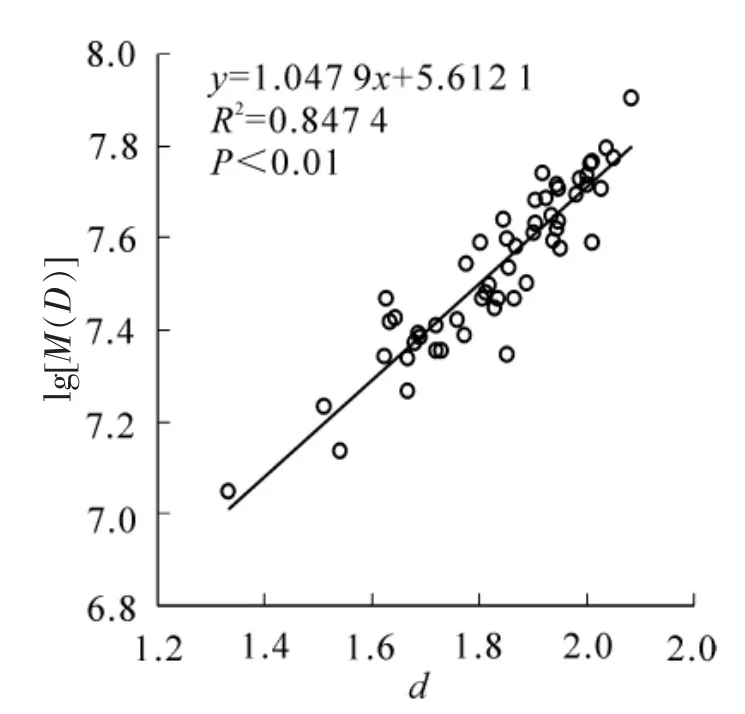
图2 毛竹地上生物量与分形维数关系Figure 2 Relationship between aboveground of Phyllostachys edulis biomass and its fractal dimensions
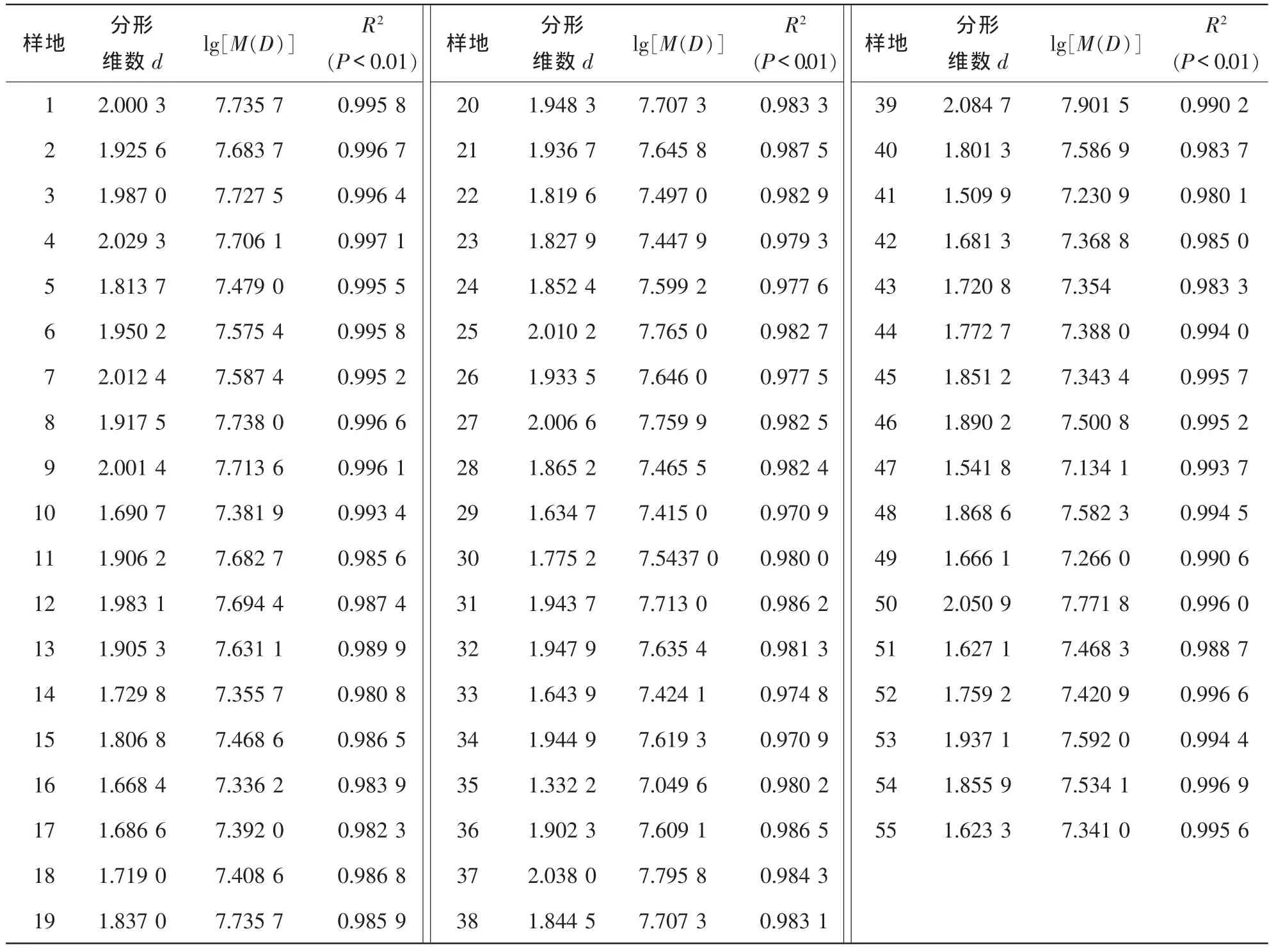
表1 不同样地分形维数与地上生物量Table 1 Fractal dimension and above ground biomass in the 55 plots
3.2 不同度数毛竹地上生物量与胸径分形关系
研究区人工经营毛竹一般保留1度和2度竹,而砍伐3度竹,因此,毛竹林总是处于动态变化之中。在此,分别以不同度数毛竹,研究生物量与胸径的分形关系,从空间上分析生物量的时间动态。
从所有样地中随机抽取毛竹3 000株°度-1,按公式计算毛竹生物量与胸径的分形关系,如图3。可以看出,不同度数毛竹胸径与生物量之间存在无标度区,相关指数R2在0.98以上,线性关系明显,故不同年龄阶段的毛竹胸径与生物量也具有分形特征。其中1度竹遵循分形维数为1.934 6的规律,2度竹遵循分形维数为1.933 9的规律,3度竹遵循分形维数为1.924 4的规律;另外,在无标度区间内,1度,2度和3度毛竹的总生物量分别为7 326.4,7 002.6和7 104.4 kg。
尽管1度毛竹的分形维数和生物量都最大,但t检验表明1度、2度和3度毛竹之间的分形维数和生物量无显著差异,即毛竹生物量在空间分布和累积上没有年龄的差异,基本上遵循分形维数为1.93的规律,接近按2维数方式的变化。从安吉县毛竹林生产经营实践看,当年生毛竹达到3度后基本全部采伐,也就是说3度毛竹的采伐生物量应近似等于1度毛竹的生物量。这样,当3度采伐后,2度变3度,1度变2度,毛竹林生长与收获将保持动态平衡。就所有55个调查样地而言,1度竹的平均生物量为 12.073 1 kg°株-1,2 度竹为 13.060 0 kg°株-1,3 度竹为 12.797 8 kg°株-1,三者之间没有显著差异。
4 结论与讨论
毛竹地上生物量与胸径存在分形特征。相对于仅有统计意义的胸径而言,分形维数将不同胸径的毛竹特征通过相似空间结构联系起来,成为控制毛竹生物量空间分布及累积过程的因素。
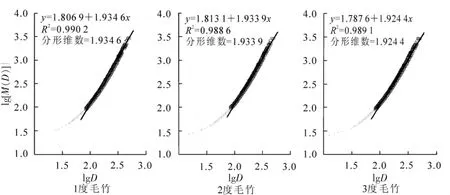
图3 各度毛竹地上生物量与胸径的分形关系(横坐标为胸径对数,纵坐标为生物量对数)Figure 3 Fractal relationships between diameter and aboveground biomass of Phyllostachys edulis with different ages(xcoordinate is natural logarithm of diameter,and y-coordinate is natural logarithm of biomass)
分形维数与生物量之间存在lgb=1.047 9d+5.612 1(b为生物量)的关系,即随着分形维数d的增加,生物量按指数函数规律增长,空间增长率为1.047 9。
1度、2度和3度毛竹地上生物量与胸径之间也存在分形特征,但三者之间生物量在空间分布和累积上没有差异,基本上遵循分形维数为1.93的规律,接近按2维数方式的变化。说明不同度数毛竹生物量在空间上处于动态平衡,符合毛竹林生产经营实践,为毛竹林可持续发展提供理论基础。相对于规则分形而言,自然界的随机分形具有上下两端的限制,即分形性质只存在于无标度区。毛竹地上生物量分形也仅在一定尺度范围内存在,然而,在这一尺度范围类,其生物量空间分布格局上具有自相似的结构,这就有可能通过尺度推移对不同尺度上毛竹生物量在空间上进行预测估算,为毛竹生物量计算提供新的理论。本研究仅从县域尺度上对人工毛竹林进行了研究,大尺度上如整个亚热带毛竹林生物量胸径分形特征需进一步研究;而对天然毛竹林地上生物量空间格局及不同度数毛竹生物量的动态分形也有待进一步深入研究。
[1]周国模,姜培坤.毛竹林的碳密度和碳储量及其空间分布[J].林业科学,2004,40(6):20-24.ZHOU Guomo,JIANG Peikun.Density,storage and spatial distribution of carbon in Phyllostachy pubescens forest[J].Sci Silv Sin,2004,40 (6): 20-24.
[2]李意德,曾庆波,吴仲民,等.我国热带天然林植被C储存量的估算[J].林业科学研究,1998,11(2):156-162.LI Yide,ZENG Qingbo,WU Zhongmin,et al.Estimation of amount of carbon pool in natural tropical forest of China[J].For Res,1998,11 (2): 156-162.
[3]阮宏华,姜志林,高苏铭.苏南丘陵主要森林类型碳循环研究:含量与分布规律[J].生态学杂志,1997,16(6):17-21.RUAN Honghua,JIANG Zhilin,GAO Suming.Preliminary studies of carbon cycling in three types of forests in the hilly regions of southern Jiangsu Province [J].Chin J Ecol,1997,16 (6): 17-21.
[4]李惠敏,陆帆,唐仕敏,等.城市化过程中余杭市森林碳汇动态[J].复旦学报:自然科学版,2004,43(6):1044-1050.LI Huimin,LU Fan,TANG Shimin,et al.Dynamic carbon sink of forests in Yuhang City with the development of urbanization [J].J Fudan Univ Nat Sci,2004,43 (6): 1044-1050.
[5]李正才,傅懋毅,徐德应.竹林生态系统与大气二氧化碳减量[J].竹子研究汇刊,2003,22(4):1-6.LI Zhengcai,FU Maoyi,XU Deying.Bamboo ecosystem and carbon dioxide sequestration [J].J Bamboo Res,2003,22(4): 1-6.
[6]LAM N S N,COLA L De.Fractal in Geography [M].Englewood Cliffs: Prentice-Hall,1993: 3-21.
[7]MIGUEL A M,YAKOV A P,EDMUND P.Scaling,fractals and diversity in soils and ecohydrology [J].Ecol Modelling,2005,182: 217-220.
[8]任海保,张林艳,马克平.不同植物类群物种丰富度垂直格局分形特征的比较[J].植物生态学报,2005,29(6):901-909.REN Haibao,ZHANG Linyan,MA Keping.Comparison of fractal characteristics of species richness patterns between different plant taxonomic groups along an altitudinal gradient[J].Acta Phytoecol Sin,2005,29 (6): 901-909.
[9]马克明,叶万辉,桑卫国,等.北京东灵山地区植物群落多样性研究(X)不同尺度下群落样带的β多样性及分形分析[J].生态学报,1997,17 (6): 626-634.MA Keming,YE Wanhui,SANG Weiguo,et al.Study on plant community diversity in Donglingshan Mountain,Beijing,China (Ⅹ): β diversity and fractal analysis on transect on different scales [J].Acta Ecol Sin,1997,17 (6):626-634.
[10]祖元刚,马克明,张喜军.植被空间异质性的分形分析方法[J].生态学报,1997,17(3):333-337.ZU Yuangang,MA Keming,ZHANG Xijun.A fractal method for analyzing spatial heterogeneity of vegetation [J].Acta Ecol Sin,1997,17 (3): 333-337.
[11]IUDIN D I,GELASHVILI D B,ROZENBERG G S.Multifractal analysis of the species structure of biotic communities [J].Doklady Biol Sci,2003,389: 143-146.
[12]杜华强,汤孟平,周国模,等.天目山物种多样性尺度依靠及其与空间格局关系多重分形[J].生态学报,2007,27(12): 5038-5049 DU Huaqiang,TANG Mengping,ZHOU Guomo,et al.Spatial scale dependence of the species diversity in Tianmu Mountain and its relationship with spatial patterns by using multifractal analysis [J].Acta Ecol Sin,2007,27 (12):5038-5049.
[13]马克明,祖元刚.羊草种群地上部生物量与株高的分形关系[J].应用生态学报,1997,8(4):417-420.MA Keming,ZU Yuangang.Fractal relationship between aboveground biomass and plant height of Aneurolepidium chinese population [J].Chin J Appl Ecol,1997,8 (4): 417-420.
[14]李连芳,王培,王警龙,等.老芒麦(Elmus sibiricus)种群地上生物量空间分布格局研究[J].生态学报,2001,21(7): 1028-1211.LI Lianfang,WANG Pei,WANG Jinglong,et al.The spatial distribution pattern of aboveground biomass of Elymus sibiricus population [J].Acta Ecol Sin,2001,21 (7): 1028-1211.
[15]孙志高,刘景双,王金达.三江平原典型小叶章种群地上生物量的空间结构分形特征生态环境[J].生态环境,2006,15 (4): 775-780.SUN Zhigao,LIU Jingshuang,WANG Jinda.The fractal characteristics of spatial structure of aboveground biomass in typical Calamagrostis angustifolia population in Sanjiang Plain [J].Ecol Environ,2006,15 (4): 775-780.
[16]李伟成,盛海燕,潘伯荣,等.3种沙漠植物地上部分形结构与生物量的自相似性[J].林业科学,2006,42(5): 11-16.LI Weicheng,SHENG Haiyan,PAN Borong,et al.Self-similarity relationship between component of shoot and biomass of three hungriness plants [J].Sci Silv Sin,2006,42 (5): 11-16.
[17]RICOTTA C,AVENA G C.Fractal modelling of the remotely sensed two-dimensional net primary production pattern with annual cumulative AVHRR NDVI data [J].Intern J Remote Sens,1998,19 (12): 2413-2418.
[18]周国模.毛竹林生态系统中碳储量、固定及其分配与分布的研究[D].杭州:浙江大学,2006.ZHOU Guomo.Carbon Storage,Fixation and Distribution in Mao Bamboo (Phyllostachys pubescens) Stands Ecosystem [D].Hangzhou: Zhejiang University,2006.
[19]朱启疆,戎太宗,孙睿,等.林火扩展的分形模拟案例研究[J].中国科学E辑,2000,30(增刊):106-112.ZHU Qijiang,RONG Taizong,SUN Rui,et al.Study of fractal simulation case of fire spread [J].Sci China E,2000,30 (supp): 106-112.
猜你喜欢
闽南师范大学学报(自然科学版)(2022年3期)2022-12-06
湖北大学学报(自然科学版)(2022年3期)2022-12-01
天津农林科技(2022年2期)2022-04-19
延安大学学报(自然科学版)(2020年4期)2021-01-15
浙江农林大学学报(2020年5期)2020-11-05
意林·少年版(2020年13期)2020-08-02
汉语世界(2020年3期)2020-06-19
东坡赤壁诗词(2020年2期)2020-06-04
湖州职业技术学院学报(2020年4期)2020-03-30
燃气涡轮试验与研究(2010年4期)2010-04-16
