
小麦GLB1蛋白单克隆抗体的制备及鉴定*
2014-03-27 04:24孔令超杨成彬詹政科刘志强刘立忠刘志刚朱清仙
中山大学学报(自然科学版)(中英文) 2014年2期
孔令超,杨成彬,詹政科,刘志强,刘立忠,刘志刚,朱清仙
(1. 深圳大学医学院过敏反应与免疫学研究所,广东 深圳 518060;2.南昌大学研究生院医学部2011级,江西 南昌330006;3.深圳大学医学院血液循环教研室,广东 深圳 518060;4. 南昌大学医学院组胚教研室,江西 南昌330006)
I型糖尿病(T1D)又称胰岛素依赖型糖尿病(Insulin- dependent Diabetes Mellitus,IDDM),是一种器官特异性自身免疫疾病,该类病人由于机体胰岛素绝对缺乏而需终身依赖外源胰岛素来维持机体血糖的平衡。I型糖尿病主要发生在青少年,占糖尿病患者的5%~10%[1-2]。过去50年中全球I型糖尿病发病率增长了2~4倍,0-14岁儿童发病率每年以3%的速率增长[3-6]。I型糖尿病患者不仅有多尿、多食、多饮、消瘦等糖尿病的典型症状,还会继发出现糖尿病酮症酸中毒,晚期还因微血管病变导致视网膜病变及肾功能损害。由此可见I型糖尿病严重危害患者的健康和生活质量,同时也给患者、家庭、社会造成巨大的经济负担。I型糖尿病是由T淋巴细胞介导,通过对胰岛的浸润、引发胰岛炎症,破坏及降低胰岛β细胞分泌胰岛素的能力,导致胰岛素绝对缺乏[7]。绝大多数I型糖尿病是在易感基因的基础上,由一个或多个环境因素诱导引起。
研究发现食物是与I型糖尿病发病最为密切的环境因素[1]。对糖尿病易感大鼠(BB大鼠)的研究揭示,食物可诱导糖尿病的发生和发展[8-11]。有研究证明小麦麸质是最强的致糖尿病的食物因素[12-15]。以小麦为主食的NOD小鼠的糖尿病的发病率明显高于饲养不含小麦麸质的对照组小鼠[16-17],越来越多的研究提示小麦蛋白在人的I型糖尿病发生和病理过程中起重要作用[18-19]。最近的研究进一步表明小麦麸质所含的Glb 1在其致糖尿病的发生中起到了重要的作用。Scott实验室利用易感BB大鼠IgG抗体从小麦cDNA表达文库中筛选出小麦储藏蛋白(Glb 1)[20]。抗Glb 1抗体不出现在非糖尿病人的血清,但对小麦敏感的I型糖尿病患者血清检测出高滴度的抗Glb 1抗体[21],提示小麦Glb 1蛋白与I型糖尿病的发生发展密切相关。然而小麦Glb 1蛋白介导的免疫反应及其在I型糖尿病发病中的作用机理尚不清楚。因此研制小麦Glb 1蛋白对I型糖尿病的发生发展研究有重要意义。小麦储藏蛋白(Glb1)是一种球蛋白,在小麦胚乳和小麦麸质蛋白中含量丰富,研究显示糖尿病大鼠Glb1的抗体浓度明显高于非糖尿病对照组,且与胰腺的炎症和破坏程度密切相关。Glb1不仅在小麦喂养的BB大鼠具有高抗原性,而且在I型糖尿病病人体内也具有高抗原性[22]。临床资料显示对小麦高度敏感的I型糖尿病患者表现过强的针对Glb1的免疫反应[21]。现有研究表明Glb1与胰岛β细胞损害密切相关,因此研究Glb1介导的免疫反应及其对胰岛β细胞和胰岛素分泌影响的机理,对于揭示I型糖尿病的病因和发病机理,为更有效的预防和治疗I型糖尿病的提供理论依据。单克隆抗体因为其特异性强,灵敏度高且可以进行抗原表位分析的生物学特性在该研究中能发挥重要作用。本实验以小麦Glb1-G3蛋白为免疫原,利用细胞融合技术.建立了稳定分泌抗Glb-1特异性单克隆抗体的细胞株。并对得到的这些单克隆抗体的生物学特性进行鉴定。抗Glb-1单克隆抗体的成功研制,对进一步探讨Glb-1的生物学功能,阐明Glb-1与I型糖尿病的致病机制提供了有力的实验工具。现将制备该单克隆抗体的过程报道如下。
1 材料与方法
1.1 材料
1.1.1 实验动物及细胞株 5周龄雌性BALB /c小鼠,购自广东省医学实验动物中心;小鼠骨髓瘤细胞NS-1由汕头大学医学院炎症学与变态反应学实验室赠与; 小麦Glb1-G3蛋白由美国GenScript The Biology CRO公司合成。
1.1.2 试剂 DMEM 高糖培养基、8-氮鸟嘌呤(8-AG)、HAT、HT、双抗、胎牛血清(FBS)购于Gibco 公司;福氏完全佐剂、福氏 不完全 佐剂、羊抗小鼠IgG-HRP、Ig 类与亚类鉴定试剂盒、PEG SOLUTION购于Sigma 公司;链酶亲和素-HRP 购于KPL 公司;蛋白Marker购于美国Fermentas 公司。
1.1.3 抗原信息 小麦Glb1-G3抗原是选取小麦Glb1蛋白序列第595至612位多肽人工合成,序列构成为:CEGFVAGPEQQSREQEQEQER,相对分子质量2 464.55。动物免疫用抗原为偶联了Keyhole Limpet Hemocyanin(KLH) 小麦GLB1-G3多肽。均由美国GenScript The Biology CRO公司合成。
1.2 方法
1.2.1 杂交瘤细胞株的建立 取0.5 mg/mL的小麦Glb1-G3蛋白0.5 mL与等体积的福氏完全佐剂充分乳化后皮下多点注射于5只BALB/c小鼠,每只200 μL。每间隔3周改用福氏不完全佐剂加强免疫1次,剂量同前。第2次加强免疫后1周取尾血测效价。取其中免疫效果最好的小鼠融合前3 d经尾静脉注射含50 μg小麦Glb1-G3蛋白的PBS溶液200 μL再加强免疫1次。小鼠骨髓瘤细胞NS-1于融合前两周复苏培养于含有8-氮鸟嘌呤的w=10%FBS高糖DMEM培养基中,融合前一天更换为普通培养基。细胞融合时,分别收集免疫小鼠脾细胞和NS-1 细胞,不完全培养基洗涤3 次后,调整脾细胞数与NS-1细胞数之比约为1∶5,w=50% PEG1500 作用1~2 min 后,加入含有HAT的完全培养基稀释,接种于96 孔培养板,37 ℃,φ=5 % CO2培养。约2周后,用间接ELISA 法筛选阳性生长的细胞克隆孔,并更换为含有HT的完全培养基,1周后更换为完全培养基,有限稀释法进行克隆及亚克隆。
1.2.2 单克隆抗体的制备及纯化 注射0.5 mL液体石蜡至BALB/c小鼠腹腔。2周后注射1 mL 1×106的杂交瘤细胞。5~7 d后,收集腹水。室温3 000 r·min-1离心5 min吸取上清。参照 HiTrap protein A亲和层析说明书进行腹水的抗体纯化。
1.2.3 单克隆抗体效价分析 使用间接ELISA方法对各株单克隆抗体腹水效价进行分析。用浓度为1 mg/mL的小麦Glb1-G3蛋白100 μL/孔包被,4 ℃过夜,洗板3次,w=3%BSA封闭37 ℃ 2 h,小鼠腹水以1∶1 000、1∶2 000、1∶4 000、1∶8 000、1∶16 000、1∶32 000、1∶64 000、1∶128 000、1∶256 000、1∶512 000、1∶1 024 000、1∶2 048 000梯度稀释后100 μL/孔 37 ℃孵育1 h,阴性小鼠血清为阴性对照,阳性小鼠血清为阳性对照,洗板5次,加入羊抗小鼠IgG-HRP作为二抗37 ℃孵育1 h,洗板3次,TMB显色,终止后酶标仪测A450nm值。
1.2.4 单克隆抗体亲和力测定 采用间接ELISA法,分别以1∶1、1∶2、1∶4的小麦Glb 1-G3蛋白抗原 (质量浓度1、0.5、0.25 mg/mL)包被酶标板,封闭后,依次加入倍比稀释的4株单克隆抗体及羊抗小鼠IgG-HRP,加TMB显色后,测定A450nm值。以每一株单克隆抗体的浓度为横坐标,以其相应的A450nm值为纵坐标,分别绘出包被不同抗原浓度的3条反应曲线。以各曲线上部趋于平坦的A值作为100%,查出A450nm值为50%点对应的单克隆抗体的浓度[Ab]t,这样每份被检样品在包被抗原浓度分别为1∶1、1∶2、1∶4时可得[Ab]t、[Ab’]t、[Ab’’]t3个值,然后按下列公式计算K值,当包被抗原浓度为1∶2时,K=1/2(2[Ab’]t-[Ab]t)和K=1/2(2[Ab”]t-[Ab’]t)计算,当包被抗原浓度为1∶4时,K=3/2(2[Ab”]t-[Ab]t)。这样每株单克隆抗体可以得到3个K值,取3个K值的平均值即为亲和常数, 以Ka表示
1.2.5 单克隆抗体亚型测定 间接ELISA方法,以标记辣根过氧化物酶的IgG,IgG1,IgG2a,IgE为二抗测定抗体亚型。
1.2.6 八大类食物过敏原交叉反应性检测 将小麦Glb1-G3蛋白及小麦、大豆、榛子、花生、鸡蛋、虾、鱼、牛奶8种食物过敏原包被在酶标板上,分别以4株单抗为一抗,羊抗小鼠IgG-HRP为二抗,免疫前小鼠血清为阴性对照,免疫后小鼠血清为阳性对照,进行间接ELISA检测,显色后A450nm值。
1.2.7 双抗夹心ELISA法检测小麦Glb1-G3蛋白 利用ELISA方法对四种单抗进行两两配对实验。将4种单抗包被酶标板,加入梯度稀释的小麦Glb1-G3蛋白作为抗原37 ℃孵育1 h,洗板5次,分别加入用生物素标记的4种单抗两两配对,37 ℃孵育1 h,洗板3次,加入链酶亲和素-HRP,TMB显色,终止后酶标仪测A450 nm值。确定包被抗体与标记抗体的种类及稀释度。用建立起来的双抗夹心ELISA系统检测小麦Glb1-G3蛋白。
2 实验结果
2.1 杂交瘤细胞株的建立
细胞融合率为 96.2% (452/470),抗体阳性检出率为 87.4% (411/470),选择其中较高读数的 10 株进行克隆和亚克隆,最终获得4株分泌抗小麦Glb1-G3蛋白单克隆抗体的杂交瘤细胞株,分别命名为:1C4、4H5、1A9、4F5。
2.2 单克隆抗体的制备及纯化
将收集好的各单抗腹水参照 HiTrapprotein A亲和层析说明书进行纯化。纯化后的抗体经过w=12%SDS-PAGE分析,可见清晰地2条蛋白条带,位于55 000附近处的为抗体重链,位于26 000附近的为轻链(图1)。
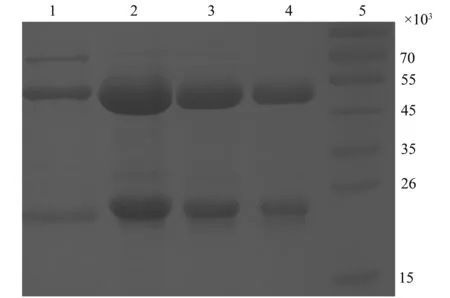
图1 小麦Glb1-G3蛋白单克隆抗体纯度鉴定Fig.1 Identification of the purity of McAbs against Glb1-G31:1C4; 2:4H5; 3: 4F5; 4: 1A9; 5: Protein Marker
2.3 单克隆抗体效价分析
将取得的小鼠腹水按一系列梯度稀释,用间接 ELISA 法测定单抗的效价,4株抗体效价均超过200万(图2)。

图2 单克隆抗体效价Fig.2 Detection of the potency of McAbs against Glb1-G3 allergen
2.4 单克隆抗体亲和力测定




图3 单克隆抗体的亲和力检测Fig.3 Aviclity of the monoclonal antibodies
分别以1∶1、1∶2、1∶4的小麦Glb1-G3蛋白抗原 (质量浓度1、0.5、0.25 mg/mL)包被酶标板,间接ELISA方法测定封闭后,依次加入倍比稀释的4株单克隆抗体及羊抗小鼠IgG-HRP,加TMB显色后,测定A450nm值。以每一株单克隆抗体的浓度为横坐标,以其相应的A450nm值为纵坐标,分别绘出包被不同抗原浓度的3条反应曲线(图3-6)。以抗体1C4为例,小麦Glb1-G3蛋白抗原包被质量浓度为1、0.5、0.25 mg/mL 时各曲线上部趋于平坦的A450nm值作为100%分别为3.802、3.88、3.92,则查出50%A450nm值1.901、1.94、1.96对应的单抗浓度分别是[Ab]t=2.83×10-11mol/L、[Ab’]t=1.96×10-10mol/L、[Ab’’]t=4.55×10-10mol/L三个值,然后按下列公式计算K值,当包被抗原浓度为1∶2时,K=1/2 (2 [Ab’]t-[Ab]t)=1.37×109L/mol和K=1/2 (2 [Ab”]t-[Ab’]t)=7.00×108L/mol计算[23],当包被抗原浓度为1∶4时,K=3/2 (2 [Ab”]t-[Ab]t)=1.71×109L/mol。3个K值的平均值即为单抗1C4亲和力常数Ka=1.26×109L/mol。同理,4H5、4F5、1A9的亲和力常数分别为:7.24×109,4.79×109,4.59×109L/mol。
2.5 单克隆抗体亚型测定
用间接ELISA方法,以各单克隆抗体为一抗,标记辣根过氧化物酶的IgG,IgG1,IgG2a,IgE为二抗测定抗体亚型。检测结果4株单抗皆为IgG1亚型。

图4 单克隆抗体的亚型检测Fig.4 Subtypes of the monoclonal antibodies
2.6 八大类食物过敏原交叉反应性检测
将小麦Glb1-G3蛋白及小麦、大豆、榛子、花生、鸡蛋、虾、鱼、牛奶8种食物过敏原包被酶标板,分别以4株单抗为一抗进行间接ELISA检测,结果显示:所制备的1C4、4H5、1A9、4F5 4株单克隆抗体与小麦Glb1-G3蛋白有很好的反应能力,与大豆、榛子、花生、鸡蛋、虾、鱼、牛奶7种食物过敏原无交叉反应,4H5、4F5与小麦粗提蛋白可反应,1C4、1A9与小麦粗提蛋白反应能力较弱,可能与粗提蛋白中相关抗原含量低或者抗原结构有关。

图5 单克隆抗体与常见食物过敏原的交叉反应Fig.5 Cross-reaction of the monoclonal antibodies with familiar food allergen
2.7 双抗夹心ELISA法检测小麦Glb1-G3蛋白
利用ELISA方法对四种单抗进行两两配对实验,发现1C4与4H5可能为不同的抗原表位。棋盘法测定单抗1C4以1∶8 000稀释度包被,生物素标记的4H5以1∶32 000稀释度反应效果较好。小麦Glb1-G3蛋白做倍比稀释,阴性对照孔用PBS代替抗原,以测定孔与阴性孔A450nm值之比大于2.1为阳性来检测该方法对抗原的检出范围。该方法对小麦Glb1-G3蛋白的检出限为3.75 ng/mL,标准曲线在3.75~120 ng/mL范围内线性良好。

图6 单克隆抗体对小麦蛋白Glb1-G3的检测Fig.6 Detection of the wheat protein Glb1-G3 with the monoclonal antibodies
3 讨 论
小麦经过长期的发展,已经成为世界上分布最广、面积最大、总产量第二、贸易额最多、营养价值最高的粮食作物之一。全世界有43个国家,有35%~40%的人口以小麦为主要粮食。小麦子粒含有丰富的淀粉、较多的蛋白质、少量的脂肪,还有多种矿物质元素和维生素B,是一种营养丰富、经济价值较高的商品粮。据联合国粮农组织2004年统计,全世界小麦收获面积32.36 亿亩(1亩约为667 m2),单产193.8 kg/亩,总产6.27 亿t ,单产较高的国家主要集中在西欧。但是于此同时,小麦也是FAO 1995年所报道的八大类常见过敏食物[24]。糖尿病(Diabetes Mellitus)是一组与胰岛素产生和作用异常相关、以血糖增高为特征的慢性疾病,该病及其并发症具有危害大、治愈难、费用高等特点,已成为严重威胁人类健康的社会问题。糖尿病的发病原因众多,从营养学角度来说,进食个体不耐受的食物以及紊乱的饮食结构是糖尿病发病的一个重要因素[25]。小麦及其相关产品在日常饮食中的广泛摄入,使小麦成为主要的食物蛋白来源,但同时也成为主要的食物过敏原。小麦过敏可以诱发哮喘、鼻炎、接触性荨麻疹、乳糜泻肠炎等症状[26]。小麦储藏蛋白作为小麦的主要过敏原,本研究是根据小麦储藏蛋白在小麦过敏中的重要作用利用杂交瘤技术制备抗Glb-1特异性单克隆抗体,并鉴定其生物学特性,为研究Glb-1与I型糖尿病的关系提供了研究基础。
根据软件分析Glb-1蛋白的结构,选择性的选取了两段亲水、暴露、无转角且抗原性强的肽段表达分别命名为G1、G2。经免疫和细胞融合后证实G1、G2免疫BALB/C小鼠后ELISA检测杂交瘤细胞抗体检出阳性率较低。而用于免疫小鼠的Glb-1 G3肽段是根据文献报道将2段与糖尿病相关的短肽人工合成表达,G3序列构成为:CEGFVAGPEQQSREQEQEQER,相对分子质量2 464.55,实验结果G3免疫后的BALB/C小鼠脾细胞与NS-1细胞融合后检测融合率和抗体阳性率均较理想。 本实验利用细胞融合技术制备了小鼠抗Glb-1的杂交瘤细胞,细胞融合率较高且用Glb-1 G3包被ELISA检测出的抗阳性检出率理想。经过对ELISAA值读数较高的10株进行克隆和亚克隆,最终获得4株稳定分泌抗小麦Glb1-G3蛋白单克隆抗体的杂交瘤细胞株,分别命名为:1C4、4H5、1A9、4F5。 通过向小鼠注射这4株杂交瘤细胞后收集腹水通过亲和层析进行纯化。将小鼠腹水梯度稀释后用间接ELISA法测定单抗的效价,4株抗体效价均超过200万。且经检测后4株单抗皆为IgG1亚型。4株单抗为一抗分别与八大类食物进行交叉反应性检测结果表明该4株单抗均能特异性识别小麦Glb-1蛋白且与其他主要食物均无交叉反应,特异性强. 双抗夹心ELISA法检测小麦Glb1-G3蛋白实验发现1C4与4H5可能为不同的抗原表位,可以准确检测Glb-1蛋白,可以制备小麦过敏原检测试剂盒,具有实际应用价值。
综上所述,本研究已获得鼠抗小麦Glb-1单克隆抗体,为研究小麦过敏与临床疾病的关系和小麦主要过敏原蛋白的蛋白的检测了研究工具和研究依据,同时也为小麦Glb-1蛋白的结构和生物学功能研究奠定了基础。
[1] LEFEBVRE D E,POWELL K L, STROM A, et al. Dietary proteins as environmental modifiers of type 1 diabetesmellitus[J]. Annu Rev Nutr,2006,26:175-202.
[2] PATRICK C, WANG G S, LEFEBVRE D E, et al. Promotion of autoimmune diabetes by cereal diet in the presence or absence of microbes associated with gut immune activation, regulatory imbalance, and altered cathelicidin antimicrobial peptide[J].Diabetes,2013, 62(6): 2036-2047.
[3] ONKAMO P, VAANANEN S, KARVONEN M,et al. Worldwide increase in incidence of type I diabetes—the analysis of the data on published incidence trends[J].Diabetologia,1999;42:1395-1403.
[4] GALE E A.The rise of childhood type 1 diabetes in the twentieth centry[J].Diabetes,2002,51:3335-3361.
[5] GILLESPIE K M, BAIN S C, BARNETT A H, et al. The rising incidence of childhood type 1 diabetes and reduced contribution of highrisk HLA haplotypes[J]. Lancet, 2004,364:1645-1647.
[6] LIESE A D,MA X, MAAHS D M, et al. Physical activity, sedentary behaviors, physical fitness, and their relation to health outcomes in youth with type 1 and type 2 diabetes: A review of the epidemiologic literature[J]. Journal of Sport and Health Science, 2013,2(1): 21-38.
[7] 许曼音,陆广华,陈名道.糖尿病学[M].2版.上海:上海科学技术出版社, 2010:87-92.
[8] ELLIOTT R B, MARTIN J M. Dietary protein: a trigger of insulin-dependent diabetes in the BB rat?[J]. Diabetologia,1984,26:297-299.
[9] ISSA-CHERGUI B,GUTTMANN R D,SEEMAYER T A,et al. The effect of diet on the spontaneous insulin dependent diabetic syndrome in the rat[J]. Diabetes Res, 1988,9:81-86.
[10] VISSER J, BRUGMAN S, KLATTER F, et al. Short-term dietary adjustment with a hydrolyzed casein-based diet postpones diabetes development in the diabetes-prone BB rat[J]. Metabolism-Clinical and Experimental,2003,52(3): 333-337.
[11] MAURANO F,ROSSI M, TRONCONE R. Gut involvement in NOD mouse diabetes[J]. Immunology Endocrine & Metabolic Agents in Medicinal Chemistry,2007,7(3): 211-217.
[12] GRAHAM S, COURTOIS P, MALAISSE W J, et al. Enteropathy precedes type 1 diabetes in the BB rat[J]. Gut, 2004,53:1437-1444.
[13] VAARALA O, ATKINSON M A, NEU J. The “perfect storm” for type 1 diabetesthe complex interplay between intestinal microbiota, gut permeability, and mucosal immunity[J]. Diabetes, 2008, 57(10):2555-2562.
[14] MACFARLANE A J,BURGHARDT K M, KELLY J,et al. A type 1 diabetes-related protein from wheat (Triticumaestivum)cDNA clone of a wheat storage globulin, glb1, linked to islet damage[J]. The Journal of Biological Chemistry, 2003,278: 54-63.
[15] SCOTT F W,SARWAR G,CLOUTIER H E. Diabetogenicity of various protein sources in the diet of the diabetes-prone BB rat[J]. Advances in Experimental Medicine and Biology, 1989, 246: 277-285.
[16] COLEMAN D L, KUZAVA J E, LEITER E H. Effect of diet on incidence of diabetes in nonobese diabeticmice[J]. Diabetes,1990,39:432-436.
[17] ANTVORSKOV J C, FUNDOVA P, BUSCHARD K,et al. Dietary gluten alters the balance of pro-inflammatory and anti-inflammatory cytokines in T cells of BALB/c mice[J]. Immunology , 2013,138(1):23-33.
[18] ZIEGLER A G, SCHMID S, HUBER D, et al.Early infant feeding and risk of developing type 1 diabetes-associated autoantibodies[J]. JAMA,2003,290:1721-1728.
[19] NORRIS J M, BARRIGA K, KLINGENSMITH G, et al. Timing of initial cereal exposure in infancy and risk of islet autoimmunity [J]. JAMA, 2003, 290:1713-1720.
[20] SCOTT F W, MONGEAU R, KARDISH M, et al. Diet can prevent diabetes in the BB rat[J].Diabetes,1985,34(10): 1059-1062.
[21] MOJIBIAN M,CHAKIR H,MACFARLANE A J, et al.Immune reactivity to a Glb 1 homologue in a highly wheat-sensitive patient with type 1 diabetes and celiac disease [J].Diabetes Care, 2006, 29(5):1108-1110.
[22] SIMPSON M, MOJIBIAN M, BARRIGA K,et al. An exploration of Glo-3A antibody levels in children at increased risk for type 1 diabetes mellitus[J].Pediatric Diabetes, 2009,10(8): 563-572.
[23] 柳忠辉. 医学免疫学实验技术[M]. 北京: 人民卫生出版社, 2008: 22-23.
[24] Food and Agriculture Organization of the United Nations Report of the FAO technical consultation on food allergies[C].Rome, Italy:Food and Agriculture Organization of the United Nations, 1995.
[25] 钱荣立.饮食紊乱与糖尿病[J].中国糖尿病杂志, 2012,20(1):5-6.
[26] JOHANSSON S, HOURIHANE J, BOUSQUET J, et al. A revised nomenclature for allergy: An EAACI position statement from the EAACI nomenclature task force [J]. Allergy,2001,56(9): 813-824.
猜你喜欢
昆明医科大学学报(2021年2期)2021-03-29
昆明医科大学学报(2021年1期)2021-02-07
中国现代医药杂志(2020年10期)2020-12-14
保健与生活(2019年12期)2019-07-31
医药前沿(2019年7期)2019-01-05
上海故事(2015年13期)2016-01-22
肿瘤影像学(2015年3期)2015-12-09
伴侣(2015年5期)2015-09-10
癌变·畸变·突变(2015年3期)2015-02-27
湖南饲料(2014年4期)2014-08-26
