
两步融合PCR法构建烟曲霉ssk1基因敲除株
2014-05-21 03:51冯文莉山西医科大学第二临床医学院皮肤科太原030001通讯作者mailmayan197522163com
山西医科大学学报 2014年4期
马 彦,岑 雯,刘 昕,冯文莉(山西医科大学第二临床医学院皮肤科,太原 030001;通讯作者,E-mail:mayan197522@163.com)
近年来随着新技术的开展和免疫抑制剂的应用,侵袭性曲霉病的发病率在临床呈现明显上升趋势且病死率高,而烟曲霉(Aspergillus fumigates,A.fumigatus)是其主要致病菌,因此探讨烟曲霉感染的发病机制变得刻不容缓[1]。通过构建基因敲除株对目的基因进行研究是目前常用来研究烟曲霉的一种方法,既往的实验技术多数通过克隆技术将指定片段的基因进行敲除,往往受到所选基因序列和载体酶切位点的限制,有时不能完整地敲除整个基因或者指定片段,且耗时较长,本研究通过使用融合PCR法成功构建烟曲霉ssk1基因敲除株,节省时间,大大简化了烟曲霉基因敲除载体构建的过程,现具体说明如下。
1 材料和方法
1.1 菌株和质粒
烟曲霉ku80作为野生对照菌株,烟曲霉转化用宿主菌为A.fumigatus ku80 pyrG-,该菌株为嘧啶营养缺陷株,在没有尿苷和尿嘧啶的培养基上不能生长。筛选标记pyrG来源于A.parasiticus(由杜克大学William J.Steinbach教授赠送)。
1.2 培养基
GMM基础培养基(1 L):20×盐溶液50 ml,微量元素液 1 ml,D - 葡萄糖10 g,琼脂15 g(1.5%),pH 值 6.5。
20 × 盐溶液(1 L):NaNO3120 g,KCl 10.4 g,MgSO4·7H20 10.4 g,KH2PO430.4 g,加水至 1 L,室温储存。
微量元素液(1 L):ZnSO4·7H20 2.2 g,H3BO31.1 g,MnCl2·4H2O 0.5 g,FeSO4·7H2O 0.5 g,CoCl2·5H2O 0.16 g,CuSO4·5H2O 0.16 g,(NH4)6Mo7O24·4H2O 0.11 g,Na4EDTA 5.0 g。
溶壁酶:Vinoflow FCE(Novozymes公司);等渗培养基:1.2 mol/L MgSO4,10 mmol/L sodium phosphate,4 ℃保存;Trapping 缓冲液:0.6 mol/L sorbitol,0.1 mol/L Tris-HCl,4 ℃保存;STC 溶液:1.2 mol/L sorbitol,10 mmol/L CaCl2,10 mmol/L Tris - HCl pH 7.5,在4℃保存;聚乙二醇氯化钙溶液:60%PEG,50 mmol/L CaCl2,50 mmol/L Tris-HCl pH 7.5在室温保存;1.5%底层琼脂培养基(SMM)∶20×盐溶液50 ml,微量元素液1 ml,葡萄糖10 g,琼脂15 g,山梨醇218.6 g,酵母提取物 1 g,pH 值6.5,加水至 1 L,高压灭菌20 min。0.7%顶层琼脂培养基:配方同上(1.5%),将其中琼脂改为7 g/L。
1.3 生物信息学分析
在烟曲霉基因组中(www.aspergillusgenome.org)找出与念珠菌ssk1基因同源的基因,并利用SMART软件对此基因功能区进行初步分析。
1.4 烟曲霉ssk1基因敲除的构建
1.4.1 基因敲除的构建 烟曲霉ssk1基因敲除构建示意图见图1。
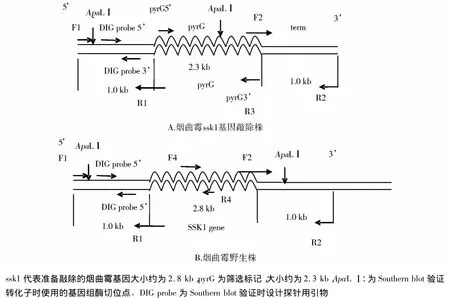
图1 ssk1基因敲除构建示意图Figure 1 Sketch map of ssk1gene deletion
1.4.2 引物 本实验中所用引物见表1。
1.4.3 基因片段的扩增与融合 本实验采用了两轮PCR反应,其体系及反应条件如下。
第一轮PCR:以野生株基因组DNA为模板,首先分别使用F1、R1;F2、R2扩增相应的片段。以A.parasiticus DNA为模板使用引物pyrG-5’,pyrG-3’扩增筛选标记pyrG。具体PCR扩增体系及反应条件如下(试剂盒为NEB公司高保真融合PCR DNA 聚合酶):无核酸酶的水31.5 μl,5 × 缓冲液10 μl,10 mmol/L dNTPs 1 μl,10 μmol/L 上游引物 2.5 μl,10 μmol/L 下游引物2.5 μl,模板 DNA(浓度100 ng/μl)2 μl,高保真融合 PCR DNA 聚合酶 0.5 μl,合计 50 μl。
反应条件:98 ℃ 5 min,98 ℃ 10 s,60 ℃30 s,72℃1 min,共30个循环。72℃ 10 min,4℃保存。
该反应中F1、R1,F2、R2扩增相应序列片段均为1.0 kb。因为扩增条带大小相等,且退火温度差别不大,故使用相同的扩增条件,产物标记为P1,P2。扩增筛选标记 pyrG:使用引物 pyrG-5’和pyrG -3’,产物大小约为 2.3 kb,体系同前,PCR 反应条件中延伸时间改为2 min,产物标记为P3。

表1 本研究中所使用的引物序列Table 1 Sequences of the primer in this study
0.8 %琼脂糖凝胶电泳,确定片段大小,并切胶纯化。纯化用试剂盒为:Fermentas公司的Gene JET gel extraction kit。
第二轮PCR反应:也是最重要的一步反应即融合基因的扩增。其反应体系及反应条件如下:无核酸酶的水 30.5 μl,5 × 缓冲液 10 μl,10 mmol/L dNTPs 1 μl,10 μmol/L 上游引物 2.5 μl,10 μmol/L下游引物 2.5 μl,模板 DNA 3 μl,高保真 DNA 聚合酶 0.5 μl,体系合计 50 μl。其中模板 DNA 为 P1,P2,P3三种产物各取1 μl的混合体,将延伸条件改为4 min进行扩增。其反应条件如下:98℃ 10 s,60℃ 30 s,72 ℃ 4 min共1个循环,98 ℃ 10 s,62 ℃30 s,72℃ 4 min,共29个循环。72 ℃ 10 min,4 ℃保存。
0.8 %琼脂糖凝胶电泳,对比Marker确定正确大小的融合基因片段,切胶纯化,并定量(试剂同前)。
1.4.4 原生质体法构建基因敲除株 250 ml基础培养基+0.5% 酵母提取物+尿苷尿嘧啶(Uracil 0.14 g,Uridine 0.315 g)接种 Δku80 pyrG-菌悬液 2 ml,30 ℃,250 r/min摇床过夜培养(16 -20 h)。收集菌丝,使用Vinoflow FCE称量3 g,放于干净的40 ml等渗培养基中,充分吹打混匀。28℃,75 r/min摇床培养3-4 h。加入Trapping缓冲液4℃,3 500 r/min离心10 min;吸出离心管夹层的原生质体,加3倍体积的STC溶液到原生质体中,上下轻轻颠倒混匀,离心。吸出原生质体层,用1 ml的STC溶液重悬原生质体,观察原生质体状态及浓度,适当调整浓度。取5 μg的融合PCR产物加入到200 μl的原生质体中,冰上静置1 h。将200 μl DNA-原生质体放入到1.25 ml的聚乙二醇氯化钙溶液中,轻弹混匀,室温放置20 min,加入3 ml STC,轻轻混匀,取300 μl与10 ml SMM顶层培养基混合后铺板于1.5%的底层培养基上,放置于37℃孵箱中培养。具体实验方法见文献[2]。
1.5 转化子的验证
1.5.1 PCR验证 在GMM液体培养基中过夜培养单克隆转化子,并提取基因组DNA。分别使用四组引物进行扩增(F1,R4;F4,R2;F1,pyrG -3’;R2,pyrG -5’)。
扩增体系及反应条件如下:无核酸酶的水7 ul,2 × 缓冲液 10 μl,10 μmol/L 上游引物 0.5 μl,10 μmol/L 下游引物 0.5 μl,模板 DNA(浓度 100 ng/μl)2 μl,合计 20 μl。反应条件:95℃ 5 min,95 ℃10 s,60 ℃ 30 s,72 ℃ 1 min,共30个循环。72 ℃延伸10 min,4℃保存。其中延伸温度根据扩增片段大小适当调整。所用试剂为Thermo scientific Dream taq Green PCR Master mix(2×)。
1.5.2 Southern Blot验证转化子 提取转化子基因组DNA,使用ApaLⅠ过夜酶切,1.5% 琼脂糖凝胶电泳,具体实验步骤按照试剂盒说明。所用试剂Roche公司的地高辛标记SouthernBlot试剂盒。
2 结果
2.1 烟曲霉ssk1基因生物学分析
通过同源基因的序列比对我们在烟曲霉基因组中找到其同源基因为Afu5g08390,位于5号染色体,编码850个氨基酸,Smart软件分析发现在560-718氨基酸间存在信号传导受体区。序列同源性比较结果:黄曲霉相似性81%,土曲霉相似性73%,黑曲霉73%,构巢曲霉72%,白念珠菌71%,裂殖酵母75%。
2.2 PCR 扩增结果
第一步PCR扩增结果见图2,可见分别获得了大约1.0 kb的ssk1基因的上下游侧翼序列。图3可见获得了大小约为2.3 kb的片段,为筛选标记pyrG。第二步融合PCR扩增结果可见有数条高亮条带(见图4),其中大约为4.3 kb的片段为我们的目的片段。
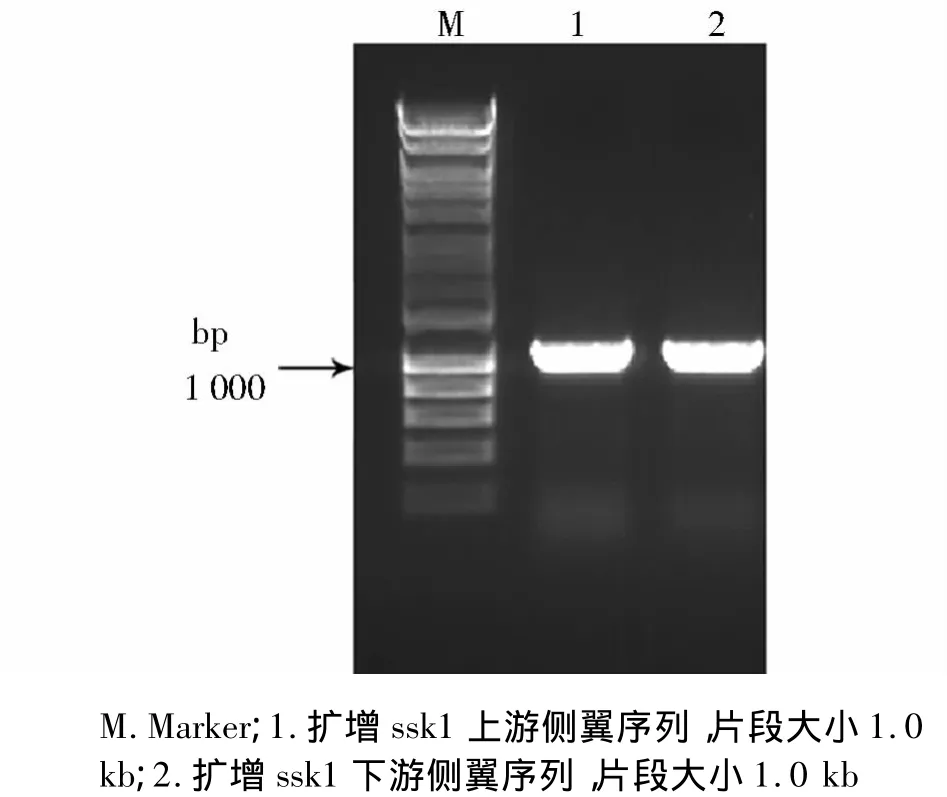
图2 ssk1基因上下游侧翼序列PCR扩增结果Figure 2 PCR amplification results of the flanking sequence of the ssk1 gene
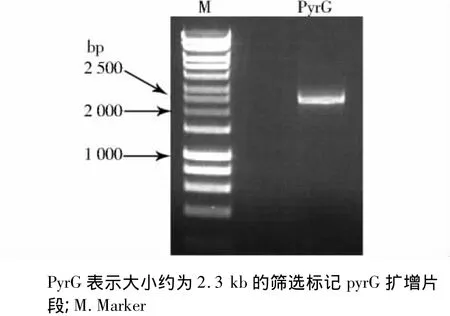
图3 筛选标记pyrG基因扩增结果Figure 3 PCR amplification results of the pyrG gene using the primer pyrG -5’and pyrG -3’
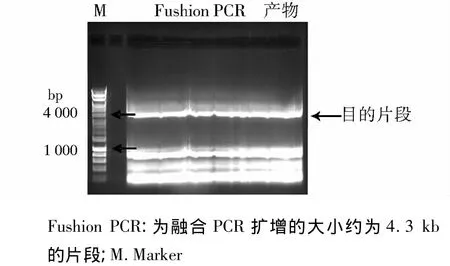
图4 融合基因PCR扩增结果Figure 4 PCR amplification results of the fushion PCR using the primer F1 and R2
2.3 转化子PCR鉴定结果
原生质体法转化烟曲霉后挑取单克隆的转化子在GMM液体培养基中进行培养,提取DNA鉴定。我们共使用四组引物进行扩增验证。
第Ⅰ组使用引物F1、R4进行扩增:由于R4引物设计在基因上,因此在野生株应该出现条带约为2 494 bp,在基因敲除株无条带出现。菌株1,3,4,5无条带出现为正确的基因敲除株,而转化子2和野生株有条带(见图5),说明转化子2不是正确的基因敲除株。
第Ⅱ组使用F4、R2进行扩增分别扩增转化子。由于F4引物设计在基因上,因此在野生株可以出现条带约为2 332 bp,在基因敲除株无条带出现。由图5可见转化子1,3,4,5无条带出现为正确的基因敲除株,而转化子2和野生株有条带,说明转化子2不是正确的基因敲除株。
第Ⅲ组使用F1、pyrG-3’这对引物进行扩增。由于pyrG-3’在筛选标记上,因此在野生株没有条带出现,在基因突变株有条带大约1.0 kb,由图5可见在野生株上无条带出现,而在转化子 1,2,3,4,5都有正确大小的条带出现,说明均有筛选标记进入到转化子中。
第Ⅳ组使用pyrG-5’、R2这对引物进行扩增。由于pyrG-5’在筛选标记上,因此在野生株没有条带出现,在基因突变株有条带大约3.5 kb的条带。由图6可见在转化子1,2,3,4,5都有正确条带出现说明均有筛选标记进入到转化子中。
综上可见1,3,4,5这四个转化子为同源转化子,而转化子2为异源转化子,也就是没有结合到正确的位置上。

图5 野生株与转化子的PCR结果Figure 5 PCR results of wild strain and transformants
2.4 Southern blot验证结果
根据PCR的结果我们进一步验证了转化子1,3,4,5的正确性。从基因敲除构建的示意图我们可以看出在野生株使用限制性内切酶ApaLⅠ酶切后得到大约4 806 bp大小的片段,而在基因敲除株则得到大约2 110 bp大小的片段(见图7),进一步说明得到的转化子1,3,4,5是正确的转化子。
3 讨论
由于烟曲霉致病引起的侵袭性曲霉病发病明显增加,致力于烟曲霉研究的学者也越来越多,而目的基因的特异性敲除是一种常用的研究方法。既往的研究显示多数学者采取了质粒克隆的方法[2],为了提高烟曲霉转化的效率,近年来使用根癌农杆菌介导的烟曲霉的转化方法有效地提高了转化效率[3,4]。然而构建敲除载体的过程仍然复杂,需要精心选择酶切位点并构建多个中间载体,耗时较长,且受到载体种类和酶切位点的限制,往往造成有些基因序列不能顺利构建载体,延长了研究周期。
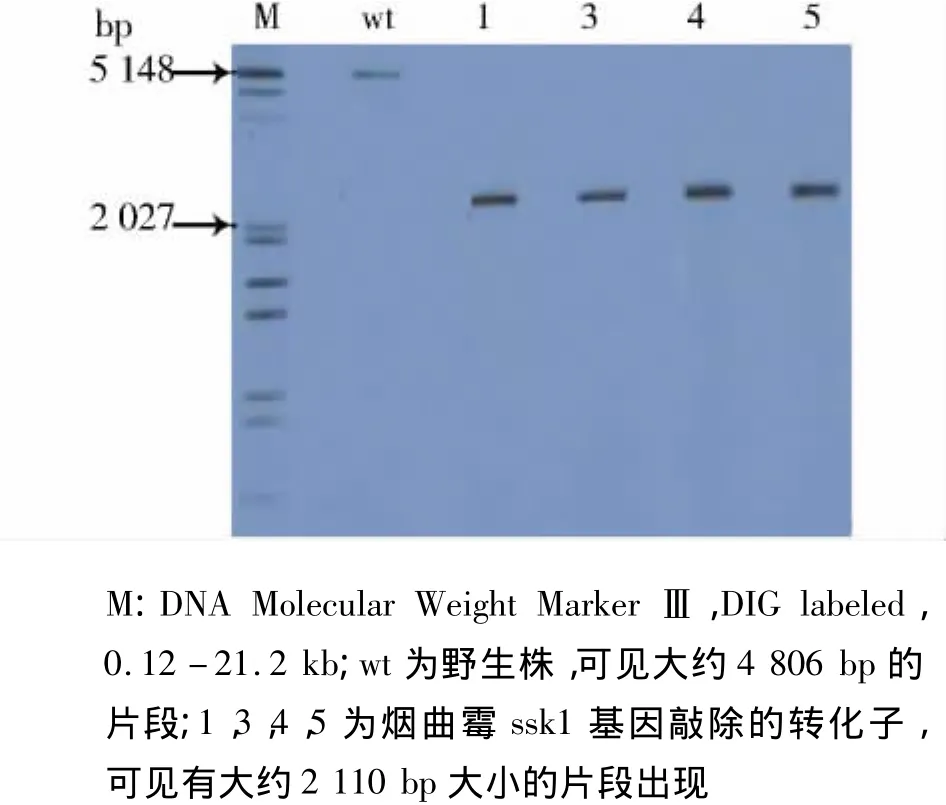
图7 使用Southern blot对转化子进行验证Figure 7 The verification of the transformants using Southern blot
近年来很多学者尝试改变构建载体的方法,以更简洁、更方便的方法来替代传统的方法。融合基因法陆续有人报道[5,6]。融合 PCR(fusion PCR)是将任意两条或三条具有同源区域的片段,通过PCR的方法实现连接的技术方法,融合PCR作为一种常用的构建重组片段或重组载体的手段常用于基因功能研究,而随着越来越多的基因组被测序出来,对目的基因进行多种功能分析成为可能,如目的基因特异性敲除、目的基因过量表达或体外表达,或者对目的基因插入荧光标记等。丝状致病真菌烟曲霉基因敲除本身有一定的难度,有研究报道在双相真菌马尔尼菲青霉的载体构建中有通过使用三步融合PCR法构建成功的报道[7]。烟曲霉基因敲除载体的构建近期也有人报道使用三步融合PCR的方法[8,9],本实验中,不同于以往报道的三步融合基因法,采用两步融合法。
首先,在引物设计时只是在特异基因的上下游侧翼序列同要融合的基因即筛选标记相邻片段的引物上设计了15个碱基的互补序列(见表1中的红色标记序列),并没有在筛选标记pyrG的上下游引物上设计与要融合的上下游侧翼序列相邻片段的互补序列,这样设计的好处是筛选标记的引物可以用途更广,在构建以pyrG为筛选标记的其他基因敲除载体时都可以使用该对引物扩增筛选标记,更具有通用性,也节约了实验成本和时间,只需要针对特异的基因要融合的片段设计特异的引物序列即可。
其次,第二步融合PCR反应中将不同来源的基因模板融合形成全长的过程和扩增合并在一起进行。有文献报道先在不添加引物的情况下使用DNA聚合酶进行片段的互补延伸产生中间产物,然后用中间产物作为模板再进行全长序列的扩增[10];还有文献则先将上下游序列同筛选标记融合产生中间产物再通过一次巢氏PCR的方法获得全长融合基因[9]。我们直接将三种片段产物进行融合扩增,减少了一次PCR的反应过程,缩短了实验时间。在这里我们使用的三种模板的体积比为1∶1∶1,融合过程中各片段在体系中的浓度不同的学者有不同的见解。韩改革等认为各片段在反应体系中的终浓度≥15 ng/μl且摩尔比为 1∶1∶1 时较易获得产物[8],而刘增然等认为上下游产物以及筛选标记产物的比例为1∶1∶3也能获得较好的效果[9]。我们的经验是体积比为1∶1∶1,在其他基因敲除的过程中也取得了较好的效果。不同于上述两位作者,他们都采用了三步法进行融合PCR扩增,而我们的试验中只使用了两步也能成功获得融合的目的片段。在这个PCR反应条件中我们的第1个循环就是将模板相互融合的过程。这样做减少了实验步骤,只要通过两次PCR实验即可扩增出需要的融合片段,纯化后,进行烟曲霉转化,不同于克隆质粒,需要进行培养,大大节约了时间。我们进行两步PCR融合基因法构建烟曲霉ssk1的基因突变株发现融合PCR的特异性可能会差一些,因此出现的条带较多,合理调整PCR的反应条件是比较重要的。如果扩增的目的片段偏弱可以适当降低第1个循环中的退火温度,在第2-29个循环中退火温度适当提高可以增加产物的特异性,当然不同的基因和引物可能需要的条件不同。如果条带不清晰则需要重新调整反应条件,进一步优化。不同的实验室根据自己的经验可能有不同的方案。
总之,融合PCR这种技术在构建烟曲霉基因敲除株的好处在于其不受序列本身和载体酶切位点的限制,在没有合适的载体或者酶切位点时进行基因敲除,融合PCR的方法不失为烟曲霉基因敲除的一个很好的策略。我们通过使用融合PCR技术得到了烟曲霉ssk1基因的全基因敲除株,这为我们的下一步的实验提供了良好的平台。
[1] Delattin N,Cammue BP,Thevissen K.Reactive oxygen speciesinducing antifungal agents and their activity against fungal biofilms[J].Future Med Chem,2014,6(1):77 -90.
[2] Steinbach WJ,Cramer RA Jr,Perfect BZ.Calcineurin controls growth,morphology,and pathogenicity in Aspergillus fumigatus[J].Eukaryot Cell,2006,5(7):1091 -1103.
[3] Ma Yan,Qiao Jianjun,Liu Wei,et al.The Sho1 sensor regulates growth,morphology,and oxidant adaptation in Aspergillus fumigatus but is not essential for development of invasive pulmonary Aspergillosis[J].Infect Immun,2008,76(4):1695 - 1701.
[4] 乔建军,刘伟,马彦,等.根癌农杆菌介导的尿嘧啶缺陷烟曲霉转化是基因敲除的有效方法[J].北京大学学报,2008,40(3):330-333.
[5] 王力华,付士华,唐青,等.三重融合PCR法构建感染性新德毕斯嵌合病毒cDNA克隆[J].病毒学报,2006,22(2):107-113.
[6] 邓玲玲,李江姣,尉妍,等.含有增强型绿色荧光蛋白报告基因的新德毕斯病毒的制备与鉴定[J].中华实验和临床病毒学杂志,2013,27(3):228 -230.
[7] 王琳,蓝秀万.融合PCR方法构建马尔尼菲青霉菌LIG4基因敲除载体[J].医药前沿,2013,(11):99 -100.
[8] 韩改革,贾晓东,韩雪琳.长片段融合 PCR在构建烟曲霉rho1基因回补株中的应用[J].中国真菌学杂志,2013,8(3):129-133.
[9] 刘增然,张光一,YU Jae-Hyuk.致病菌烟曲霉新基因Afu4g13170生孢致毒相关性初步研究[J].微生物学通报,2012,39(1):68 -74.
[10] 李敏,杨谦.一组高效构建同源重组DNA片段的方法—融合PCR[J].中国生物工程杂志,2007,27(8):53 -58.
猜你喜欢
广东药科大学学报(2022年3期)2023-01-04
生物学通报(2022年1期)2022-11-22
华人时刊(2022年9期)2022-09-06
华人时刊(2020年15期)2020-12-14
山西地震(2019年1期)2019-03-20
罕少疾病杂志(2017年2期)2017-02-23
新农业(2016年23期)2016-08-16
系统工程与电子技术(2016年2期)2016-04-16
广西林业科学(2016年3期)2016-03-16
广州大学学报(自然科学版)(2015年4期)2015-12-23
