
籼稻R996转Pi9基因纯系的筛选及稻瘟病抗性鉴定
2014-08-31 07:25潘素君李魏高佳戴良英
湖南农业大学学报(自然科学版) 2014年6期
潘素君,李魏,,高佳,戴良英, *
(湖南农业大学 a.作物基因工程湖南省重点实验室;b.植物保护学院,湖南 长沙 410128)
籼稻R996转Pi9基因纯系的筛选及稻瘟病抗性鉴定
潘素君a,李魏a,b,高佳b,戴良英a,b *
(湖南农业大学 a.作物基因工程湖南省重点实验室;b.植物保护学院,湖南 长沙 410128)
采用农杆菌介导法将抗稻瘟病Pi9基因导入籼稻品种R996,获得转基因植株,并对这些转基因植株进行遗传分析。结果表明:Pi9基因已经整合到受体细胞基因组中,并在转录水平上得到了有效表达;对转基因植株后代进行潮霉素抗性、GUS染色和PCR检测分析,鉴定出Pi9基因阳性转化子;经稻瘟病抗性鉴定,从T1代符合孟德尔分离比(1∶2∶1)的分离群体中,成功筛选出转Pi9基因的籼稻R996纯系。
籼稻R996;Pi9基因;转基因植株;稻瘟病抗性
1 材料和方法
1.1 材 料
R996转 Pi9基因植株;含 Pi9基因植株75–1–127;稻瘟病菌生理小种110–2。
1.2 方 法
1.2.1 潮霉素抗性筛选
选择 T1代符合孟德尔分离比(1∶2∶1)的转基因植株,经几代繁殖筛选,获得的单株成熟种子去壳,用体积比为 30%的次氯酸钠溶液消毒处理 30 min,无菌水清洗5~6次后,将种子转移到含有50 mg/L潮霉素的1/2MS培养基中,置于26 ℃的光照培养箱中培养,10 d后观察筛选情况。
1.2.2 GUS 活性测定
转基因水稻细胞中GUS基因的表达基本按文献[12]的方法测定。以受体 R996的根和叶作染色对照,将经过潮霉素筛选的转基因植株的根和叶片剪取一小部分,放入 1.5 mL离心管中,加入适量X–Gluc染色液,在37 ℃恒温箱中保温过夜。倒掉染色液,用70%的乙醇脱色。
1.2.3 PCR检测
按CTAB法提取经过潮霉素筛选和GUS活性测定的转基因苗和对照受体植株 R996的基因组DNA。PCR引物选用潮霉素抗性基因引物HPT–1F/HPT–1R,其扩增引物序列为:HPT–1F,5´–TACCTCTACACAGCCATC–3´;HPT–1R,5´–TAT GTCCTGCGGGTAAAT–3´。扩增片段大小为837 bp。分别以受体为阴性对照,以质粒pCAMMBIA1301–Pi9为阳性对照。PCR扩增反应条件:94 ℃预变性3 min,94 ℃变性50 s,52 ℃退火50 s,72 ℃延伸60 s,循环37次;最后72 ℃延伸10 min。
1.2.4 转基因植株的稻瘟病抗性鉴定
为检测呈阳性的转Pi9基因R996植株是否具有稻瘟病抗性,用稻瘟病菌小种110–2对上述3种方法均鉴定为阳性的转Pi9基因R996株系进行喷雾接种,并以具有Pi9基因的植株75–1–127和受体R996为抗、感病对照。具体做法:将稻瘟病菌接种在燕麦片番茄汁琼脂培养基上,于 26 ℃培养 7~10 d,用0.1‰的Tween20水溶液洗下分生孢子,接种孢子浓度约为2×105个/mL。清水浸种24 h,34~37 ℃催芽20 h左右至种子露白,播种后放入26 ℃的光照培养箱中生长,幼苗长到 3~4叶期时,进行稻瘟病菌喷雾接种。接种后 26 ℃保湿暗培养24 h,然后光照培养7 d,调查发病情况。
2 结果与分析
2.1 转Pi9基因R996株系的潮霉素筛选结果
将受体和转基因植株的种子消毒后,在潮霉素浓度为50 mg/L的1/2MS培养基中筛选,受潮霉素抑制,受体R996不萌发,而转Pi9基因R996种子的根能正常生长,还能长出叶片(图1),说明转Pi9基因R996植株中由于具有hpt基因使植株具有潮霉素抗性。

图1 潮霉素抗性筛选转基因植株Fig.1 Screening of transgenic plants using hygromycin
2.2 GUS基因在转Pi9基因R996植株中的瞬时表达
GUS染色结果显示,R996转Pi9基因植株后代株系的根呈明显蓝色(图2–A),而受体R996的根不显色(图2–B)。同样,转Pi9基因R996植株叶片被剪两端也呈现蓝色,中间部分叶片因蜡质层阻挡,染色液难到达,所以基本无色(图 2–C)。对照R996叶片被剪两端没有蓝色(图 2–D)。表明 GUS基因在转基因植株后代中能高效表达,目的基因Pi9在后代中稳定遗传。2.3 转Pi9基因R996植株的PCR检测结果

图2 转基因植株GUS检测Fig.2 Result of GUS detection of transgenic plants
由于Pi9基因同源片段在水稻基因组中广泛分布,较难设计Pi9的特异引物,故选用潮霉素抗性基因引物。T1代表现为1∶2∶1孟德尔分离规律的转Pi9基因R996植株后代,经PCR检测均能扩增出837 bp的特异条带,表明转Pi9基因的R996植株后代不再发生分离。这与 GUS染色和潮霉素筛选结果一致,即有 GUS显色和潮霉素抗性的植株均可扩增出837 bp的目的片段,而受体R996基因组DNA不能扩增出该目的条带(图3)。
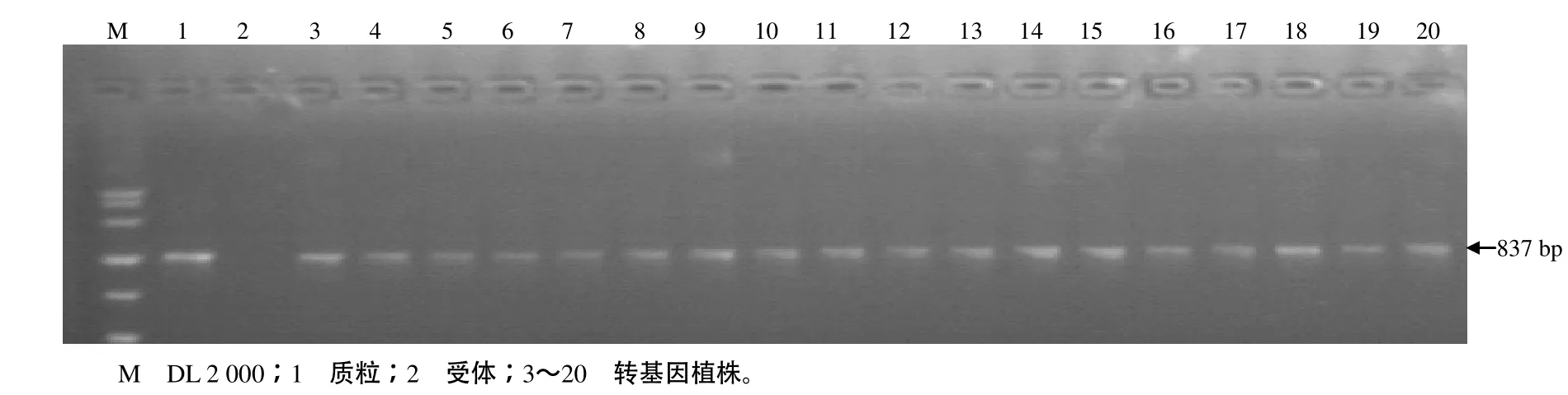
图3 转基因植株的PCR检测结果Fig.3 PCR result for transgenic plants
2.4 转Pi9基因R996植株的稻瘟病抗性检测

图4 稻瘟病菌接种结果Fig.4 Result for rice blast inoculation test
转Pi9基因R996植株的稻瘟病抗性检测结果(图4)表明,转基因植株没有出现明显的稻瘟病病斑,对稻瘟病表现出强抗性。含有 Pi9基因的75–1–127植株叶片也没有病斑,而受体R996病斑明显,感病严重。
综合上述 GUS染色、潮霉素抗性检测、PCR分析以及稻瘟病抗性检测结果,说明Pi9基因不仅整合到转基因植株的染色体上,而且成功在子代中稳定遗传,并表现出对稻瘟病的强抗性。
3 讨 论
有研究[13]表明,稻瘟病广谱抗性是持久抗性的重要组成部分。随着分子生物学的发展,广谱抗性基因在抗病育种中的应用越来越重要。Pi9基因作为稻瘟病广谱、持久抗源,目前相对水稻其他功能基因在育种上的应用更有效、更广泛。潘素君等[14]将Pi9基因导入籼稻1701品种获得的转基因植株对稻瘟病菌具有很强的抗性。梁海福等[15]利用常规回交育种结合分子标记辅助选择技术,将高抗水稻白叶枯病Xa4和Xa23基因、广谱高抗稻瘟病Pi9基因聚合到同一株系中,获得了三基因聚合的纯合株系,对稻瘟病的抗性与供体亲本 75–1–127水平相当。倪大虎等[16–18]利用分子标记辅助选择将广谱高抗稻瘟病的 Pi9基因和高抗白叶枯病的 Xa21和Xa23基因聚合到优良株系中,获得了多抗基因的优良新株系。文绍山等[19]将Pi9基因导入水稻品种泸恢17,经田间抗性鉴定,抗性基因纯合株系的稻瘟病抗性比受体明显提高。殷得所等[20]将Pi9基因通过杂交导入水稻品种扬稻6号和R6547,经病圃鉴定有Pi9基因株系的稻瘟病抗性水平较受体品种扬稻6号和R6547有不同程度的提高。
本研究利用农杆菌介导法将Pi9基因导入籼稻R996,通过GUS染色、潮霉素抗性筛选和PCR检测,鉴定出转基因植株的纯合系。经稻瘟病菌接种检测表明,转Pi9基因R996植株对稻瘟病抗性比受体R996明显增强。下一步将选用高产、优质但抗性差的水稻品种与转基因纯合系进行杂交育种,以期获得抗稻瘟病新种质。
[1] Baker B,Zambryski P,Staskawicz B,et al.Signaling in plant-microbe interactions[J].Science,1997,276:726–733.
[2] Ballini E,Morel J B,Droc G,et al.A genome-wide meta-analysis of rice blast resistance genes and quantitative trait loci provide new insights into partial and complete resistance[J].Mol Plant Microbe Interact,2008,21(7):859–868.
[3] Moffat A S.Mapping the sequence of disease resistance[J].Science,1994,265:1804–1805.
[4] Liu Y,Liu B,Zhu X,et al.Fine-mapping and molecular marker development for Pi56(t),a NBS–LRR gene conferring broad-spectrum resistance to Magnaporthe oryzae in rice[J].Theoretical and Applied Genetics,2013,126(4):985–998.
[5] Wang Z X,Yano M,Yamanouchi U,et al.The Pib gene for rice blast resistance belongs to the nucleotide binding and leucine-rich repeat class of plant disease resistance genes[J].The Plant Journal,1999,19(1):55–64.
[6] Qu S,Liu G,Zhou B,et al.The broad-spectrum blast resistance gene Pi9 encodes a nucleotide-binding siteleucine-rich repeat protein and is a member of a multigene family in rice[J].Genetics,2006,172:1901–1914.
[7] Zhou B,Qu S,Liu G,et al.The eight amino acid differences within three leucine-rich repeats between Pi2 and Piz-t resistance proteins determine the resistance specificity to Magnaporthe grisea[J].Molecular Plant-Microbe Interactions,2006,19(11):1216–1228.
[8] Hua L,Wu J,Chen C,et al.The isolation of Pi1,an allele at the Pik locus which confers broad spectrum resistance to rice blast[J].Theoretical and Applied Genetics,2012,125(5):1047–1055.
[9] Hayashi N,Inoue H,Kato T,et al.Durable panicle blast–resistance gene Pb1 encodes an atypical CC-NBSLRR protein and was generated by acquiring a promoter through local genome duplication[J].The Plant Journal,2010,64(3):498–510.
[10] 何秀英,王玲,吴伟怀,等.水稻稻瘟病抗性基因的定位、克隆及育种应用研究进展[J].中国农学通报,2014,30(6):1–12.
[11] 潘素君,戴良英,刘雄伦,等.广谱抗稻瘟病基因Pi9对籼稻的转化研究[J].分子植物育种,2006,4(5):650–654.
[12] Jefferson R A.Assaying chimeric genes in plants:The GUS gene fusion system[J].Plant Molecular Biology Reporter,1987,5(4):387–405.
[13] Wu J L,Fan Y Y,Li D B,et al.Genetic control of rice blast resistance gene in the durably resistant cultivar Gumei 2 against multiple isolates[J].Theoretical and Applied Genetics,2005,111(1):50–56.
[14] 潘素君,戴良英,刘雄伦,等.籼稻1701转Pi9基因株系的遗传分析及抗瘟性鉴定[J].湖南农业大学学报:自然科学版,2008,34(5):579–582.
[15] 梁海福,阎 勇,秦钢,等.利用分子标记辅助选择聚合Xa4、Xa23和Pi9基因[J].广西农业科学,2010,41(5):408–411.
[16] 倪大虎,易成新,李莉,等.利用分子标记辅助选择聚合水稻基因 Xa21 和 Pi9 (t) [J].分子植物育种,2005,3(3):329–334.
[17] 倪大虎,易成新,杨剑波,等.利用分子标记辅助选择聚合Pi9 (t) 和Xa23[J].分子植物育种,2007,5(4):491–496.
[18] 倪大虎,易成新,李莉,等.分子标记辅助培育水稻抗白叶枯病和稻瘟病三基因聚合系[J].作物学报,2008,34(1):100–105.
[19] 文绍山,高必军.利用分子标记辅助选择将抗稻瘟病基因 Pi–9 (t)渗入水稻恢复系泸恢 17[J].分子植物育种,2012,10(1):42–47.
[20] 殷得所,夏明元,李进波,等.抗稻瘟病基因 Pi9的STS连锁标记开发及在分子标记辅助育种中的应用[J].中国水稻科学,2011,25(1):25–30.
责任编辑:苏爱华
英文编辑:罗 维
Screening of homozygous Pi9 gene transgenic lines of Indica cultivar R996 and identification of its rice blast resistance
PAN Su-juna, LI Weia,b, GAO Jiab, DAI Liang-yinga,b*
(a.Crop Gene Engineering Key Laboratory of Hunan Province; b.College of Plant Protection, Hunan Agricultural University, Changsha 410128,China)
Pi9, the first cloned gene with broad-spectrum resistance to rice blast, was introduced into Indica cultivar R996 via Agrobacterium-mediated transformation. The subsequent genetic analysis showed that Pi9 was successfully introduced into the genome of the receptors, which displayed stable expression of Pi9 at transcription level. The offspring of the transgenic plants were analyzed by hygromycin resistance assay, GUS staining and PCR detection. And the homozygous Pi9/R996 transgenic lines were identified from the T1generation showing 1∶2∶1 Mendel segregation through rice blast resistance assay conducted on Pi9 positive transformants.
Indica cultivar R996; Pi9 gene; transgenic plant; the resistance of rice blast
10.13331/j.cnki.jhau.2014.06.005
投稿网址:http://www.hunau.net/qks
S511.2+10.34
A
1007−1032(2014)06−0589−04
稻瘟病是水稻的三大病害之一,常给水稻生产造成巨大的损失[1–3]。利用转基因技术将抗稻瘟病基因转入优质的易感病的水稻品种中,通过杂交、回交等方法获得高抗品系是减少稻瘟病病害的有效途径之一。目前,在水稻的不同染色体上已定位了超过86个稻瘟病主效抗性基因,350个微效基因[2,4]。1999年,通过图位克隆得到了第1个稻瘟病抗性基因Pib[5]。迄今,采用此方法已从不同的水稻品种中成功分离克隆了Pi9[6]、Pi2[7]、Pi1[8]、Pb1[9]等22个稻瘟病抗性基因[10]。稻瘟病菌生理小种的多样性及稻瘟病与抗性基因之间的协同进化关系,限制了稻瘟病抗性基因和抗性品种的推广应用。
Pi9基因位于水稻第6号染色体短臂近着丝粒区域的Pi2/9位点,基因全长约9.5 kb,包括2个长度分别为5 362 bp和128 bp的内含子,以及由3 099 bp的编码区和910 bp 3' UTR组成的cDNA。Pi9编码1个由1 032个氨基酸组成的蛋白产物。该产物羧基端含有富亮氨酸重复域(LRRs),在氨基端含有核苷酸结合位点(NBS)。目前,已证实Pi9基因对100多个稻瘟病菌小种具有抗性[6]。笔者采用农杆菌介导法将Pi9转入籼稻品种R996[11],并进行了转基因植株纯系筛选以及稻瘟病抗性的鉴定,以期通过下一步遗传杂交方法获得高抗稻瘟病水稻新品种。
2013–06–23
国家自然科学基金项目(31371246,31300250);转基因生物新品种培育重大专项(2013ZX08001–002,2012ZX08009001)
潘素君(1973—),女,湖南宁远人,博士,副研究员,主要从事分子生物学研究,sujunpan@163.com;*通信作者,daily@ hunau.net
猜你喜欢
古今农业(2022年2期)2022-08-15
亚热带植物科学(2022年1期)2022-05-17
安徽农学通报(2022年8期)2022-05-06
作物学报(2022年6期)2022-04-08
四川农业科技(2019年5期)2019-07-01
河南农业(2016年6期)2016-11-26
西南农业学报(2016年4期)2016-05-17
浙江柑橘(2016年1期)2016-03-11
现代农业(2016年5期)2016-02-28
现代农业(2015年1期)2015-02-28
