
退化伊犁绢蒿荒漠草地特征植物光谱特征
2014-12-24 00:53靳瑰丽何龙安沙舟范燕敏武鹏飞
草业科学 2014年10期
靳瑰丽,何龙,安沙舟,范燕敏,武鹏飞
(1.新疆农业大学草业与环境科学学院新疆草地资源与生态重点实验室,新疆乌鲁木齐830052;2.乌鲁木齐气象卫星地面站,新疆乌鲁木齐830011)
我国仅分布于新疆的伊犁绢蒿(Seriphidium transiliense)荒漠草地属于中亚气候型荒漠,是新疆主要的春秋放牧场[1]。但近年来,该类草地因气候变化和超载过牧而严重退化。针对该类草地,已有从退化程度[2]、种子和幼苗[3]、植物生理[1]、牧草营养[4]、植物多样性[5]、群落演替特征[6]、种群生态位[7]、载畜力[8]等角度的研究。从遥感应用角度来看,已有很多以遥感目视解译结合地面调查对退化草地的研究[9-10]。然而,在利用植被指数对退化草地进行等级划分时,时间段较窄,仅限于7―8月,而利用高光谱数据进行模式识别分析,不仅在不同季节鉴别精度可超过90%[10],而且高光谱数据还可反映植物的盖度、生物量,并提供荒漠化监测中所需的特征植物指标[11]以及改善对特征植物的识别与分类精度[12],在退化草地的监测、评价及特征植物的识别与分类中有其他遥感信息源不可比拟的优势。而对草地单物种进行高光谱特征的分析可为利用高光谱遥感数据进行大面积草地物种识别与动态监测提供借鉴。本研究从光谱角度分别对该类退化草地8种特征植物(包括减少种、增加种、侵入种)长势较好时期的原始光谱、包络线去除后光谱及一阶微分光谱曲线特征进行对比分析,以期为实现该类退化草地高光谱遥感所需特征植物地面高光谱特征的提取与分析提供参考依据。
1 材料与方法
1.1 研究区域概况
研究区位于天山北坡中段昌吉市三工镇以南2 km处的山前倾斜冲积-洪积扇,地处87.136 02°―87.142 16°E,43.858 31°― 43.870 11°N,海拔825 ~897 m。具有中亚荒漠气候的特征,年降水量为180~190 mm,年均温为6.5℃。土壤是灰漠土,成土母质为黄土状物质,土层较厚。该研究区是伊犁绢蒿荒漠草地的典型分布区,由于放牧过度而退化[13]。
1.2 测定方法
基于Dykterhuis关于物种划分的草原生态学评价方法[14],从光谱角度分别对该类退化草地中减少种、增加种、侵入种的原始、包络线去除及一阶微分变换后光谱特征进行对比分析。2011年4、5月下旬,通过使用美国SVC HR-768便携式光谱仪(光谱范围350-2 500 nm)对伊犁绢蒿荒漠草地退化过程中的减少种伊犁绢蒿,侵入种萹蓄(Polygonum aviculare)和骆驼蓬(Peganum harmala),增加种叉毛蓬(Petrosimonia sibirica)、角果藜(Ceratocarpus arenarius)、画眉草(Eragrostis pilosa)、地肤(Kochia scoparia)、弯果胡卢巴(Trigonella arcuata)共8种特征植物冠层的地面高光谱反射率进行测定。选择在晴朗无云的天气采集光谱,同时为减少不同太阳高度角对反射率的影响,测定时间为北京时间12:00―14:00。在测定光谱反射率时,探头与地面始终保持垂直,高度1 m,在每个退化梯度选取各个典型特征植物群落分布最集中部分进行6个重复的测定,以确保所测光谱为单一植物,每个重复有10条光谱反射率曲线。
1.3 数据分析
利用白色参照板获取绝对反射率,计算出8种特征植物相对反射特征值,并将每个重复测定的10条光谱反射率曲线作异常曲线去除后,取平均作为特征植物的光谱反射率曲线;光谱反射率数据分别以原始、包络线去除以及一阶微分变换后光谱曲线3种方法进行处理;利用包络线去除法对(350-1 800 nm)原始高光谱数据去噪处理,再将原始高光谱反射率数据进行一阶微分的数学变换以后加以综合对比分析;包络线去除的全过程在ENVI 4.5中进行,分析数据使用Excel 2003,再利用Sigma Plot 10.0软件制图。
2 结果与分析
2.1 退化伊犁绢蒿荒漠草地减少种
伊犁绢蒿是蒿类荒漠草地植物组成中的典型代表种,它作为减少种其群落基本性状在该类草地退化过程中有偏离顶级的迹象,退化严重时甚至会被新的类型替代;以它为代表的半灌木类群在季节畜牧业中有极为重要的作用。
伊犁绢蒿的光谱特征曲线有明显的“红边”效应,红边位置在678-760 nm(图1A)。在可见光波段(350-760 nm),伊犁绢蒿的反射率较弱。在叶绿素强吸收的450-670 nm波段内,502-580 nm波段为伊犁绢蒿的一个明显叶绿素吸收峰。在近红外波段(760-926 nm),呈现出连续的强反射率(58% ~61%);在700-1 300 nm植被强烈反射带波段的930-1 300 nm波段,伊犁绢蒿的光谱特征曲线都有两个明显的波峰和波谷,而且呈现出先升高后下降的趋势,两峰中第1个波峰在1 083 nm处反射率为65.71%,比1 273 nm处的第2峰高3.3百分点。在1 300-1 400 nm波段附近为伊犁绢蒿的水吸收带,在1 448 nm处反射率为30.52%。
经过包络线去除处理后伊犁绢蒿的红边位置显示为681-751 nm波段,包络线去除后的光谱反射曲线峰谷特点较为明显(图1B)。在叶绿素强吸收的450-670 nm波段内,伊犁绢蒿叶绿素吸收峰显示在536 nm且峰值为0.666 8。
经过一阶微分变换后在400-760 nm波段间曲线的差异明显变大,900-1 000 nm波段附近波动剧烈,723 nm达到光谱一阶微分最大值即伊犁绢蒿的红边所在位置为718 nm,红边幅值为0.925 4(图1C)。伊犁绢蒿原始光谱经过一阶微分变换后的叶绿素吸收峰峰值出现在524 nm,峰值为0.191 6。
2.2 退化伊犁绢蒿荒漠草地侵入种
一年生草本植物萹蓄与多年生草本植物骆驼蓬是伊犁绢蒿荒漠草地的侵入种。随着该类草地长期处于退化状态,萹蓄、骆驼蓬逐渐成为该类草地常见的植物。
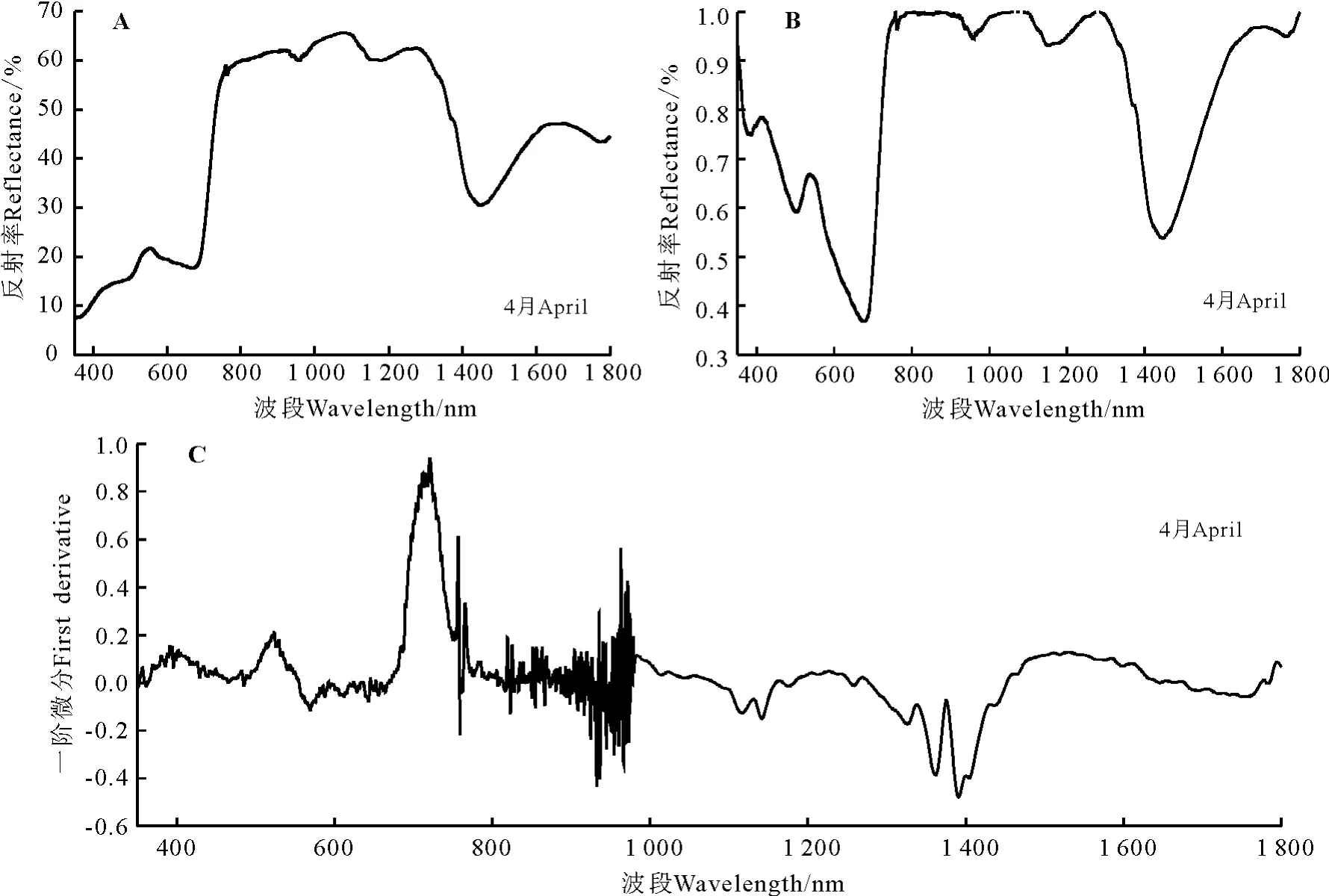
图1 伊犁绢蒿高光谱特征Fig.1 Spectral features of Seriphidium transiliens
2.2.1 萹蓄高光谱特征分析 萹蓄的光谱特征曲线有明显的“红边”效应,红边位置在680-760 nm之间(图2A)。在可见光波段(350-770 nm),萹蓄的反射率较弱。在叶绿素强吸收的450-670 nm波段内,506-580 nm波段有萹蓄的一个明显叶绿素吸收峰。在近红外波段(760-960 nm),呈现出强反射;在700-1 300 nm植被强烈反射带波段的960-1 300 nm波段,萹蓄的光谱特征曲线都有两个明显的波峰和波谷,而且呈现出先升高后下降的趋势,第1个波峰在1 078.68 nm处达到45.97%的反射率最大值。在1 300-1 400 nm波段附近为萹蓄的水吸收带,在1 445 nm处反射率为17.53%。
经过包络线去除后的光谱反射曲线具有峰谷的特点明显,经过包络线去除处理后萹蓄的红边位置显示为更为清晰的680-760 nm波段(图2B)。在叶绿素强吸收的450-670 nm波段内,萹蓄叶绿素吸收峰显示的更为清楚在543 nm。
经过一阶微分变换后在400-760 nm波段间曲线的差异明显变大,900-1 000 nm波段附近波动剧烈,708 nm达到光谱一阶微分最大值即萹蓄的红边所在位置为708 nm,红边幅值为0.685 4(图2C)。萹蓄原始光谱经过一阶微分变换后的叶绿素吸收峰峰值出现在521 nm,峰值为0.142 0。

图2 萹蓄高光谱特征Fig.2 Spectral features of Polygonum aviculare
2.2.2 骆驼蓬高光谱特征分析 骆驼蓬的光谱特征曲线有明显的“红边”效应,红边位置在680-760 nm(图3A)。在可见光波段(350-760 nm),骆驼蓬的反射率较弱。在叶绿素强吸收的450-670 nm波段内,512-588 nm波段为骆驼蓬的1个明显叶绿素吸收峰。在近红外波段(760-926 nm)呈现出连续的强反射率;在700-1 300 nm植被强烈反射带波段的930-1 300 nm波段,骆驼蓬的光谱特征曲线都有两个明显的波峰和波谷,而且呈现出先升高后下降的趋势,两峰中第1个波峰在1 073 nm处反射率为35.56%,比1 261 nm处的第2峰高10个百分点。在1 300-1 400 nm波段附近为骆驼蓬的水吸收带,在1 448 nm处反射率为3.72%。
经过包络线去除后的光谱反射曲线峰谷特点较为明显,经过包络线去除处理后骆驼蓬的红边位置显示为681-758 nm波段(图3B)。在叶绿素强吸收的450-670 nm波段内,骆驼蓬叶绿素吸收峰显示的更为清楚,在549 nm处峰值为0.303 7。
经过一阶微分变换后在400-760 nm波段间曲线的差异明显变大,900-1 000 nm波段附近波动剧烈,724 nm达到光谱一阶微分最大值即骆驼蓬的红边所在位置为718 nm,红边幅值为 0.799 8(图3C)。骆驼蓬原始光谱经过一阶微分变换后的叶绿素吸收峰峰值出现在524 nm,峰值为0.111 9。
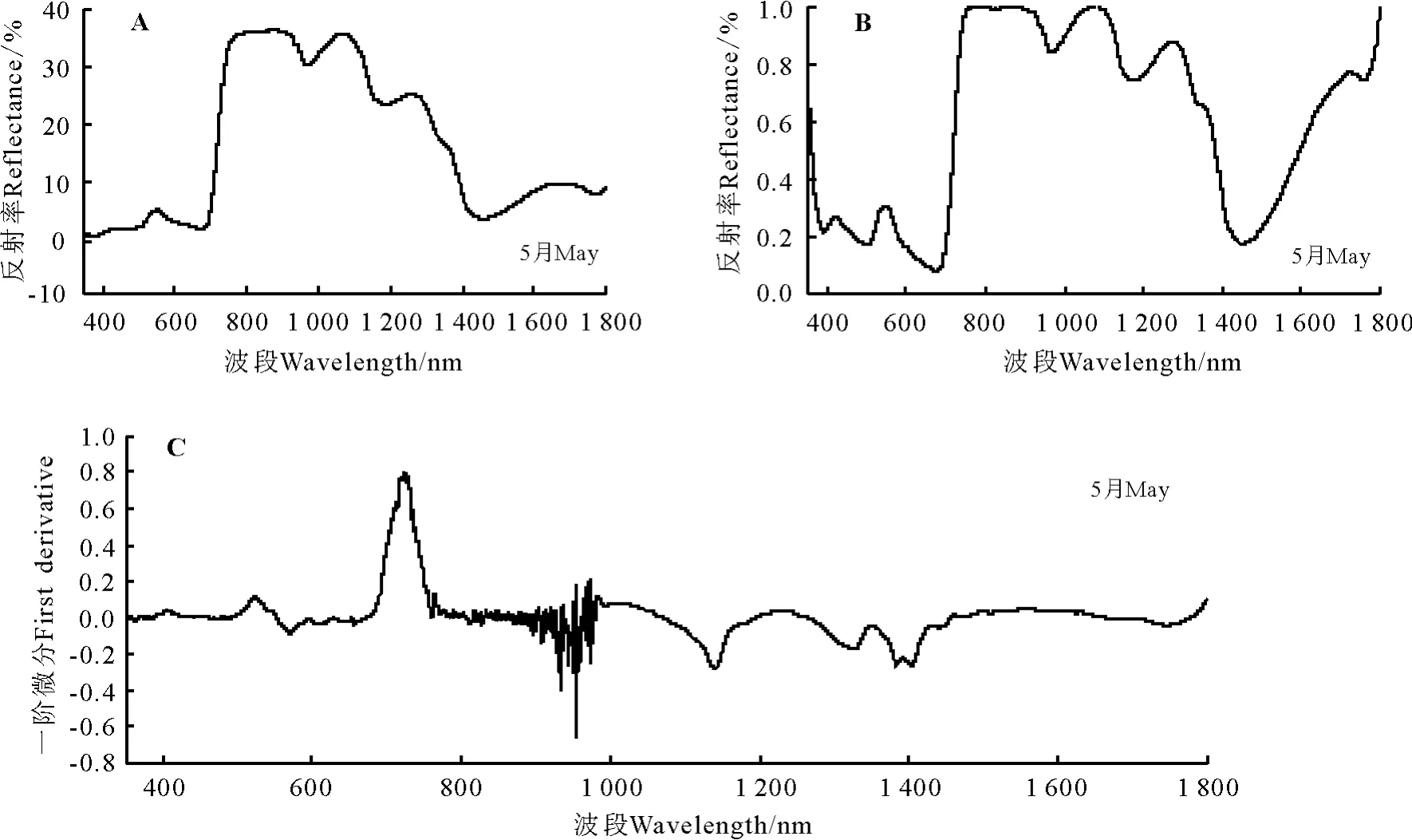
图3 骆驼蓬高光谱特征Fig.3 Spectral features Peganum harmala
2.3 退化伊犁绢蒿荒漠草地增加种
一年生长期营养期草本类群中禾草亚类的画眉草,杂草亚类的地肤、叉毛蓬、角果藜,短生、类短生中生植物弯果胡卢巴是退化伊犁绢蒿荒漠草地的增加种。这两类植物类群对于荒漠草地而言都有重要的饲用价值,对指示草地的退化也有重要作用。
2.3.1 叉毛蓬高光谱特征分析 叉毛蓬的光谱特征曲线有明显的“红边”效应,红边位置在690-750 nm(图4A)。在可见光波段(350-760 nm),叉毛蓬的反射率较弱。在叶绿素强吸收的450-670 nm波段内,叉毛蓬未见明显叶绿素吸收峰。在近红外波段(760-1 276 nm),保持了较高的反射强度;在700-1 300 nm植被强烈反射带波段的960-1 300 nm波段,叉毛蓬的光谱特征曲线出现了一个明显的波谷,而且除一个波谷外呈现持续升高最后下降的趋势。在1 300-1 400 nm波段附近为叉毛蓬的水吸收带,在1 430 nm处反射率为18.89%。
经过包络线去除后的光谱反射曲线峰谷特点明显,经过包络线去除处理后叉毛蓬的红边位置显示为更为清晰的681-751 nm波段(图4B)。在叶绿素强吸收的450-670 nm波段内,叉毛蓬叶绿素吸收峰显示的更为清楚,在546 nm处峰值为0.884 9。
经过一阶微分变换后在400-760 nm波段间曲线的差异明显变大,900-1 000 nm波段附近波动剧烈,718 nm达到光谱一阶微分最大值即萹蓄的红边所在位置为718 nm,红边幅值为0.188 5(图4C)。叉毛蓬原始光谱经过一阶微分变换后的叶绿素吸收峰峰值出现在518 nm,峰值为0.057 7。
2.3.2 角果藜高光谱特征分析 角果藜的光谱特征曲线有明显的“红边”效应,红边位置在680-761 nm(图5A)。在可见光波段(350-761 nm),角果藜的反射率较弱。在叶绿素强吸收的450-670 nm波段内,537-580 nm、580-615 nm波段有角果藜4个明显的叶绿素吸收峰。在近红外波段(761-1 300 nm),呈现出强反射;在700-1 300 nm植被强烈反射带波段的960-1 300 nm波段,角果藜的光谱特征曲线有一个不太明显的波谷,而且呈现出先下降后升高的趋势,在1 300 nm处达到29.19%的反射率值。在1 300-1 400 nm波段附近为角果藜的水吸收带,在1 445 nm处反射率为24.43%。角果藜的反射率最大值出现在1 651 nm,为29.80%。
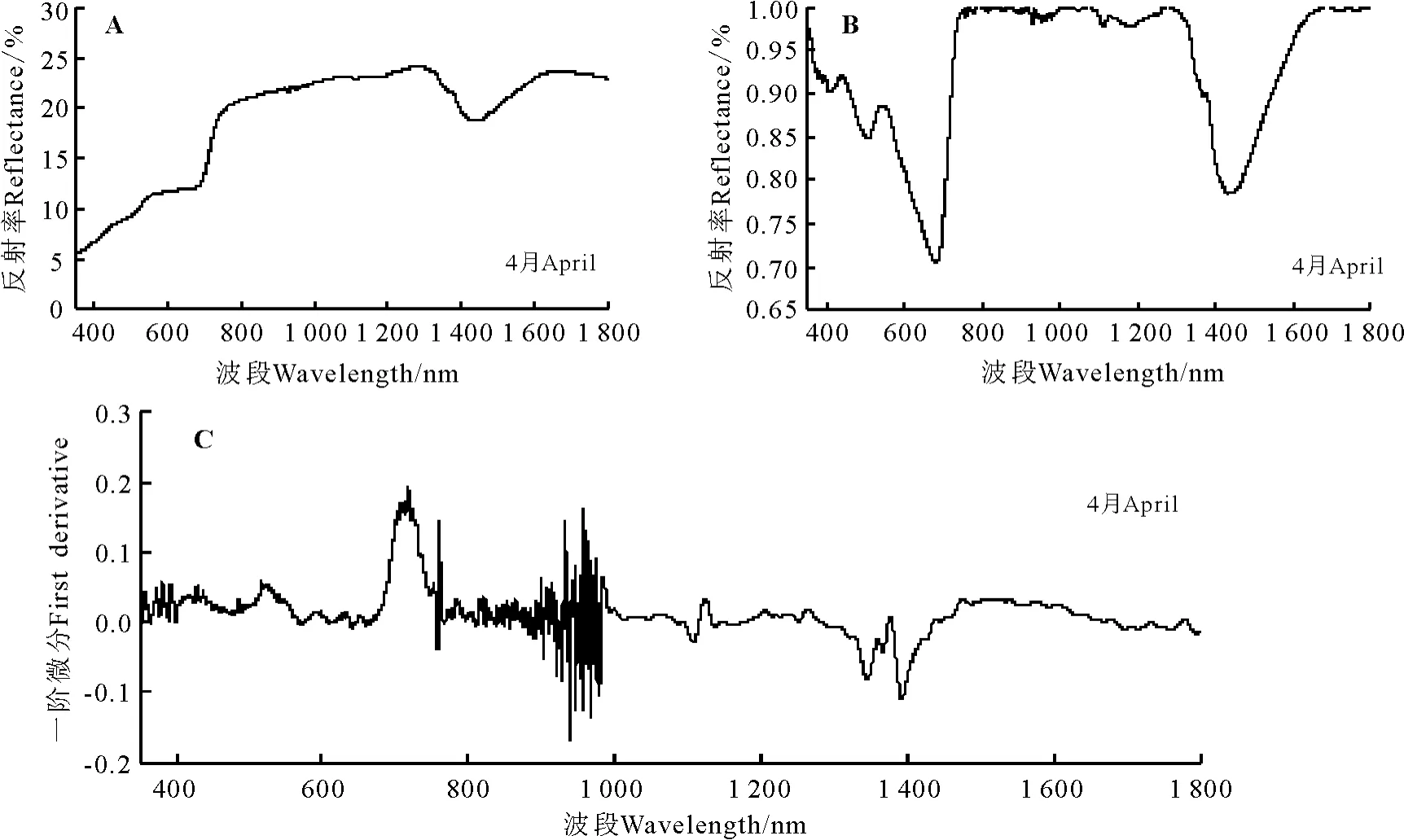
图4 叉毛蓬高光谱特征Fig.4 Spectral features of Petrosimonia sibirica
经过包络线去除后的光谱反射曲线峰谷特点较不明显,经过包络线去除处理后角果藜的红边位置显示为684-751 nm波段(图5B)。在叶绿素强吸收的450-670 nm波段内,角果藜叶绿素吸收峰显示在558 nm。
经过一阶微分变换后在400-760 nm波段间曲线的差异明显变大,900-1 000 nm波段附近波动剧烈,718 nm达到光谱一阶微分最大值即角果藜的红边所在位置为718 nm,红边幅值为0.132 7(图5C)。角果藜原始光谱经过一阶微分变换后的叶绿素吸收峰谷变化差异变大,4个峰分别出现在515、549、586 和 626 nm,峰值分别为 0.077 9、0.129 1、0.125 6和0.122 0。
2.3.3 地肤高光谱特征分析 地肤的光谱特征曲线有明显的“红边”效应,红边位置在687-751 nm(图6A)。在可见光波段(350-760 nm),地肤的反射率较弱。在叶绿素强吸收的450-670 nm波段内,地肤未见明显叶绿素吸收峰。在近红外波段(751-1 350 nm),保持了较高的反射强度;在700-1 300 nm 植被强烈反射带波段的1 114-1 255 nm波段,地肤的光谱特征曲线出现了一个明显的波谷,而且除一个波谷外呈现持续升高最后下降的趋势。在1 350-1 400 nm波段附近为地肤的水吸收带,在1 430 nm处反射率为29.11%。
经过包络线去除后的光谱反射曲线峰谷特点明显,经过包络线去除处理后地肤的红边位置在687-751 nm波段显示的更为清晰(图6B)。在叶绿素强吸收的450-670 nm波段内,地肤叶绿素吸收峰显示的更为清楚,在564 nm处峰值为0.999 5。
经过一阶微分变换后在400-760 nm波段间曲线的差异明显变大,900-1 000 nm波段附近波动剧烈,718 nm达到光谱一阶微分最大值即地肤的红边所在位置为718 nm,红边幅值为0.183 9(图6C)。地肤原始光谱经过一阶微分变换后的叶绿素吸收峰峰值出现在515 nm处,峰值为0.096 7。
2.3.4 弯果胡卢巴高光谱特征分析 弯果胡卢巴的光谱特征曲线有明显的“红边”效应,红边位置在680-760 nm(图7A)。在可见光波段(350-770 nm),弯果胡卢巴的反射率较弱。在叶绿素强吸收的450-670 nm波段内,500-580 nm波段有弯果胡卢巴的1个明显叶绿素吸收峰。在近红外波段(760-930 nm),呈现出强反射;在700-1 300 nm植被强烈反射带波段的930-1 300 nm波段,弯果胡卢巴的光谱特征曲线都有两个明显的波峰和波谷,而且呈现出先升高后下降的趋势,第1个波峰在1 073 nm处达到 31.5%的反射率最大值。在1 300-1 400 nm波段附近为弯果胡卢巴的水吸收带,在1 448 nm处反射率为11.24%。
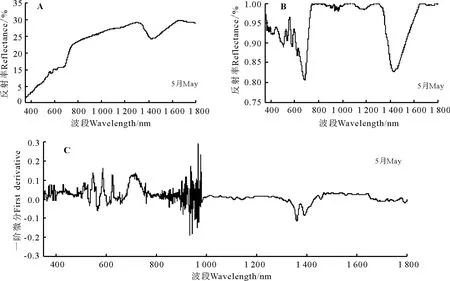
图5 角果藜高光谱特征Fig.5 Spectral features of Ceratocarpus arenariuse
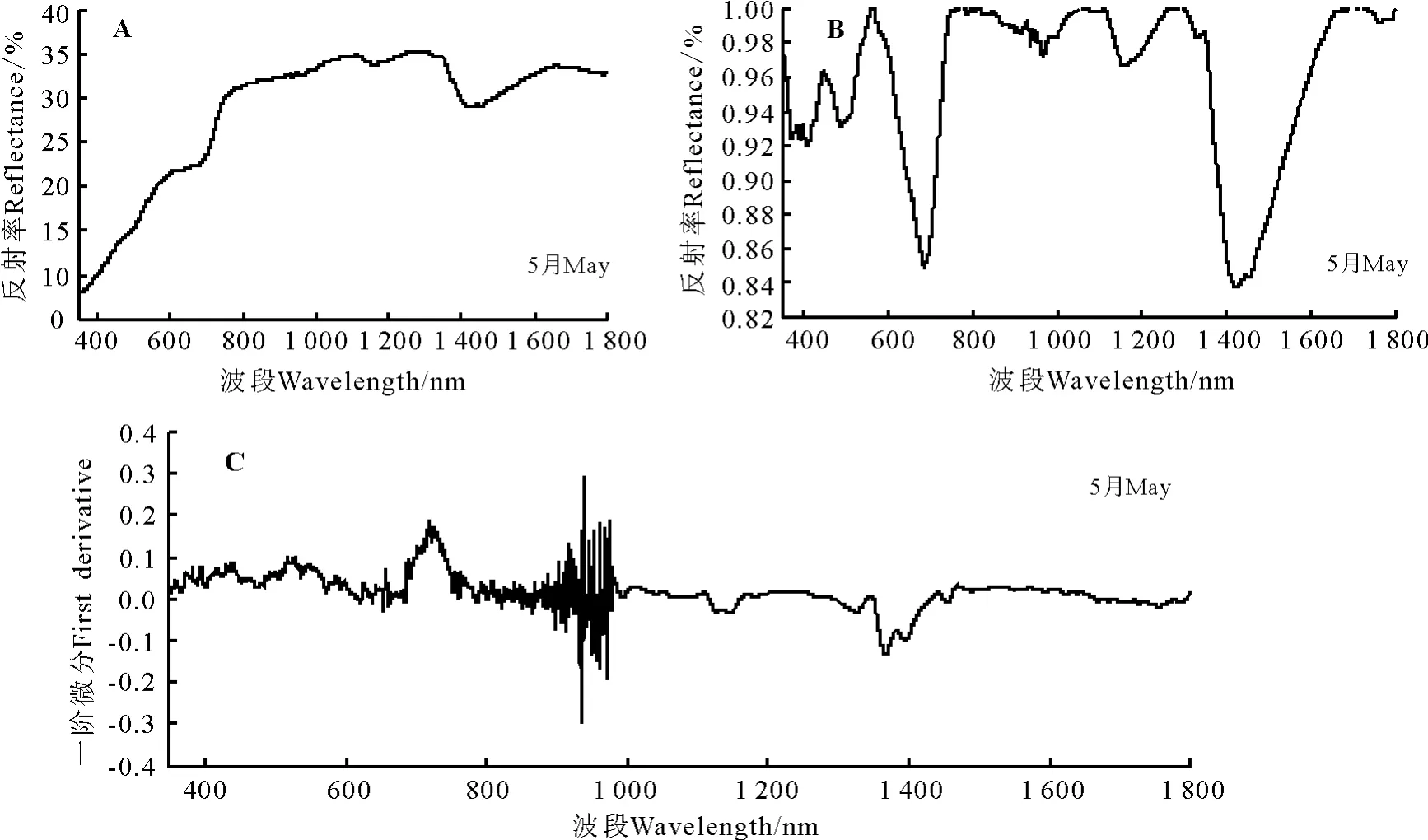
图6 地肤高光谱特征Fig.6 Spectral features of Kochia prostrate
经过包络线去除后的光谱反射曲线峰谷特点明显,经过包络线去除处理后弯果胡卢巴的红边位置显示为更为清晰的678-748 nm波段(图7B)。在叶绿素强吸收的450-670 nm波段内,弯果胡卢巴叶绿素吸收峰显示的更为清楚,在540 nm处峰值为0.584 0。
经过一阶微分变换后在400-760 nm波段间曲线的差异明显变大,900-1 000 nm波段附近波动剧烈,718 nm达到光谱一阶微分最大值即弯果胡卢巴的红边所在位置为718 nm,红边幅值为0.538 7(图7C)。弯果胡卢巴原始光谱经过一阶微分变换后的叶绿素吸收峰峰值出现在524 nm,峰值为0.109 2。
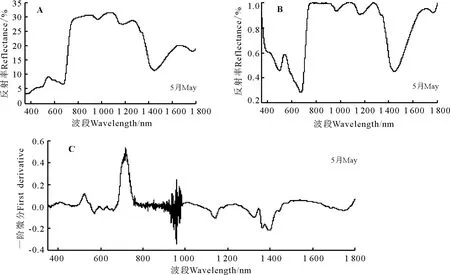
图7 弯果胡卢巴高光谱特征Fig.7 Spectral features of Trigonella arcuata
2.3.5 画眉草高光谱特征分析 画眉草的光谱特征曲线有明显的“红边”效应,红边位置在681-761 nm(图8A)。在可见光波段(350-761 nm),画眉草的反射率较弱。在叶绿素强吸收的450-681 nm波段内,画眉草有一个明显叶绿素吸收峰位于558 nm处达到12.85%的反射率。在近红外波段(761-1 350 nm),保持了较高的反射强度;在700-1 350 nm植被强烈反射带波段的936-1 350 nm波段,画眉草的光谱特征曲线出现了两个明显的波峰-波谷,而且呈现先升高最后下降的趋势。在1 350-1 400 nm波段附近为画眉草的水吸收带,在1 454 nm处反射率为22.61%。
经过包络线去除后的光谱反射曲线峰谷特点明显,经过包络线去除处理后画眉草的红边位置显示为更为清晰的678-751 nm波段(图8B)。在叶绿素强吸收的450-670 nm波段内,画眉草叶绿素吸收峰显示的更为清楚在546 nm,且峰值为0.747 5。
经过一阶微分变换后在400-760 nm波段间曲线的差异明显变大,900-1 000 nm波段附近波动剧烈,718 nm达到光谱一阶微分最大值即画眉草的红边所在位置为718 nm,红边幅值为 0.441 6(图8C)。画眉草原始光谱经过一阶微分变换后的叶绿素吸收峰峰值出现在515 nm处,峰值为0.159 4。
2.4 伊犁绢蒿荒漠草地特征植物高光谱曲线综合分析
350-1 800 nm波段间伊犁绢蒿荒漠草地8种特征植物都呈现出绿色植物高光谱曲线所具备的明显特征(图9)。伊犁绢蒿、萹蓄、弯果胡卢巴、骆驼蓬在500-600 nm波段间比其他植物表现出更明显的叶绿素吸收峰,峰值从大到小依次为伊犁绢蒿>萹蓄>弯果胡卢巴>骆驼蓬。植被强烈反射带中的760-930 nm波段间8种特征植物的反射率值最大达60%,最小约为20%,反射率从大到小依次为伊犁绢蒿>萹蓄>骆驼蓬>画眉草>地肤>弯果胡卢巴>角果藜>叉毛蓬。1 400 nm附近水分吸收带光谱反射率值最大30.52%最小3.72%,从大到小依次为伊犁绢蒿>地肤>角果藜>画眉草>叉毛蓬>萹蓄>弯果胡卢巴>骆驼蓬。
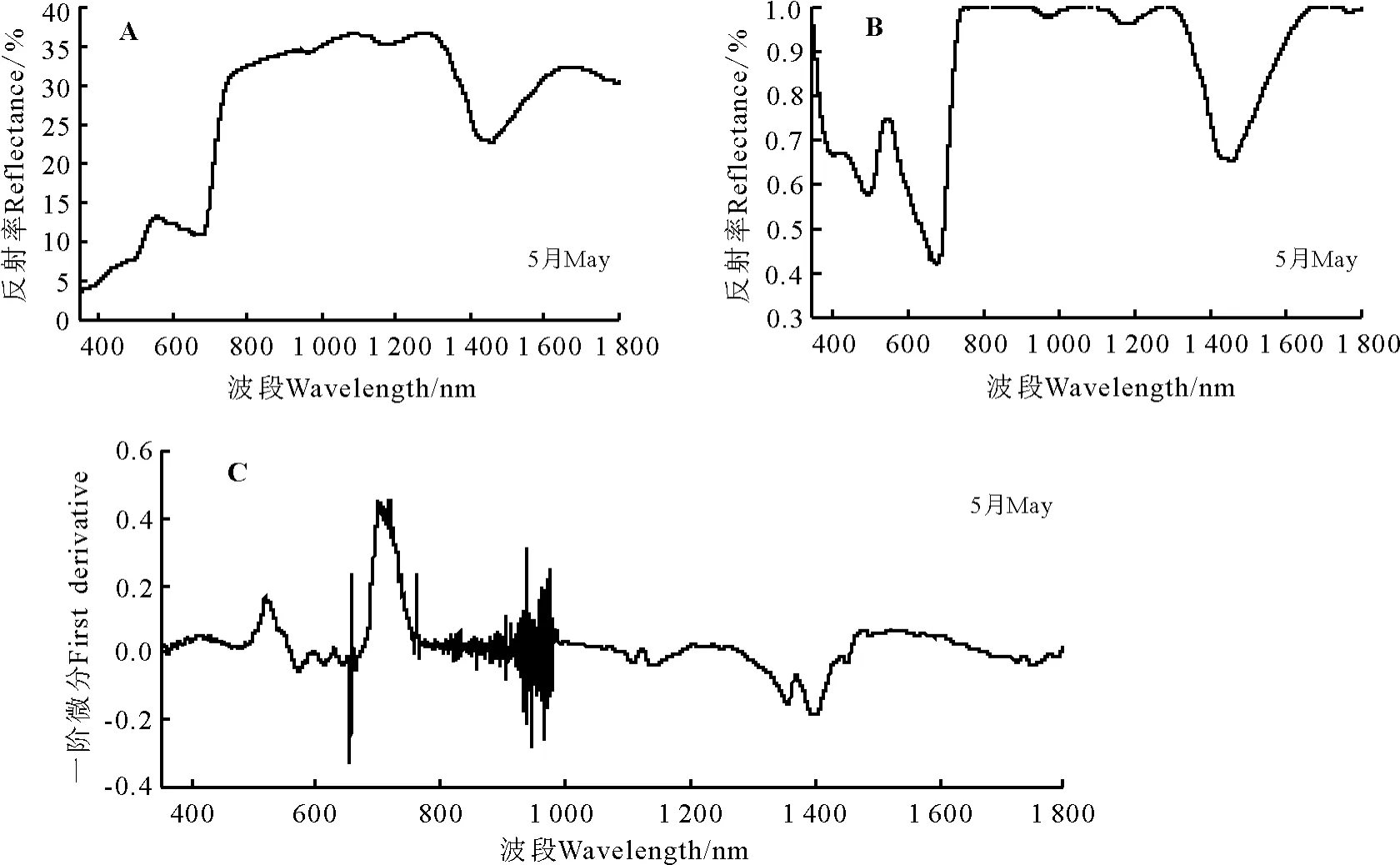
图8 画眉草高光谱特征Fig.8 Spectral features of Eragrostis pilosa
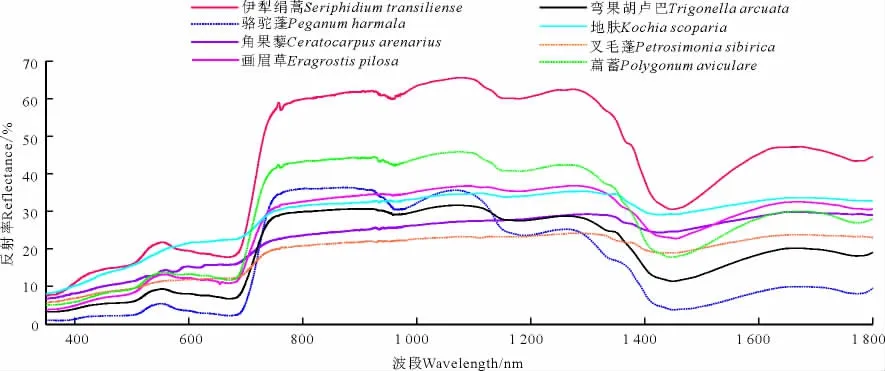
图9 8种特征植物原始高光谱曲线综合分析图Fig.9 Original spectral features of 8 characteristic species
3 讨论与结论
绿色植物光谱曲线一般都会呈现明显的“峰和谷”特征[15-16],而其反射光谱特征规律性都比较明显、独特,本研究结果与之一致。绿色植物光谱曲线在(400-760 nm)的可见光波段有一个小的反射峰,两侧有两个吸收带,即在450 nm(蓝)与670 nm(红)波段为低谷,原因是叶绿素对蓝光和红光吸收作用强,而对绿光的反射作用强[17],这与本研究中特征植物所具有的叶绿素反射峰的特征相吻合。植被光谱反射率曲线在680-760 nm波段间,近红外波段有一反射的“陡坡”称为红边[18],再至1 100 nm附近有一峰值,形成植被的独有特征,本研究中特征植物所具备的红边位置所在范围与其基本一致。植被的“红边”效应可以用于估测植被生物参数[19-23]和反映植被生长状况[24-25]。由于叶子内部液态水分的强烈吸收作用,在1 400 nm附近有明显的低谷,在1 600 nm处有反射峰。
本研究分析结果所选对应光谱数据的时间是在特征植物长势较好时期,并不能绝对代表特征植物各个生长期的光谱特征。对8种特征植物原始光谱的采集是在自然条件下进行的,由于自然因素的影响,光谱数据不可避免的会有一些噪声,所以需要对原始光谱数据进行去噪处理。光谱数据噪音消除的方法有很多,包括有中值、移动平均法、静态平均、傅立叶级数近似、Savitzky Golay、低通滤波、小波去噪、Gaussian滤波、去包络线等[26]。本研究去除包络线后的光谱曲线能有效地抑制噪音,更加突出特征植物光谱的特征信息。定量描述植被光谱红边特征的红边参数包括,1)红边幅值:红光范围(680-760 nm)内一阶导数光谱的最大值;2)红边位置:红光范围(680-760 nm)内反射光谱一阶导数(即一阶导数光谱)最大值所对应的波长。本研究还用光谱归一化微分分析技术,对反射光谱进行一阶微分[27]。从一阶微分光谱数值中精确提取了8种特征植物的光谱曲线中的红边幅值以及红边位置[28]等有效信息。有研究表明[29],结合地面光谱测定和对退化草地物种特征微弱光谱差异的定量分析,可为草地退化监测及其治理提供群落演替过程及其态势的重要指标,为草地退化监测提供新的遥感技术方法。
通过分析包络线去除后的光谱曲线,发现包络线去除后的光谱红边特征与原始光谱曲线相比显得更为清晰,原始光谱曲线中一些不明显的峰谷特征变得明显化。对于经过一阶微分变换后的光谱曲线进行分析,发现一阶微分对于精确提取该类草地植物光谱特征的红边参数特征值有着重要作用。这两种对原始光谱曲线的处理,都为该类退化草地上8种特征植物光谱曲线特征的微弱光谱差异定量分析奠定了基础。
[1]靳瑰丽,朱进忠,刘洪来,唐式敏,王彩虹.退化伊犁绢蒿荒漠草地主要植物生理生态适应性研究[J].草地学报,2011,19(1):26-30.
[2]刘洪来,朱进忠,靳瑰丽,范燕敏,王彩虹,杨兴伟.伊犁绢蒿荒漠草地退化演替序列的划分[J].新疆农业科学,2007,44(2):137-141.
[3]鲁为华,任爱天,杨洁晶,靳瑰丽.伊犁绢蒿年际结实量、土壤种子库及幼苗输入特征[J].草业科学,2013,30(3):390-396.
[4]朱进忠,吴咏梅.伊犁绢蒿荒漠不同退化阶段草地经济性状演变的分析[J].草业科学,2005,22(10):1-4.
[5]刘洪来,朱进忠,靳瑰丽,吴咏梅,范艳敏,郑伟.伊犁绢蒿荒漠不同退化阶段草地植物多样性分析[J].新疆农业科学,2007,44(5):632-636.
[6]靳瑰丽,朱进忠,陈乐宁,刘洪来,吴咏梅.伊犁绢蒿荒漠受损草地植物群落春季演变趋势分析[J].草业科学,2007,24(10):26-30.
[7]梁娜,靳瑰丽,安沙舟,赛米拉克孜·台外库力,迪丽努尔·玉苏甫江.伊犁绢蒿退化荒漠草地植物种群生态位的变化特征[J].草业科学,2013,30(12):2059-2065.
[8]刘洪来,杨学东,张靖云,靳瑰丽,陈超,宋高翔.伊犁绢蒿荒漠不同退化阶段草地特征与载畜力研究[J].草地学报,2013,21(1):50-55.
[9]钟城,何晓蓉,李辉霞.遥感技术在西藏那曲地区草地退化评价中的应用[J].遥感技术与应用,2003,18(2):99-102.
[10]王艳荣,雍世鹏.利用多时相近地面反射波谱特征对不同退化等级草地的鉴别研究[J].植物生态学报,2004,28(3):406-413.
[11]范文义,徐程扬,叶荣华,王君厚.高光谱遥感在荒漠化监测中的应用[J].东北林业大学学报,2000,28(5):139-141.
[12]Salehia B,Zoej M J V.Wavelet-based reduction of hyperspectral imagery[J].Remote Sensing of Environment,2002,36:332-337.
[13]何龙,安沙舟,靳瑰丽,范燕敏,张廷.退化伊犁绢蒿荒漠草地高光谱特征分析[J].草地学报,2014,22(2):271-276.
[14]Dyksterhuis E J.Condition and management of rangeland based on quantitative ecology[J].Journal of Range Management,1949,2:104-115.
[15]浦瑞良,宫鹏.高光谱遥感及其应用[M].北京:高等教育出版社,2000:52-96.
[16]赵春江,刘良云,周汉昌,王纪华,薛绪掌.归一化差异植被指数仪的研制与应用[J].光学技术,2004,30(3):324-326.
[17]杨可明,郭达志.植被高光谱特征分析及其病害信息提取研究[J].地理与地理信息科学,2006,22(4):31-34.
[18]童庆禧,张兵,郑兰芬.高光谱遥感——原理、技术与应用[M].北京:高等教育出版社,2006.
[19]唐延林,王秀珍,李建龙,黄敬峰,王人潮.棉花高光谱及其红边特征(Ⅱ)[J].棉花学报,2003,15(4):215-220.
[20]Takebe M,Yoneyama T,Inada K,Murakami T.Spectral reflectance ratio of rice canopy for estimating crop nitrogen status plant and soil[J].Plant and Soil,1990,122(2):295-297.
[21]王秀珍,王人潮,黄敬峰.微分光谱遥感及其在水稻农学参数测定上的应用研究[J].农业工程学报,2002,18(1):9-14.
[22]Shibayama M,Akiyama T.Estimating grain ield of maturing rice canopies using high spectral resolution reflectance measurements[J].Remote Sensing of Environment,1991,36(1):45-53.
[23]唐延林,王纪华,黄敬峰,王人潮,何秋霞.水稻成熟过程中高光谱与叶绿素、类胡萝卜素的变化规律研究[J].农业工程学报,2003,19(6):167-173.
[24]乔红波.麦蚜、白粉病危害后冬小麦冠层高光谱的测量与分析[D].北京:中国农业科学院,2004:17-32.
[25]黄木易.冬小麦条锈病害的高光谱遥感监测[D].合肥:安徽农业大学,2004:62-63.
[26]王恋恋.干旱区山地垂直带草地资源类型解译方法的探讨[D].乌鲁木齐:新疆农业大学,2013.
[27]Tsai F,Philpot W.Derivative analysis of hyperspectral data[J].Remote Sensing of Environment,1998,66(1):41-51.
[28]姚付启,张振华,杨润亚,孙金伟,崔素芳.基于红边参数的植被叶绿素含量高光谱估算模型[J].农业工程学报,2009,(S2):123-129.
[29]屈冉,李双,徐新良,王昌佐,通拉嘎.草地退化杂类草入侵遥感监测方法研究进展[J].地球信息科学学报,2013,15(5):761-767.
猜你喜欢
石油和化工设备(2022年3期)2022-07-13
数学物理学报(2021年2期)2021-06-09
中学数学研究(广东)(2019年21期)2019-12-16
数学物理学报(2019年5期)2019-11-29
新教育论坛(2019年35期)2019-09-10
现代机械(2018年1期)2018-04-17
散文诗(2017年15期)2018-01-19
连环画报(2017年1期)2017-07-13
广东技术师范大学学报(2016年5期)2016-08-22
大众考古(2015年1期)2015-06-26
