
内皮素3在不同毛色绵羊皮肤的表达与定位
2015-03-22 12:45李亚楠赵兵令马淑慧赫晓燕范瑞文王海东耿建军董常生
畜牧兽医学报 2015年12期
李亚楠,赵兵令,马淑慧,赫晓燕,范瑞文,王海东,耿建军,董常生*
(1.山西农业大学动物科技学院,太谷 030801;2.中国农业大学动物医学院,北京 100193)
内皮素3在不同毛色绵羊皮肤的表达与定位
李亚楠1,赵兵令1,马淑慧2,赫晓燕1,范瑞文1,王海东1,耿建军1,董常生1*
(1.山西农业大学动物科技学院,太谷 030801;2.中国农业大学动物医学院,北京 100193)
旨在探索内皮素3(Endothelin 3,EDN3)在不同毛色绵羊皮肤的表达差异以及对毛色的影响。以不同毛色绵羊为研究对象,通过荧光定量PCR技术分析EDN3基因在不同毛色绵羊皮肤的表达量,通过免疫组织化学和ELISA方法对EDN3蛋白在不同毛色绵羊皮肤中的定位和表达进行研究。1)qRT-PCR结果显示,EDN3 在白色绵羊皮肤的相对表达量为5.540 6±0.030 7,在黑色绵羊皮肤的相对表达量为1.005 8±0.062 0。黑白花色绵羊的白色区和黑色区的相对表达量分别为4.341 9±0.087 7和1.150 1±0.005 3。2)ELISA结果显示,EDN3在白色绵羊皮肤中的表达量为128.424 7±2.223 4,在黑色绵羊皮肤的相对表达量为25.114 4±3.248 3。在黑白花色绵羊白色皮肤中的表达量为93.945 3±7.562 2,黑色皮肤的表达量为28.606 0±9.295 9。3)免疫组织化学显示,EDN3在绵羊皮肤毛囊的毛基质、内外毛根鞘、毛乳头等区域均有表达。综上表明,EDN3在不同毛色绵羊皮肤中均可正常表达,且表达量存在显著差异。EDN3是维持黑色素细胞存在不可缺少的基因,但不影响绵羊黑白花的形成。
内皮素3;绵羊;免疫组织化学
哺乳动物被毛的生长是极其复杂的生理过程,其中毛色的形成更是受到遗传、环境、营养和代谢水平等多种因素的影响[1],且遗传基因的调控是其决定性的因素。在动物毛色形成过程中所涉及的调控基因多达上百个[2],例如MC1R、MITF、TRP1、TRP2、EDN3等。内皮素(Endothelin,EDN)由内皮细胞合成,有3种形态的异构体(EDN1,EDN2,EDN3),均由21个氨基酸组成[3],是一类具有强大的促分化和促有丝分裂的生长因子,广泛作用于包括皮肤在内的大部分组织。目前已知的内皮素受体有A和B两种亚型,均属于G蛋白偶联受体家族。两种受体分布的位置不尽相同,对内皮素的亲和力也存在差异,EDN3是近来研究较多的一种活性肽,对黑色素细胞的生存、发育以及色素的沉积有着不可或缺的作用。EDN3主要是通过与内皮缩血管肽B型受体(EDNRB)结合,对于由神经嵴细胞发育而来的黑色素细胞的成熟及分化发挥至关重要的作用[4],是其有效的促进因子。已有研究表明,EDN3对小鼠的黑色素细胞色素沉积有明显作用,因而进一步明确EDN3对不同毛色皮肤的黑色素沉积的影响作用可以对毛色的相关研究提供新的研究依据。目前有研究表明,内源性的EDN3在胚胎形成的早期诱导神经嵴细胞的正常迁移和分化[5-7],随着胚胎的发育,EDN3对真皮黑色素细胞的正常维持也是必不可少的[8];同时还可在后期的发育中代偿性的补充Kit的作用[9],对早期色素的形成有至关重要的作用。而外源性的EDN3不仅对黑色素细胞前体的分化有明显的促进作用,还能影响已分化的黑色素细胞的分裂增殖,最终可产生一种类似于人类黑色素细胞增多症的表型。在以角化驱动的小鼠为模型的研究中,EDN3可诱导成年小鼠皮肤真皮层出现色素沉着[8]。前期的试验大多以黑色素细胞为主要研究对象阐述EDN3对其产生的影响,对于EDN3与色素形成的关系研究较少,且对于EDN3是否会影响绵羊黑白花的形成少有提及,其与纯色绵羊是否存在差异?本研究主要通过对不同毛色的绵羊皮肤中的EDN3进行检测,以探究EDN3对绵羊毛色的影响及其与绵羊黑白花形成的关系。
1 材料与方法
1.1 试剂
RNA提取试剂盒,反转录试剂盒( TaKaRa 公司),QuantiFast SYBR Green PCR Kit( QIAGEN 公司),绵羊内皮素3 ELISA 试剂盒(上海西塘生物科技公司进口),DAB显色试剂盒(康为世纪公司),EDN3抗体(博奥森公司),Streptavidin-HRP试剂盒(康为世纪公司)。
1.2 材料
随机挑取1岁左右小尾寒羊全白、全黑、花黑绵羊各3只(由浑源羊场提供),背部皮肤净毛后,用取皮器取约1 cm的皮肤组织3块,其中2块快速置于液氮保存,用于提取总RNA及总蛋白,1块置于Bouin’s液中固定,制作石蜡切片,用于免疫组化染色。
1.3 方法
1.3.1 RNA提取和cDNA合成 从液氮中取出皮肤于液氮中研磨成粉末,Trizol 法提取绵羊皮肤总 RNA,并电泳检测其完整性。按照 TaKaRa公司反转录试剂盒进行 cDNA 合成。反转录体系:oligdT Primer 1 μL,dNTP Mix 1 μL,总 RNA 浓度<5 μg,加水至 10 μL。体系混匀后置于 PCR 仪中,按条件65 ℃ 5 min,冰上2 min进行反应;取上述产物10 μL,5×Primer script buffer 4 μL,RNase Inhibitor 0.5 μL,Primer script II RTase 1 μL,加水至20 μL。混匀后,置于 PCR 仪中按 45 ℃ 40 min,70 ℃ 15 min进行反应,-20 ℃保存 cDNA备用。
1.3.2 引物设计及PCR反应 根据GenBank 上绵羊的EDN3 序列并利用 Premier5.0 引物设计荧光定量 PCR 扩增引物,并通过 NCBI 初步检测引物的特异性,引物由北京华大基因公司合成。引物序列见表1。
PCR反应体系(25 μL):2×Taq Master Mix 12.5 μL,特异性上下游引物各1 μL,cDNA≤1 μg,加无菌水补足25 μL。反应条件:94 ℃ 预变性5 min;94 ℃ 变性30 s,60 ℃ 退火30 s,72 ℃延伸30 s,35个循环;72 ℃延伸5 min.,利用未加模板(以灭菌蒸馏水代替)的反应液作为阴性对照,以检查是否存在污染。用 1% 琼脂糖凝胶电泳检测 PCR 产物,200 V 电泳15 min 后使用凝胶成像系统采集图像。PCR产物纯化后送北京华大基因生物公司双向测序。
表1 目的基因引物序列及PCR产物
Table 1 Sequence of primer and condition of PCR amplification

基因Gene引物序列(5′→3′)PrimersequencePCR产物/bpProductsizeEDN3F:GAGCAGGGACCAAGTCAGTT,R:ATAGGGCACAGTCCGTTCAG16218SF:GAAGGGCACCACCAGGAGT,R:CAGACAAATCACTCCACCAA158
1.3.3 qRT-PCR 扩增 荧光定量 PCR 根据 QIAGEN 试剂盒进行,反应体系(10 μL):2×QuantiFast SYBR Green PCR Master Mix 5 μL,上下游引物各1 μL,模版≤100 ng,加水至10 μL。反应条件:95 ℃预变性5 min;95 ℃ 10 s,60 ℃ 30 s,40~45个循环。反应结束通过2-△△CT法计算EDN3在不同毛色绵羊皮肤中的相对表达水平。△CT目的基因=CT目的基因-CT内参基因,△△CT = △CT白色绵羊组-△CT黑色绵羊组,EDN3 mRNA表达差别倍数以2-△△CT表示。
1.3.4 ELISA 蛋白含量测定 取液氮冻存的绵羊皮肤组织,用总蛋白提取试剂盒提取总蛋白。根据ELISA试剂盒对皮肤组织中EDN3的浓度进行测定。将蛋白样品加入包被过绵羊EDN3单抗的酶标板上,并于37 ℃孵育40 min,反应完成后,甩去多余的液体,并用洗涤液充分清洗30 s×5次;每孔加入蒸馏水和一抗工作液各50 μL(空白除外),混匀,37 ℃放置20 min;反应完成后甩去多余的液体并用洗涤液充分清洗30 s×5次;每孔加入酶标抗体工作液100 μL,混匀放置37 ℃ 10 min;反应完成后,甩去多余的液体,并用洗涤液充分清洗30 s×5次;每孔加入底物工作液100 μL,置于37 ℃暗处反应15 min;每孔加入终止液100 μL混匀,用酶标仪在450 nm处测吸光值。
1.3.5 免疫组织化学 石蜡切片脱蜡,滴加内源性过氧化物酶封闭液;置37 ℃烘箱10 min,去除内源性过氧化物酶,PBS 缓冲液摇洗切片5 min×3 次;滴加适量正常羊血清封闭,37 ℃温箱孵育10 min,甩去多余液体,并滴加 1∶100 兔抗EDN3多克隆抗体,阴性对照滴加PBS缓冲液代替;4 ℃过夜;次日室温放置30 min,PBS缓冲液冲洗5 min×3 次;滴加生物素标记羊抗兔二抗工作液,放入37 ℃温箱反应10 min,用 PBS缓冲液冲洗5 min×3次;滴加HRP标记的链霉亲和素,放入37 ℃温箱反应10 min,用PBS 缓冲液冲洗5 min×3次;滴加DAB显色10 min,PBS 缓冲液冲洗3 min×3次,苏木素轻度复染、脱水、透明,中性树胶封片,显微镜下观察。
1.3.6 免疫组织化学图像数据分析 用DP软件在每张毛囊组织切片上,采集5~8个视野,应用Image-pro plus 6.0软件(美国MediaCybernetics公司) 对不同毛色绵羊皮肤EDN3免疫组织化学结果进行分析,测得阳性细胞的IOD(Integral optical density)值,单因素方差分析,结果用“平均值±标准误(Mean±SE)”表示。
2 结 果
2.1 PCR扩增电泳检测
1%凝胶电泳结果显示,EDN3条带清晰,无非特异性条带。条带大小为162 bp左右(图1)。切胶后送华大基因测序,结果比对后正确,说明EDN3可在绵羊体内正常表达。

M.DL2000 DNA marker;1.全黑绵羊皮肤EDN3 PCR产物;2.花黑绵羊皮肤EDN3 PCR产物;3.全白绵羊皮肤EDN3 PCR产物;4.花白绵羊皮肤EDN3 PCR产物M.DL2000 DNA marker;1.EDN3 PCR product of black sheep skin;2.EDN3 PCR product of flower black sheep skin;3.EDN3 PCR product of white sheep skin;4.EDN3 PCR product of flower white sheep skin图1 EDN3 PCR产物电泳图Fig.1 Electrophoresis pattern of EDN3 PCR product
2.2 qRT-PCR的扩增
qRT-PCR结果显示,目的基因与内参基因的熔解曲线仅有一个明显的峰,扩增产物的Tm值均一,目的基因及内参基因没有产生非特异性扩增及引物二聚体(图2A、B);且EDN3荧光定量的扩增动力学曲线为“S”型,曲线平行性良好,无明显偏差,起峰点清晰,符合标准曲线要求(图2C)。qRT-PCR 结果显示,EDN3 在全白色绵羊皮肤中 mRNA 的相对表达量为5.540 6±0.030 7;而在黑白花绵羊的白色皮肤中EDN3 的 mRNA 相对表达量为4.341 9±0.087 7,黑白花绵羊的黑色皮肤中mRNA的相对表达量为1.150 1±0.005 3;在全黑色绵羊皮肤的相对表达量为1.005 8±0.062 0。全白绵羊皮肤中EDN3的表达量是黑白花绵羊黑色皮肤的4.8倍,是全黑绵羊皮肤表达量的5.5倍;黑白花绵羊白色皮肤EDN3的表达量是黑白花绵羊黑色皮肤的3.8倍,是全黑绵羊皮肤的4.3倍。白色绵羊皮肤与黑色绵羊皮肤的EDN3 mRNA水平的表达差异极显著(P<0.01)(图2D)。
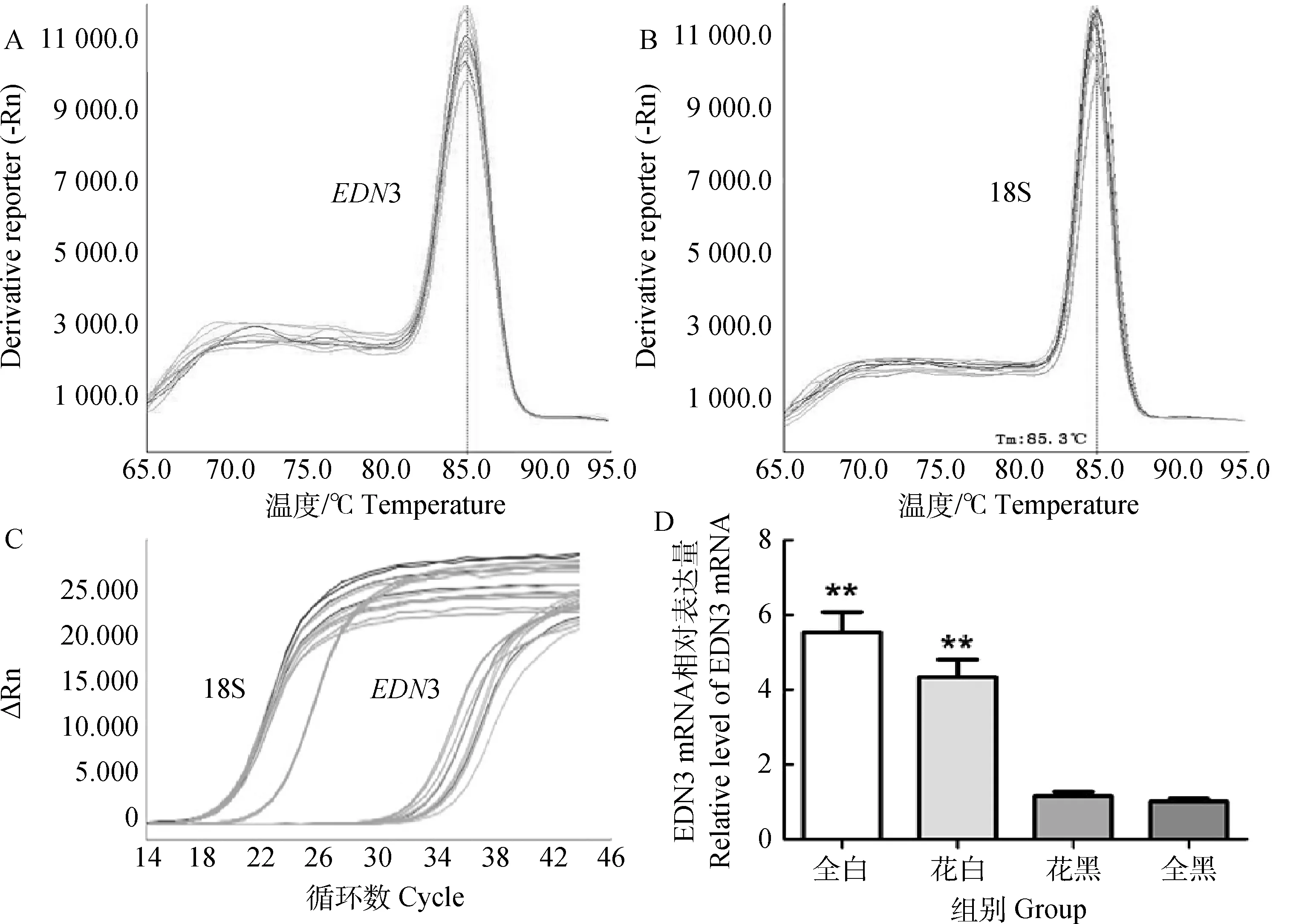
**.P<0.01。下同。A.EDN3的熔解曲线;B.18S的熔解曲线;C.EDN3和18S的扩增曲线;D.不同绵羊皮肤EDN3的qRT-PCR分析结果**.P<0.01.The same as below.A.qRT-PCR melt curve for EDN3;B.qRT-PCR melt curve for 18S;C.qRT-PCR amplification plots for EDN3 and 18S;D.qRT-PCR analysis of EDN3 in different colors of sheep skin图2 EDN3 mRNA表达的qRT-PCR分析Fig.2 qRT-PCR analysis of the expression of EDN3 mRNA
2.3 ELISA试验
经酶标仪在450 nm处测得OD值,使用CurveExpert 1.4软件分析制作标准曲线(图3),获得样品中EDN3蛋白的相对表达量。EDN3在全白色绵羊皮肤中蛋白的相对表达量为128.424 7±2.223 4;而在黑白花绵羊的白色皮肤中EDN3蛋白相对表达量为93.945 3±7.562 2,黑白花绵羊的黑色皮肤中蛋白的相对表达量为28.606 0±9.295 9;在全黑色绵羊皮肤的相对表达量为25.114 4±3.248 3。全白绵羊皮肤中EDN3蛋白表达量是黑白花绵羊黑色皮肤的4.5倍,是全黑绵羊皮肤表达量的5.1倍;黑白花绵羊白色皮肤EDN3蛋白表达量是黑色皮肤的3.3倍,是全黑绵羊皮肤的3.7倍。白色绵羊皮肤与黑色绵羊皮肤EDN3蛋白水平表达差异极显著(P<0.01)。
2.4 免疫组织化学分析
免疫组织化学结果显示(图4),绵羊黑色皮肤和白色皮肤中均有EDN3棕黄色的阳性表达,且其主要表达部位为毛基质、内外毛根鞘等处,着色深浅不一。另黑色绵羊毛囊的毛乳头区有少量表达,白色皮肤则比黑色的表达更少,几乎不可见。通过光密度分析比较EDN3在不同毛色绵羊皮肤的表达水平,阳性试验组结果表明(图5),白色皮肤EDN3的表达量高于黑色皮肤且差异极显著(P<0.01),花黑EDN3表达量高于全黑,差异显著(P<0.05),该结果与ELISA试验结果相一致。
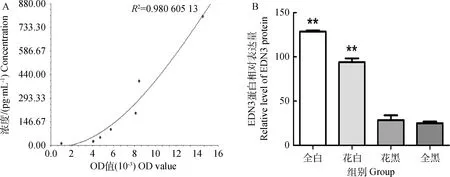
A.EDN3的标准曲线;B.EDN3蛋白表达量的ELISA分析A.The standard curve of EDN3;B.ELISA analysis of EDN3 in different colors of sheep skin图3 不同毛色绵羊皮肤中EDN3蛋白的相对表达量Fig.3 Relative expression of EDN3 in different hair color sheep skin
A.白色绵羊皮肤EDN3阳性组;a.白色绵羊皮肤EDN3阴性对照组;B.花白绵羊皮肤EDN3阳性组;b.花白绵羊皮肤EDN3阴性对照组;C.花黑绵羊皮肤EDN3阳性组;c.花黑绵羊皮肤EDN3阴性对照组;D.黑色绵羊皮肤EDN3阳性组;d.黑色绵羊皮肤EDN3阴性对照组。1.毛基质;2.内毛根鞘;3.外毛根鞘;4.毛乳头A.White sheep skin EDN3 positive group;a.White sheep skin EDN3 negative control group;B.Flower white sheep skin EDN3 positive group;b.Flower white sheep skin EDN3 negative control group;C.Flower black sheep skin EDN3 positive group;c.Flower black sheep skin EDN3 negative control group;D.Black sheep skin EDN3 positive group;d.Black sheep skin EDN3 negative control group.1.Hair follicle matrix;2.Inside root sheath;3.Outside root sheath;4.Dermal papilla图4 EDN3在不同毛色绵羊毛囊的免疫定位200×Fig.4 Immunohistochemistry results of EDN3 in different hair color sheep skin 200×
3 讨 论
*.P<0.05图5 EDN3在不同毛色绵羊皮肤组织中的平均光密度分析Fig.5 Average optical density analysis of EDN3 in different hair color sheep skin
哺乳动物毛色的形成取决于成熟黑素细胞的相对数量、分布部位以及其合成的黑色素的含量、种类、分布和运输途径等因素。黑色素细胞来源于神经嵴细胞[10],在黑色素小体中合成黑色素,并由其决定哺乳动物皮肤、毛发及眼睛的最终表型[11]。内皮素(Endothelin,EDN)由内皮细胞合成,有3种形态的异构体(EDN1,EDN2,EDN3)。EDN通过与G蛋白偶联受体结合激活磷脂酶C(PLC),诱发细胞内钙离子浓度的增加进而参与各种生理过程[12],主要包括血管收缩和细胞增殖[13]。已有研究表明,EDN3在鸟类神经嵴细胞的培养中可以通过与EDNRB结合促进神经嵴细胞的增殖和黑色素细胞的分化[14]。在黑色素通路中,EDN3与KIT共同参与黑色细胞的存活、增殖和分化等,而同时阻断EDNRB和KIT信号则会导致黑色素前体细胞完全消失,任一通路的存在均可维持黑色素前体细胞的存在[9]。A.G.Baynash等[15]通过细胞试验发现EDN3及其受体在神经嵴细胞和黑色素细胞的增殖和分化中起重要的作用,并推测人类EDN3缺失可能会导致神经性巨结肠病症。另有报道称KIT受体酪氨酸激酶W位点的突变不影响黑色素细胞的迁移和分化,但是严重影响黑色素细胞的存在[16]。体外研究表明,黑色素前体细胞的增殖和分化对EDN3活性肽存在一定的剂量依赖[17-19]。本试验蛋白定位和定量结果显示,EDN3在不同毛色皮肤毛囊内均有表达,但是白色皮肤表达量明显多于黑色。不同毛色皮肤定位图所显示的阳性反应区也存在一些差异,白色与花白色皮肤定位显示阳性反应的区域均匀分布于内外毛根鞘和毛基质等处,而在毛乳头区仅有少量存在。黑色与花黑色皮肤则显示EDN3阳性反应在毛基质等黑色素细胞活跃的区域大量存在,毛乳头区的阳性反应也有明显的增多。试验结果提示,EDN3对黑色与花黑色绵羊皮肤内的黑色素细胞有一定的促进作用,活化细胞增加色素的合成。EDN3对白色及花白色绵羊皮肤毛囊内的黑色素细胞有维持作用,即EDN3对黑色素细胞的生存是必不可少的,白色绵羊皮肤需要EDN3较高的表达量以维持色素细胞的存在。杨姗姗等[20]认为EDN3可能与绵羊毛色形成具有相关性,但从本试验结果推测EDN3可能与维持黑色素细胞活性及其发育有较大关系,在色素颗粒形成上具有协调作用。同时由于存在样品采集时对绵羊品种和年龄等选择上的不一致,均可能导致结果存在差异。
EDN3与EDNRB任一位点的隐形突变会产生相似的表型,包括不同程度的色素减退和无神经节的巨结肠疾病,其中色素减退的表型是胚胎期黑色素细胞的减少和随后黑色素细胞的异常迁移所造成。R.J.Garcia等[8]成功得到内皮素3过表达转基因小鼠模型,结果显示转基因小鼠胚胎早期EDN3的表达对表型的形成非常重要,黑色素前体细胞的数量显著增多,成年转基因鼠的真皮层已分化的黑色素细胞数量增多,且成年鼠皮肤色素沉着的表型需要内皮素3的持续表达来维持。该转基因鼠与Agouti、KIT等基因的突变体小鼠杂交可改变其原有表型。有研究显示,EDN3和EDNRB途径对黑色素前体细胞和已分化成熟的黑色素细胞都有促进作用。但是也有研究表明,毛发与皮肤中的黑色素细胞即使在分享或共用同一信号通路时,表型颜色的形成也是相互独立的[21]。EDN3在成年绵羊皮肤中存在差异,且在白色绵羊皮肤内表达量较高,说明在成年绵羊中EDN3并未直接作用于色素的生成,后期毛发颜色形成时受到其他路径的调控,影响表型的形成。C.B.Kaelin等[22]对猫科动物花纹表型的形成做了相关研究,发现EDN3在花纹间的表达存在差异,并进一步证实EDN3对早期花纹的形成有一定的影响,且作为第二信使诱导胚胎时期黑色素细胞增多,对后期花纹的形成有明显的异化作用。同时他们还证实EDN3在相邻花纹区域维持高低不同的表达进而维持毛发空间的差异,这一差异始终贯穿于毛囊的发育周期。而表型的变化则归因于EDN3引起的色素颗粒类型的变化,即真黑素向褐黑素的转变。该研究说明EDN3对褐黑素的生成和颗粒的沉积有一定的影响并对花纹的形成有促进作用,但是本试验结果却显示白色与花白色绵羊皮肤内EDN3的表达量均显著高于花黑色绵羊皮肤的表达量,与其研究结果不符,表明EDN3不是影响绵羊皮肤黑白花形成的主要基因,影响绵羊皮肤黑白花形成的因素还有待进一步研究。试验结果显示真黑素大量聚集的黑色绵羊皮肤内,EDN3的存在较少,表明EDN3与真黑素及褐黑素之间存在一定的协调相关性。
4 结 论
本研究结果表明,EDN3在不同毛色绵羊皮肤中均可正常表达,且存在显著差异。EDN3是维持黑色素细胞存在不可缺少的基因,但不影响绵羊黑白花的形成。
[1] SLOMINSKI A,TOBIN D J,SHIBAHARA S,et al.Melanin pigmentation in mammalian skin and its hormonal regulation[J].PhysiolRev,2004,84(4):1155-1228.[2] WILCZEK A,KONDH H,MISHIMA Y.Composition of mammalian eumelanins analyses of DHICA-derived units in pigments from hair and melanoma cells[J].PigmentCellRes,1996,9(2):63-67.
[3] INOUE A,YANAGISAWA M,KIMURA S,et al.The human endothelin family;three structurally and pharmacologically distinct isopeptides predicted by three separate genes[J].ProcNatlAcadSciUSA,1989,86(8):2863-2867.
[4] LAHAV R,DUPIN E,LECOIN L,et al.Endothelin 3 selectively promotes survival and proliferation of neural crest-derived glial and melanocytic precursorsinvitro[J].ProcNatlAcadSciUSA,1998,95(24):14214-14219.
[5] ARAI H,HORI S,ARAMORI I,et al.Cloning and expression of a cDNA encoding an endothelin receptor[J].Nature,1990,348(6303):730-732.
[6] SAKAMOTO A,YANAGISAWA M,SAWAMURA T,et al.Distinct subdomains of human endothelin receptors determine their selectivity to endothelinA-selective antagonist and endothelin B-selective agonists[J].JBiolChem,1993,268(12):8547-8553.
[7] LEE H O,LEVORSE J M,SHIN M K.The endothelin receptor-B is required for the migration of neural crest-derived melanocyte and enteric neuron precursors[J].DevBiol,2003,259(1):162-175.
[8] GARCIA R J,ITTAH A,MIRABAL S,et al.Endothelin 3 induces skin pigmentation in a keratin-driven inducible mouse model[J].JInvestDermatol,2008,128(1):131-142.
[9] AOKI H,MOTOHASHI T,YOSHIMURA N,et al.Cooperative and indispensable roles of endothelin 3 and KIT signalings in melanocyte development[J].DevDyn,2005,233(2):407-417.
[10] THOMAS A J,ERICKSON C A.The making of a melanocyte:the specification of melanoblasts from the neural crest[J].PigmentCellMelanomaRes,2008,21(6):598-610.
[11] ABDEL-MALEK Z,SWOPE V B,SUZUKI I,et al.Mitogenic and melanogenic stimulation of normal human melanocytes by melanotropic peptides[J].ProcNatlAcadSciUSA,1995,92(5):1789-1793.
[12] CLOUTHIER D E,HOSODA K,RICHARDSON J A,et al.Cranial and cardiac neural crest defects in endothelin-A receptor-deficient mice[J].Development,1998,125(5):813-824.[13] KEDZIERSKI R M,YANAGISAWA M.Endothelin system:the double-edged sword in health and disease[J].AnnuRevPharmacolToxicol,2001,41:851-876.[14] LAHAV R,DUPIN E,LECOIN L,et al.Endothelin 3 selectively promotes survival and proliferation of neural crest-derived glial and melanocytic precursorsinvitro[J].ProcNatlAcadSciUSA,1998,95(24):14214-14219.
[15] BAYNASH A G,HOSODA K,GIAID A,et al.Interaction of endothelin-3 with endothelin-B receptor is essential for development of epidermal melanocytes and enteric neurons[J].Cell,1994,79(7):1277-1285.
[16] CABLE J,JACKSON I J,STEEL K P.Mutations at the W locus affect survival of neural crest-derived melanocytes in the mouse[J].MechDev,1995,50(2-3):139-150.
[17] LAHAV R,ZILLER C,DUPIN E,et al.Endothelin 3 promotes neural crest cell proliferation and mediates a vast increase in melanocyte number in culture[J].ProcNatlAcadSciUSA,1996,93(9):3892-3897.
[18] REID K,TURNLEY A M,MAXWELL G D,et al.Multiple roles for endothelin in melanocyte development:regulation of progenitor number and stimulation of differentiation[J].Development,1996,122(12):3911-3919.
[19] OPDECAMP K,KOS L,ARNHEITER H,et al.Endothelin signalling in the development of neural crest-derived melanocytes[J].BiochemCellBiol,1998,76(6):1093-1099.
[20] 杨姗姗,白俊明,范瑞文,等.内皮素3在不同毛色绵羊皮肤中的差异表达[J].畜牧兽医学报,2015,46(11):2111-2116. YANG S S,BAI J M,FAN R W,et al.The different expression of endotholin 3 in sheep skins[J].ActaVeterinariaetZootechnicaSinica,2015,46(11):2111-2116.(in Chinese)
[21] VAN RAAMSDONK C D,BARSH G S,WAKAMATSU K,et al.Independent regulation of hair and skin color by two G protein-coupled pathways[J].PigmentCellMelanomaRes,2009,22(6):819-826.
[22] KAELIN C B,XU X,HONG L Z,et al.Specifying and sustaining pigmentation patterns in domestic and wild cats[J].Science,2012,337(6101):1536-1541.
(编辑 程金华)
EDN3 Expression and Localization in Sheep Skin with Different Coat Color
LI Ya-nan1,ZHAO Bing-ling1,MA Shu-hui2,HE Xiao-yan1,FAN Rui-wen1WANG Hai-dong1,GENG Jian-jun1,DONG Chang-sheng1*
(1.CollegeofAnimalScienceandVeterinaryMedicine,ShanxiAgriculturalUniversity,Taigu030801,China;2.CollegeofVeterinaryMedicine,ChinaAgriculturalUniversity,Beijing100193,China)
To explore the expression of endothelin 3 in different coat color of sheep skin and the effect ofEDN3 on the sheep coat color,the experiment analyzed the mRNA and protein expression level ofEDN3 in sheep skin of different coat colors in different coat color ship by real-time quantitative PCR,ELISA and immunohistochemistry.1)qRT-PCR showed that the relative expressive ofEDN3 mRNA in white sheep skin was 5.540 6±0.030 7,while in black was 1.005 8±0.062 0.The gene expression quantity ofEDN3 in white and black skin tissue of the same individuality was 4.341 9±0.087 7 and 1.150 1±0.005 3.2)ELISA results showed that the expression ofEDN3 protein in white and black were 128.424 7±2.223 4 and 25.114 4±3.248 3.The protein expression ofEDN3 in white and black skin of the same individuality were 93.945 3±7.562 2 and 28.606 0±9.295 9.3)TheEDN3 was located in the matrix,outer root sheath and dermal papilla of hair follicular by immunohischemistry.Experiments showed that theEDN3 expressed normally in black and white sheep skin,and the expression quantity had significant differences.The research indicated thatEDN3 was indispensable to maintain the presence of the melanocyte,and didn’t affect the formation of friesian sheep.
endothelin-3;sheep;immunohistochemistry
10.11843/j.issn.0366-6964.2015.12.026
2014-03-12
国家“863”计划(2013AA102506);国家自然科学基金项目(31302049;31172283);山西省自然科学基金项目(2012011046-6);高校博士点基金(20131403120002)
李亚楠(1989-),女,河南扶沟人,硕士生,主要从事毛色相关研究,E-mail:18404966095@163.com
*通信作者:董常生,教授,E-mail:csdong18@163.com
S826.2
A
0366-6964(2015)12-2322-07
猜你喜欢
养猪(2021年4期)2021-08-26
阅读与作文(小学低年级版)(2019年12期)2019-12-26
奥秘(创新大赛)(2019年8期)2019-11-30
小天使·四年级语数英综合(2019年9期)2019-11-09
畜牧兽医科学(2018年14期)2018-02-14
中华老年口腔医学杂志(2016年4期)2017-01-15
天然产物研究与开发(2016年11期)2016-06-15
中西医结合心血管病杂志(电子版)(2015年13期)2015-10-14
畜牧兽医学报(2015年3期)2015-07-05
现代检验医学杂志(2015年1期)2015-02-06
