
康氏木霉内切β—葡聚糖苷酶的分离纯化及酶学性质
2015-04-17 21:06覃益民张静茹叶锦等
江苏农业科学 2015年1期
关键词:分离纯化
覃益民 张静茹 叶锦+ 等
摘要:康氏木霉GIMP3.444经摇瓶发酵,获得分泌到胞外的纤维素酶,粗酶液经(NH4)2SO4分级沉淀、Sephacryl S200凝胶过滤层析、DEAE Sepharose FF离子交换层析及Octrl Separose CL-4B疏水层析分离纯化出一种内切型的β-葡聚糖苷酶,SDS-PAGE电泳鉴定为单一条带,BIO-RAD分析相对分子量为61.8 ku。 该内切β-葡聚糖苷酶的最适反应温度为55 ℃,最适反应pH值为4.4,作用于羧甲基纤维素钠时,Lineweaver-Burk法求得Km、Vmax分别为481 mg/mL、2.48 mg/(min·mL)。
关键词:康氏木霉;内切β-葡聚糖苷酶;分离纯化;酶学性质
中图分类号: Q556+.2文献标志码: A文章编号:1002-1302(2015)01-0040-04
收稿日期:2014-03-27
项目基金:国家自然科学基金(编号:21276053);广西壮族自治区生物炼制重点实验室培育基地开放课题(编号:GXBF11-04)。
作者简介:覃益民(1962—),男,广西贵港人,博士,副教授,主要从事生物加工与分离方面的研究。Tel:(0771)3233718;E-mail: qym6289@sina.com。植物纤维由纤维素、半纤维素和木质素三大部分组成并共同构成了植物细胞壁的主要成分,是最丰富的天然可再生生物资源,纤维素通过纤维素酶水解转化为可发酵性糖,继而转化为燃料乙醇及相关化学品的研究越来越引起人们的关注[1]。纤维素酶是一类能够将纤维素降解为葡萄糖的多组分酶系的总称,由内切β-葡聚糖苷酶 (EG) 、 外切β-葡聚糖苷酶 (CBH)和 β- 葡萄糖苷酶 3 类酶组成。尽管大量研究已表明,只有在这3种主要成分酶的协同作用下,纤维素分子最终才能被降解成葡萄糖。但是各成分酶是如何协同起作用的,学者们的看法还不尽相同[2],近来人们还发现了一些不具有纤维素酶活性的蛋白也能与纤维素酶起协同作用,并最终促进纤维素的降解[3-5]。因此,无论是对纤维素酶系各成分酶的协同作用机理,还是对纤维素酶系与其他非酶蛋白的协同增效作用的更深入了解,都将会为提高纤维素酶的催化效率提供重要理论基础。康氏木霉GIMP 3.444是一株产纤维素酶系的真菌菌株[6],笔者从该菌株中分离纯化出一种纤维素酶协同增效蛋白。为进一步研究该增效蛋白对纤维素酶各成分酶的协同作用机理,需获得该菌株产的纤维素酶系各组分纯酶。因此,本研究对康氏木霉GIMP3.444分泌的内切β-葡聚糖苷酶进行分离纯化,并研究其酶学性质。
1材料与方法
1.1菌株
康氏木霉(Trichoderma koningii)GIMP 3.444购于广东省微生物菌种保藏中心,现保存于广西大学化学化工学院生物化工实验室。
1.2培养基和试剂
(1)斜面活化培养基:200.0 g马铃薯、20.0 g 葡萄糖、30 g KH2PO4、1.5 g MgSO4·7H2O、20.0 g 琼脂粉,加自来水至1 000 mL,调 pH值至 6.0,121 ℃灭菌 20 m in。(2)摇瓶种子培养基:200.0 g 马铃薯、20.0 g 葡萄糖、30 g KH2PO4、1.5 g MgSO4·7H2O,加自来水至1 000 mL,调pH值至 6.0,121 ℃灭菌 20 m in。(3)液体发酵培养基:15.0 g 甘蔗渣(过80目筛)、8.0 g 麦麸粉、4.0 g (NH4)2SO4 、3.0 g KH2PO4 、1.5 g MgSO4·7H2O,加自来水至1 000 mL,调pH值至6.0,121 ℃灭菌 20 min。(4)DNS试剂:称取6.3 g 3,5-二硝基水杨酸、20.96 g NaOH、182.0 g酒石酸钾钠、5.0 g苯酚、5.0 g 亚硫酸钠,分别用去离子水溶解后混合,待混合液冷却至室温定容至 1 000 mL。保存在棕色瓶中,在室温下放置 7 d 即可使用。(5)考马斯亮蓝G-250溶液:称取考马斯亮蓝G-250 100 mg,溶于50 mL 95%乙醇中,再加入100 mL 85%磷酸,混合均匀后用去离子水定容至1 000 mL。
1.3菌种的培养
取保存的康氏木霉接种于斜面活化培养基,28 ℃条件下活化培养3~7 d,刮取平面孢子,用无菌水制成孢子悬浮液,按1%体积比(V/V)接种至种子培养液中,于180 r/min、28 ℃下培养30~36 h。将摇瓶种子按5%体积比(V/V)接种量接种至装有150 mL发酵培养基的500 mL三角瓶中,于 180 r/min、28 ℃下培养120 h。
1.4内切β-葡聚糖苷酶的分离纯化
1.4.1发酵液的预处理取“1.3”节培养好的发酵液,用4层纱布过滤,在4 ℃、6 000 r/min条件下离心20 min,收集上清液,用截留分子量为10 000 Da的VIVAFLOW 200平板超滤膜浓缩20倍左右,得到粗酶液。粗酶液先用20%饱和度的硫酸铵沉淀,4 ℃下静置过夜;6 000 r/min离心20 min,取上清液继续加入硫酸铵至饱和度为80%,4 ℃下静置过夜;离心得沉淀,用0.05 mol/L pH值7.0的磷酸二氢钠-磷酸氢二钠缓冲液复溶,再次离心后,去除不溶物,复溶酶液4 ℃冰箱保存。
1.4.2Sephacryl S-200凝胶过滤层析取2.0 mL的盐析复溶液,上样于Sephacryl S-200 HR凝胶过滤柱(10 mm×700 mm)中,用0.05 mol/L pH值 7.0的磷酸二氢钠-磷酸氢二钠缓冲液(含NaCl 0.2 mol/L)洗脱,用自动收集器按 2.0 mL/(管·10 min) 收集组分,测定每管内切β-葡聚糖酶活力,收集有内切β- 葡聚糖酶活性洗脱峰,4 ℃保存备用。
1.4.3DEAE Sepharose FF离子交换层析取“1.4.2”节收集的蛋白样,用VIVAFLOW 50平板超滤膜浓缩,并用 0.05 mol/L pH值 7.0磷酸二氢钠-磷酸氢二钠缓冲液置换,上样至DEAE Sepharose FF阴离子交换柱(16 mm×200 mm)中,0~0.8 mol/L NaCl浓度梯度洗脱,自动收集器 5 mL/(管·5min) 收集组分,测定各峰的内切β-葡聚糖酶活力,收集内切β- 葡聚糖酶活性峰,4 ℃条件保存备用。
1.4.4Octrl Separose CL-4B疏水层析取“1.4.3”节中收集的内切β-葡聚糖酶活性峰,超滤浓缩并用含(NH4)2SO4 2.0 mol/L的0.05 mol/L pH值 7.0磷酸二氢钠-磷酸氢二钠缓冲液置换,上样至Octrl Separose CL-4B疏水层析柱(10 mm×400 mm)中,2.0~0 mol/L (NH4)2SO4 浓度梯度洗脱,自动收集器3 mL/(管·10 min) 收集组分,测定各峰的内切β- 葡聚糖酶活力,收集内切β-葡聚糖酶活性峰,4 ℃条件保存备用。
1.5测定方法
1.5.1蛋白浓度的测定以牛血清白蛋白为标准,用考马斯亮蓝G-250法[7]测蛋白液浓度。
1.5.2内切β-葡聚糖苷酶活力(EG)测定采用DNS法[8]测定内切酶活性。向试管中加入1.5 mL 10 g/L的羧甲基纤维素钠柠檬酸-柠檬酸钠溶液(0.1 mol/L,pH值 4.8)及 0.5 mL 酶液,50 ℃水浴中保温30 min,取出后加入 3.0 mL DNS试剂终止反应,沸水浴5 min,取出流水冷却,定容至 25 mL,在540 nm波长处测定吸光度。以每分钟催化底物生成1 μmol葡萄糖所需的酶量定义为一个活力单位,单位用IU/mL表示。
1.5.3蛋白质纯度鉴定及相对分子量测定采用SDS-PAGE法[9]。聚丙烯酰胺凝胶质量分数为12%,蛋白标准品的分子质量范围1.44×104~9.74×104 ku。
1.5.4最适反应温度测定将纯酶稀释后,在pH值 4.8条件下,于不同温度(25 ℃~80 ℃)水浴中与底物保温30 min,测定内切β-葡聚糖苷酶活力,以最大酶活力为100%,确定酶的最适反应温度。
1.5.5最适反应pH值测定分别用不同pH值(2.6~8.0)的磷酸氢二钠-柠檬酸缓冲液配置1%的羧甲基纤维素钠悬浮液,作为不同pH值条件下的酶作用底物,加入用相应pH值缓冲液稀释的纯酶,在50 ℃水浴中保温30 min,测定不同pH值下的内切β-葡聚糖苷酶活力,以最大酶活力为100%,确定酶的最适反应温度。
1.5.6底物动力学参数测定分别配置浓度为0.1%、015%、0.2%、0.25%、0.3%、0.35%的羧甲基纤维素钠悬浮液(pH值 4.8),加入适当稀释纯酶液,在50 ℃水浴中保温30 min,DNS法测定各不同浓度底物时产生的还原糖,然后按照Lineweaver-Burk双倒数作图法[10]作图,计算出该酶作用于羧甲基纤维素钠时的Km值和Vmax值。
2结果与分析
2.1内切β-葡聚糖苷酶的分离纯化及分析
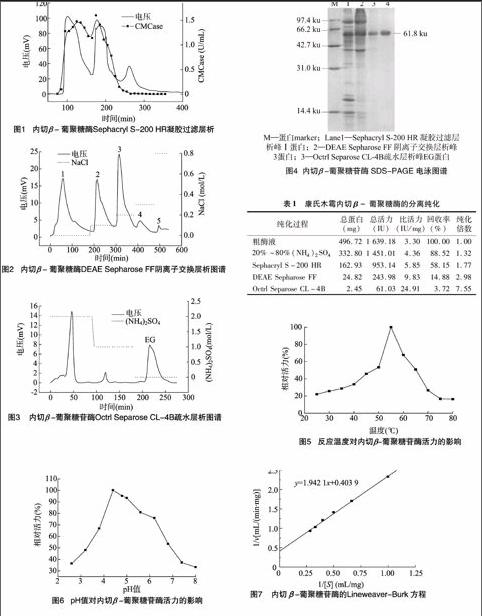
康氏木霉发酵液经20~80%硫酸铵分级沉淀,上样至Sephacryl S-200 HR凝胶过滤层析柱,洗脱流速为 0.2 mL/min,洗脱结果如图1所示,共有3个蛋白峰。酶活测定发现第3个蛋白峰没有纤维素酶活性,为杂蛋白,而峰Ⅰ、峰Ⅱ均具有较高的内切β-葡聚糖苷酶活性,说明康氏木霉的发酵液中至少含有2种内切酶。本研究仅介绍对第1个蛋白峰的分离纯化。
收集图1中的峰Ⅰ蛋白,超滤法浓缩并进行缓冲液置换后,上样至用0.05 mol/L pH值 7.0磷酸二氢钠-磷酸氢二钠缓冲液平衡的DEAE Sepharose FF离子交换层析柱,分别用3倍柱体积的含NaCl 0.1、0.2、0.3、0.8 mol/L的缓冲液进行洗脱,流速为1.0 mL/min,结果如图2所示,共有5个蛋白吸收峰,经测定内切酶活主要集中在峰3中。
合并图2峰3蛋白,超滤浓缩用含(NH4)2SO4 2.0 mol/L 的磷酸二氢钠-磷酸氢二钠缓冲液置换,上样至平衡的Octrl Separose CL-4B疏水层析柱,分别用3倍柱体积的含 1.0 mol/L (NH4)2SO4缓冲液和无(NH4)2SO4的缓冲液洗脱,流速为0.3 mL/min,如图3所示,得到3个蛋白峰,经酶活测定,发现只有第3个峰具有内切酶活性(图中记为EG)。
取经硫酸铵分级沉淀、Sephacryl S-200 HR凝胶过滤层析、DEAE Sepharose FF离子交换层析、Octrl Separose CL-4B疏水层析纯化的内切β- 葡聚糖酶进行SDS-PAGE凝胶电泳分析,结果见图4。图4表明,通过以上4步纯化过程,内切β-葡聚糖酶的纯度逐步提高,且经疏水层析后的酶组分在SDS-PAGE电泳胶上显示为单一条带,表明该组分已达电泳纯,凝胶成像系统分析内切β-葡聚糖酶的相对分子量约为61.8 ku,比文献[11-14]报道的康氏木霉内切葡聚糖酶大,也偏大于里氏木霉[15-16]内切葡聚糖酶,而与绿色木霉[17]和拟康氏木霉[18]中的内切葡聚糖酶相近。
内切β-葡聚糖酶的分离纯化过程如表1。表1表明硫酸铵分级沉淀除去了部分的非酶杂蛋白,纯度提高了132倍。而Sephacryl S-200 HR凝胶过滤层析对内切β-葡聚糖
酶的提纯效果不太理想,在盐析的基础上内切酶只被纯化了1.34倍,这是因为在盐析样中有多种内切酶共存,此时测得的内切酶活力会高于凝胶过滤层析后得到的内切β- 葡聚糖酶活力。DEAE Sepharose FF离子交换层析也能去除较多的杂蛋白和多糖物质,内切β-葡聚糖酶的比活力达到 983 IU/mL,纯化了2.98倍。经过Octrl Separose CL-4B疏水层析,内切葡聚糖酶最终达到电泳纯,此时比活力为 24.91 IU/mg,与粗酶液相比,提高了7.55倍,回收率为3.72%。
表1康氏木霉内切β- 葡聚糖酶的分离纯化
纯化过程总蛋白
(mg)总活力
(IU)比活力
(IU/mg)回收率
(%)纯化
倍数粗酶液496.721 639.183.30100.001.0020%~80%(NH4)2SO4 332.801 451.014.3688.521.32Sephacryl S-200 HR162.93953.145.8558.151.77DEAE Sepharose FF 24.82243.989.8314.882.98Octrl Separose CL-4B 2.4561.0324.913.727.55
2.2内切β-葡聚糖苷酶的酶学性质
2.2.1内切酶的最适反应温度为了测定反应温度对内切β-葡聚糖苷酶活性的影响,确定酶的最适反应温度,在同一pH值条件下,分别测定不同反应温度时的酶活力,以最大酶活力为100%,计算相对酶活力,结果如图5。图5表明,在 25~55 ℃ 范围内,内切酶活力随着温度升高而增加,在55 ℃时酶活力最大,说明内切酶的最适反应温度为55 ℃;温度继续升高,内切酶活力逐渐降低,80 ℃时,酶活力仅有最高活力的16.3%。在55~75 ℃范围内,酶活力随温度升高而降低的斜率的绝对值大于低温范围内酶活力随着温度升高而增加的斜率,说明该内切酶对高温更敏感。
2.2.2内切酶的最适反应pH值用不同pH值的缓冲液配置酶反应底物,测定pH值对内切β-葡聚糖苷酶活性的影响,以确定酶的最适反应pH值,分别将该酶与上述底物在 50 ℃ 条件下反应,测定内切β-葡聚糖苷酶活力,以最大酶活力为100%,计算相对酶活力,结果如图6。可见,当pH值小于4.4时,内切酶活力随着pH值的增大而增加,至pH值 4.4时酶活力达最大值,故该内切酶的最适最适pH值为44;pH值 4.4~8.0范围内,内切酶活力随着pH值的继续增大而降低,但在pH值为6.2时,酶活力仍有最大值的758%,这可能是因为该内切酶在pH值 4.4~6.2之间活性较稳定。
2.2.3内切酶的底物动力学性质将稀释的纯酶液分别与不同浓度底物反应,测定生成的还原糖量,然后按照Lineweaver-Burk双倒数作图法作图,结果如图7。回归得出标准方程为1/v=1.942 1(1/[S])+0.403 9,求得以羧甲基纤维素钠为底物时,内切β-葡聚糖苷酶的Km值为 4.81 mg/mL,Vmax值为2.48 mg/(min·mL)。
3结论
本研究从康氏木霉液态摇瓶发酵液中分离纯化得到一种电泳纯的内切β-葡聚糖苷酶,比活力为24.91 IU/mg,相对分子量为61.8 ku。酶学性质测定结果显示,纯化的内切β-葡聚糖苷酶无外切β-葡聚糖苷酶及β- 葡萄糖苷酶活性,作用于羧甲基纤维素钠时,Km为4.81 mg/mL,Vmax为 2.48 mg/(min·mL),最适反应温度为55 ℃,最适反应pH值为4.4,可用来分析纤维素酶系蛋白之间的协同水解作用。
本研究结果还表明,康氏木霉分泌的内切β-葡聚糖苷酶不止一种,因此,在研究纤维素酶系各成分酶的协同作用机理或研究纤维素酶系与其他非酶蛋白的协同增效作用时,须考虑同功酶存在的影响。
参考文献:
[1]Menon V,Rao M L. Trends in bioconversion of lignocellulose: Biofuels,platform chemicals and biorefinery concept[J]. Progress in Energy and Combustion Science,2012,38(4): 522-550.
[2]Van Dyk J S,Pletschke B I. A review of lignocellulose bioconversion using enzymatic hydrolysis and synergistic cooperation between enzymes-factors affecting enzymes,conversion and synergy[J]. Biotechnology Advances,2012,30(6): 1458-1480.
[3]卢迪,陈洪章,马润宇. 秸秆质外体蛋白对纤维素酶活力的影响[J]. 生物工程学报,2006,22(2):257-262.
[4]Kim E S,Lee H J,Bang W G,et al. Functional characterization of a bacterial expansin from Bacillus subtilis for enhanced enzymatic hydrolysis of cellulose[J]. Biotechnology and Bioengineering,2009,102(5): 1342-1353.
[5]Han Ye Jun,Chen Hong Zhang. Synergism between hydrophobic proteins of corn stover and cellulase in lignocellulose hydrolysis[J]. Biochemical Engineering Journal,2010,48(2): 218-224.
[6]覃益民,高进,刘幽燕,等. 一种来源于康氏木霉的纤维素酶增效蛋白的分离纯化[J]. 化工学报,2011,62(10):2893-2897.
[7]Bradford M M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding[J]. Analytical Biochemistry,1976,72(1/2): 248-254.
[8]Miller G L. Use of dinitrosalicylic acid reagent for determination of reducing sugar[J]. Analytical Biochemistry,1959,31(3): 426-428.
[9]汪家政,范明. 蛋白质技术手册[M]. 北京:科学出版社,2000:77-100.
[10]Shyamala S,Ravikumar S,Vikramathithan J,et al. Isolation,purification,and characterization of two thermostable endo-1,4-β-D-glucanase forms from Opuntia vulgaris[J]. Applied Biochemistry and Biotechnology,2011,165(7/8): 1597-1610.
[11]Wood T M,Mccrae S I. The cellulase of Trichoderma koningii. Purification and properties of some endoglucanase components with special reference to their action on cellulose when acting alone and in synergism with the cellobiohydrolase[J]. Biochemical Journal,1978,171(1): 61-72.
[12]Fujii M,Mori J I,Homma T,et al. Synergy between an endoglucanase and cellobiohydrolases from Trichoderma koningii[J]. The Chemical Engineering Journal,1995,59(3): 315-319.
[13]那安,马建华,崔福绵,等. 康氏木霉纤维素酶系中Cx酶多分子型的分离纯化及某些性质[J]. 真菌学报,1983,2(1):50-58,66.
[14]黄艳,凌敏,覃拥灵,等. 康氏木霉内切葡聚糖酶(EGⅠ)基因的克隆及表达[J]. 生物技术,2008,18(2):10-13.
[15]Samanta S,Basu A,Halder U C,et al. Characterization of trichoderma reesei endoglucanase Ⅱ expressed heterologously in pichia pastoris for better biofinishing and biostoning[J]. Journal of Microbiology,2012,50(3): 518-525.
[16]朱年青,夏文静,勇强. 里氏木霉纤维素酶的分离纯化及酶学性质[J]. 生物加工过程,2010,8(3):40-43.
[17]杨树林,孟广荣,李校堃,等. 绿色木霉MJ1产内切β-葡聚糖苷酶的分离纯化及酶学性质[J]. 微生物学通报,2006,33(5):75-79.
[18]阎伯旭,高培基. 内切葡聚糖苷水解酶的分离纯化和内源荧光性质[J]. 生物化学杂志,1997,13(5):89-94.刘波,张晓明,郭巧生,等. 百蕊草根系总RNA提取方法比较及优化[J]. 江苏农业科学,2015,43(1):44-46.
猜你喜欢
现代养生·下半月(2015年12期)2016-03-04
河北渔业(2015年11期)2015-11-18
