
卵巢切除对APP/PS1双转基因小鼠脑内老年斑及自噬的影响
2015-06-28 15:40徐明亮龙志敏廖娥钟国超杜烨鸿骆世芳贺桂琼
解放军医学杂志 2015年1期
徐明亮,龙志敏,廖娥,钟国超,杜烨鸿,骆世芳,贺桂琼
卵巢切除对APP/PS1双转基因小鼠脑内老年斑及自噬的影响
徐明亮,龙志敏,廖娥,钟国超,杜烨鸿,骆世芳,贺桂琼
目的研究雌激素对APP/PS1双转基因小鼠脑内老年斑及自噬的影响,探讨雌激素缺乏导致阿尔茨海默病(AD)的病理机制。方法将3月龄雌性APP/PS1双转基因AD模型小鼠分为卵巢切除组与假手术组,术后2个月采用免疫组化染色及透射电镜观察两组小鼠脑内老年斑及自噬功能的变化,同时检测自噬相关蛋白LC3、Beclin-1的表达情况。结果免疫组化结果显示,与假手术组相比,卵巢切除组小鼠脑内老年斑显著增加,分布更广,自噬相关蛋白LC3、Beclin-1阳性神经元数量均显著下降。电镜观察可见,与假手术组相比,卵巢切除组小鼠海马神经元及突起肿胀明显,暗神经元增加,神经元突起周围自噬小泡显著增多。结论雌激素缺乏可导致自噬泡成熟受阻、活性降低、自噬功能减弱,从而导致AD病理改变加重。
卵巢切除术;阿尔茨海默病;老年斑;自噬;小鼠,转基因
阿尔茨海默病(Alzheimer disease,AD)是一种发病机制未明的慢性进行性神经系统变性疾病[1],是临床上最为常见的痴呆类型。神经细胞外淀粉样β蛋白(amyloid β peptide,Aβ)沉积形成的老年斑是AD的典型病理特征之一[2]。研究显示,女性绝经后罹患AD的风险较同龄男性更高,这与绝经后女性脑内雌激素水平骤降及随后持续处于低水平有关[3-4],但雌激素缺乏导致AD的分子机制尚不明确。研究表明,细胞自噬异常在AD发病中起重要作用,自噬泡被认为是Aβ产生的源头[5]。那么,雌激素缺乏是否影响自噬泡的数量或活性,进而影响体内Aβ的水平及老年斑的形成?关于这一问题尚无文献报道。本课题拟以卵巢切除AD模型小鼠为研究对象,应用形态学检测手段,观察雌激素缺乏对痴呆小鼠脑内老年斑形成及自噬功能的影响,以探讨AD发生发展的病理机制。
1 材料与方法
1.1 动物及试剂 APP/PS1双转基因痴呆模型小鼠购于中国医学科学院医学实验动物研究所,体重25~35g,进行基因鉴定后喂养于重庆医科大学动物中心SPF级动物房。实验遵循实验动物使用伦理相关要求。鼠抗4G8单克隆抗体购自Convance公司,兔抗LC3多克隆抗体购自Sigma公司,兔抗Beclin-1多克隆抗体购自Abcam公司,小鼠二步法免疫组化试剂盒、兔二步法免疫组化试剂盒、DAB试剂盒均购自北京中杉金桥生物技术有限公司,其他试剂均为国产分析纯。
1.2 实验方法
1.2.1 动物模型建立 3月龄APP/PS1双转基因雌性小鼠共16只,随机分为假手术组与卵巢切除组(n=8)。经3.5%水合氯醛麻醉后暴露腹腔,切除双侧卵巢,制成雌激素缺乏模型。假手术组麻醉后暴露腹腔,不做其他处理,逐层缝合后2组小鼠均于SPF级条件中持续喂养2个月。
1.2.2 标本制备 小鼠麻醉后,开胸暴露心脏。经左心室插管至升主动脉,剪开右心耳,灌注生理盐水,冰上断头取脑。将脑组织置于4%多聚甲醛溶液中固定,常规石蜡包埋,冠状切片,厚度4μm。
1.2.3 形态学观察 免疫组化染色:石蜡切片经二甲苯脱蜡,梯度乙醇水化,PBS清洗5min×3次,枸橼酸钠微波抗原修复,室温自然冷却,PBS清洗5min×3次,3%H2O210min消除内源性过氧化物酶,PBS冲洗5min×3次,加入一抗(抗体稀释比例4G8为1:500,LC3为1:400,Beclin-1为1:400),4℃冰箱过夜。次日用PBS清洗5min×3次,加入相应二抗,室温孵育60min,PBS清洗5min×3次。加入DAB显色,显微镜下观察并适时终止显色,梯度乙醇和二甲苯1、二甲苯2脱水透明后用中性树胶封固,光镜下观察并采集图像。计数4G8阳性斑块数量并计算其面积,计数Beclin-1、LC3阳性神经元数量。透射电镜观察:小鼠用4%多聚甲醛、2.5%戊二醛灌注固定后,在解剖镜下取小鼠海马CA1区1mm3组织块,2.5%戊二醛中固定2h,1%四氧化锇固定,乙醇丙酮梯度脱水,环氧树脂包埋聚合,半薄切片定位,超薄切片,铅铀染色,透射电镜(Hitachi-7500)下观察脑组织超微结构。
1.3 统计学处理 采用SPSS 17.0软件进行数据处理。计量资料以表示,组间比较采用两样本均数的t检验。P<0.05为差异有统计学意义。
2 结 果
2.1 卵巢切除后小鼠脑皮质及海马Aβ沉积的变化
免疫组化染色显示,两组AD小鼠脑内均出现了4G8阳性斑块,说明两组小鼠脑组织神经元外均有Aβ的沉积。但假手术组小鼠脑内的散在阳性斑块主要分布在大脑皮质,海马内偶见。卵巢切除小鼠大脑皮质和海马内4G8阳性斑块数量明显增多(P=0.001)、斑块面积增加(P=0.011,图1)。透射电镜观察显示:两组小鼠海马内均出现神经元突起肿胀。与假手术组相比,卵巢切除组小鼠海马内暗神经元增多,且神经元突起周围出现数量较多的自噬小泡(图2)。
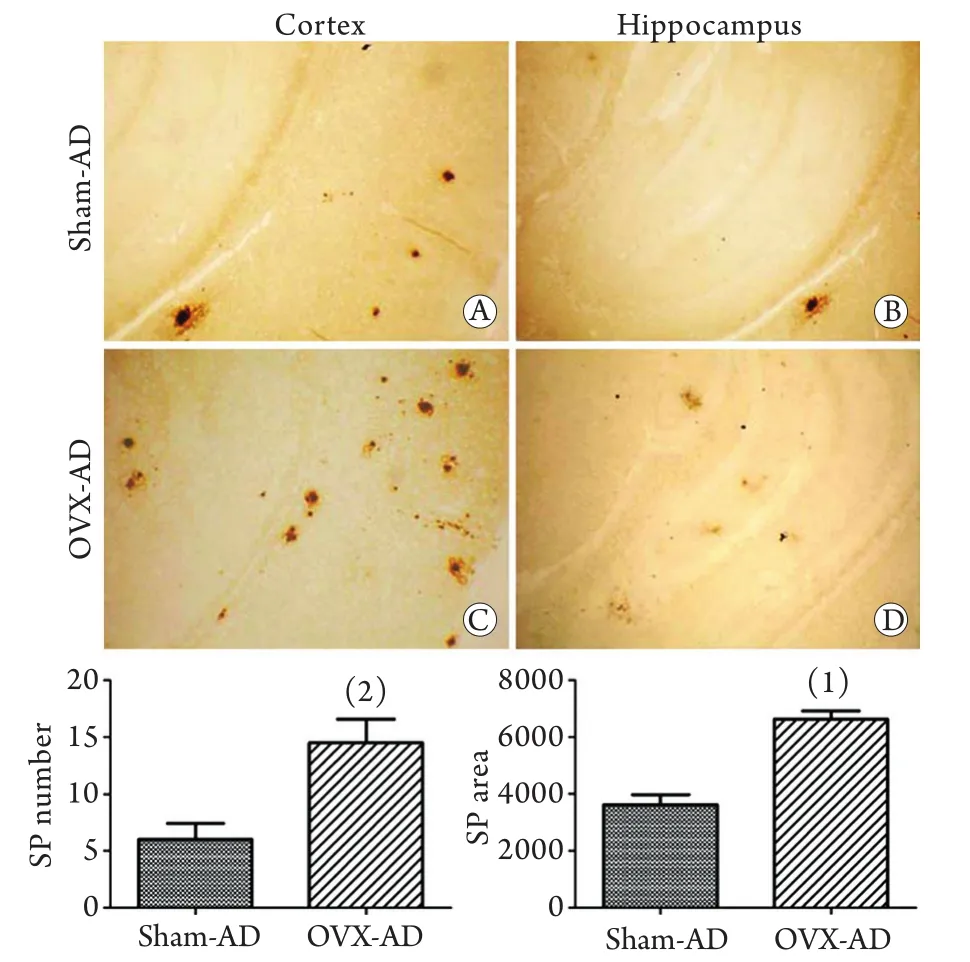
图1 两组小鼠脑内老年斑情况比较(免疫组化染色×400)Fig.1 Senile plaques (SP) in the brain of two groups of shamoperated AD (Sham-AD) and ovariectomy AD (OVX-AD) mice (Immunohistochemistry staining ×400)
2.2 卵巢切除后小鼠脑内自噬相关蛋白表达的变化 免疫组化染色显示,两组小鼠脑内均出现较多的自噬相关蛋白Beclin-1阳性神经元,假手术组小鼠脑内Beclin-1阳性神经元数量较多(88.50±12.45个),但免疫阳性物主要分布于近胞膜胞质处,卵巢切除组小鼠脑内的Beclin-1阳性神经元数目明显减少(44.00±3.74,P<0.001)且免疫阳性物均匀分布于整个胞质。另一自噬相关蛋白LC3免疫阳性细胞也广泛分布于两组小鼠脑内,假手术组小鼠脑内LC3阳性神经元数量较多(88.50±12.45个),胞体和突起浓染,且突起较长,而卵巢切除组小鼠脑内LC3阳性神经元数量明显减少(62.00±11.92个,P=0.006),虽然胞体和突起均浓染,但突起较短,提示突起可能有变性、断裂(图3)。
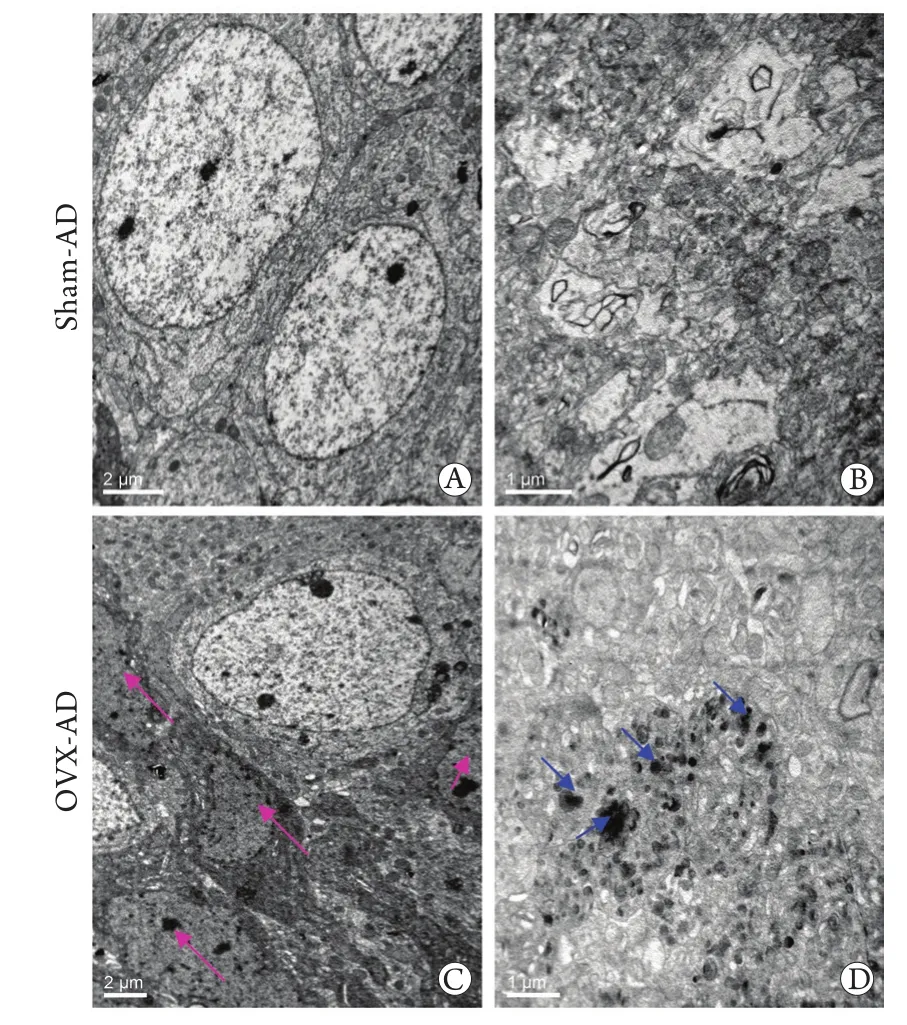
图2 两组小鼠脑组织超微结构的电镜观察(×4000)Fig.2 Ultrastructures of neurons under transmission electron microscopy (TEM) in the 2 groups (×4000)

图3 两组小鼠大脑皮质Beclin-1与LC3表达情况(免疫组化 ×400)Fig.3 Expression of Beclin-1 and LC3 in the brain of two groups of sham-operated AD and ovariectomy AD (OVX-AD) mice (Immunohistochemistry staining ×400)
3 讨 论
老年斑(senile plaque,SP)是AD脑部典型的病理特征之一[6],主要由Aβ聚集和沉积而成,Aβ被认为是AD发病的始动因素[7-9]。Aβ由淀粉蛋白前体蛋白(amyloid precursor protein,APP)经β-分泌酶和早老素1(presenilin 1,PS1)/γ-分泌酶相继切割产生[10]。研究已证实APP与PS的基因突变是引起家族性AD的主要原因[11-12]。本实验所用小鼠同时携带“瑞典家族”突变的APP(APPswe)与PS1(PS△E9)基因:APPswe基因中,突变发生在APP编码序列末端的670(Lvs/Asn)、671(Met/Leu)位点,PS△E9则是在家族性AD中发现的第9个外显子缺失突变。此类小鼠随月龄增加出现认知功能的进行性损害,脑组织出现SP沉积及神经元丢失,类似AD患者的病理特征。因此,APP/PS1双转基因小鼠被认为是研究AD病理机制及药物筛选的理想动物模型[13]。
本研究对3月龄APP/PS1小鼠行双侧卵巢切除,术后2个月观察到小鼠脑内SP数量增加,体积增大,且有大量小体积的新生SP产生,同时SP的分布范围更广,可见于大脑皮质和海马等脑区。电镜观察还发现卵巢切除小鼠脑内神经元突起肿胀明显,暗神经元数量增多,此外,Aβ沉积及神经元变性等病理特征有加重趋势,提示雌激素缺乏可加速AD的病程。本研究结果与AD流行病学调查相吻合。流行病学调查显示,绝经后妇女AD的发病率比同年龄男性高1.5~3倍,女性AD患者脑内的病理改变及认知功能障碍较男性患者严重,这可能与绝经后雌激素水平下降有关[3]。因此,本研究采用的双侧卵巢切除术AD模型小鼠可模拟临床绝经期女性AD患者,是探寻雌激素缺乏导致AD发生发展病理机制的一种可靠动物模型[14]。
本研究观察到,卵巢切除AD模型小鼠脑内自噬小泡显著增多,且主要围绕在轴突周围,提示雌激素缺乏可诱导自噬泡增加。自噬是细胞利用溶酶体降解自身受损的细胞器和错误折叠的蛋白质等有害大分子物质的重要途径[15],是真核细胞特有的生命现象[16]。细胞接受自噬诱导信号后,首先形成双层扁平状膜结构的吞噬泡,并逐渐扩展包裹待降解物质,两端融合形成密闭球状的自噬体。自噬体沿微管运行,与多泡体或内涵体融合并成熟。成熟的自噬体与溶酶体融合形成单层膜的自噬溶酶体,进而酸化,激活酸性蛋白水解酶执行降解功能[17]。自噬途径异常表现为早期自噬体在数量和体积上的增加。细胞自噬异常在AD发病中发挥重要作用[5,18-19],研究发现自噬泡(包括自噬体和自噬溶酶体)中富含APP、Aβ及γ-分泌酶,诱导自噬可引起Aβ增多,因此,自噬泡被认为是Aβ产生的场所[5,20]。AD患者及模型鼠脑中各种不成熟自噬泡的异常增多是自噬被激活的有力证据[21]。本实验观察到卵巢切除小鼠脑内Aβ沉积加重,可能系雌激素缺乏导致自噬小泡增多所致。
有研究表明,自噬具有神经保护作用,中枢神经系统基础水平的自噬受抑制可导致泛素化蛋白的累积和神经退行性疾病,诱导自噬可加速有毒物质的清除[22-23]。本研究观察到卵巢切除后自噬泡显著增加,但增加的自噬泡不仅未能清除具有神经毒性的Aβ,反而导致Aβ沉积增加,所以我们推测雌激素缺乏可能导致了自噬成熟受阻、自噬活性降低。为验证此假设,本研究观察了反映自噬活性的分子标志物Beclin-1,发现卵巢切除小鼠脑内Beclin-1表达显著低于对照组小鼠,且另一自噬相关蛋白LC3的表达改变与其一致。上述结果表明雌激素缺乏可影响自噬溶酶体的成熟,从而导致自噬活性降低、自噬功能减弱,相应地,不成熟自噬泡聚集增加,Aβ降解受阻,从而导致Aβ的沉积增多。但雌激素缺乏导致自噬异常的机制尚有待深入研究。
[1] Carrillo MC, Dean RA, Nicolas F,et al. Revisiting the framework of the National Institute on Aging-Alzheimer's Association diagnostic criteria[J]. Alzheimers Dement, 2013, 9(5): 594-601.
[2] Kobayashi K, Nakano H, Hayashi M,et al. Association of phosphorylation site of tau protein with neuronal apoptosis in Alzheimer's disease[J]. J Neurol Sci, 2003, 208(1/2): 17-24.
[3] Gillies GE, McArthur S. Estrogen actions in the brain and the basis for differential action in men and women: a case for sexspecific medicines[J]. Pharmacol Rev, 2010, 62(2): 155-198.
[4] Cahill L. Why sex matters for neuroscience[J]. Nat Rev Neurosci, 2006, 7(6): 477-484.
[5] Ohta K, Mizuno A, Ueda M,et al. Autophagy impairment stimulates PS1 expression and gamma-secretase activity[J]. Autophagy, 2010, 6(3): 345-352.
[6] Wang YR, Liu YH, Liang CR,et al.Expression profile of p75NTR and its relationship with formation of amyloid plaques in the brain of Alzheimer's disease patients[J]. Med J Chin PLA, 2014, 39(5):379-382. [王叶冉,刘雨辉,梁春荣,等.阿尔茨海默病患者脑内p75神经营养因子受体(p75NTR)的表达及与老年斑位置关系的研究[J].解放军医学杂志, 2014, 39(5): 379-382.]
[7] Reitz C. Alzheimer's disease and the amyloid cascade hypothesis: a critical review[J]. Int J Alzheimers Dis, 2012, 2012: 369808.
[8] Hardy JA, Higgins GA. Alzheimer's disease: the amyloid cascade hypothesis[J]. Science, 1992, 256(5054): 184-185.
[9] Guo XH, Lu H, Niu RN,et al. Detection of serum matrix metalloproteinase 2 and 9 in patients with Alzheimer disease[J]. J Zhengzhou Univ (Med Sci), 2014, 49(1): 137-139. [郭晓红,卢宏, 牛瑞娜, 等. 阿尔茨海默病患者血清 基质金属蛋白酶2和9水平的测定[J]. 郑州大学学报(医学版), 2014, 49(1): 137-139.]
[10] Duan LJ, Zu HB. The research development of target therapy of Alzheimer disease[J]. Chin J Pract Intern Med, 2014, 34(5): 518-520. [段立洁, 祖衡兵. 阿尔兹海默病药物治疗靶点的研究进展[J]. 中国实用内科杂志, 2014, 34(5): 518-520.]
[11] Ikeda T, Yamada M. Risk factors for Alzheimer's disease[J]. Brain Nerve, 2010, 62(7): 679-690.
[12] Michelsen KA, Korr H, Steinbusch HW,et al. Presenilin 1-related alterations in DNA integrity in a transgenic mouse model of Alzheimer's disease[J]. Brain Res, 2010, 1316: 139-144.
[13] Dickey CA, Loring JF, Montgomery J,et al. Selectively reduced expression of synaptic plasticity-related genes in amyloid precursor protein + presenilin-1 transgenic mice[J]. J Neurosci, 2003, 23(12): 5219-5226.
[14] Wang JM, Hou X, Adeosun S,et al. A dominant negative ERbeta splice variant determines the effectiveness of early or late estrogen therapy after ovariectomy in rats[J]. PLoS One, 2012, 7(3): e33493.
[15] Yang JH, Gao XG, Fu ZR.Autophagy in liver immune tolerance: mechanisms and progress[J]. Med J Chin PLA, 2014, 39(6): 503-506. [杨璟辉,高晓刚, 傅志仁. 自噬在肝脏免疫耐受中的作用机制研究进展[J]. 解放军医学杂志, 2014, 39(6): 503-506.]
[16] Kragh CL, Ubhi K, Wyss-Coray T,et al. Autophagy in dementias[J]. Brain Pathol, 2012, 22(1): 99-109.
[17] Banerjee R, Beal MF, Thomas B. Autophagy in neurodegenerative disorders: pathogenic roles and therapeutic implications[J]. Trends Neurosci, 2010, 33(12): 541-549.
[18] Wolfe DM, Lee JH, Kumar A,et al. Autophagy failure in Alzheimer's disease and the role of defective lysosomal acidification[J]. Eur J Neurosci, 2013, 37(12): 1949-1961.
[19] Salminen A, Kaarniranta K, Kauppinen A,et al. Impaired autophagy and APP processing in Alzheimer's disease: The potential role of Beclin 1 interactome[J]. Prog Neurobiol, 2013, (106/107): 33-54.
[20] Yu WH, Cuervo AM, Kumar A,et al. Macroautophagy--a novel Beta-amyloid peptide-generating pathway activated in Alzheimer's disease[J]. J Cell Biol, 2005, 171(1): 87-98.
[21] Schaeffer V, Lavenir I, Ozcelik S,et al. Stimulation of autophagy reduces neurodegeneration in a mouse model of human tauopathy[J]. Brain, 2012, 135(Pt 7): 2169-2177.
[22] Martinez-Vicente M, Cuervo AM. Autophagy and neurodegeneration: when the cleaning crew goes on strike[J]. Lancet Neurol, 2007, 6(4): 352-361.
[23] Xue Z, Zhang S, Huang L,et al. Upexpression of Beclin-1-dependent autophagy protects against beta-amyloid-induced cell injury in PC12 cells[J]. J Mol Neurosci, 2013, 51(1): 180-186.
Effects of ovariectomy on autophagy and senile plaques in the brain of adult APP/PS1 double transgenic mice
XU Ming-liang1, LONG Zhi-min1, LIAO E1, ZHONG Guo-chao1, DU Ye-hong1, LUO Shi-fang1,2, HE Gui-qiong1,2*1Institute of Neuroscience,2Department of Human Anatomy, Chongqing Medical University, Chongqing 400016, China
*Corresponding author, E-mail: guiqionghe@hotmail.com
This work was supported by the National Natural Science Foundation of China (81371221), the Program for New Century Excellent Talents in University of the National Department of Education (NCET-11-1084), the Technology Foundation for Selected Overseas Chinese Scholar, Ministry or Personnel of China[(2011)235], and the College Students' Scientific Research and Innovation Experimental Program of Chongqing Medical University (201221)
ObjectiveTo investigate the effect of ovarian estrogen on autophagy and senile plaques (SP) in the brain of adult APP/PS1 double transgenic mice, and inquire into the mechanism of Alzheimer disease (AD) neuropathogenesis due to estrogen deficiency.MethodsAPP/PS1 AD mice (3-month old) were divided into ovariectomy group (OVX-AD) and sham group (sham-AD). Two months after operation, transmission electron microscopy (TEM) was employed to observe the morphological structures of the brains of AD model mice, and immunohistochemical staining was performed to examine the changes in SP and autophagy-related proteins LC3 and Beclin-1 in the brain of AD model mice.ResultsImmunohistochemical staining showed that the number of SP in the brain of OVX-AD mice incrcased significantly compared with that in sham-AD mice, and distribution of SP was observed more extensively, including the cortex and hippocampus. The autophagy related proteins LC3 and Beclin-1 positive neurons were decreased greatly in the OVX-AD mice brain compared with that of sham-AD mice. TEM revealed a larger number of swollen and dark neurons in hippocampus of OVX-AD mice, while more autophagosome was observed around the neuronal processes in OVX-AD mice brain compared with sham-AD mice. Conclusion Estrogen deficiency may lead to delayed maturation of autophagy, decrease the activity and weaken the function of autophagy, which in turn may result in aggravation of pathological features of AD.
ovariectomy; Alzheimer disease; senile plaques; autophagy; mice, transgenic
R749.16
A
0577-7402(2015)01-0026-04
10.11855/j.issn.0577-7402.2015.01.06
2014-05-08;
2014-11-20)
(责任编辑:沈宁)
国家自然科学基金(81371221);教育部“新世纪优秀人才”支持计划(NCET-11-1084);留学人员科技活动项目择优资助[渝人社办(2011)235号];重庆医科大学大学生科研与创新实验项目(201221)
徐明亮,硕士研究生。主要从事阿尔茨海默病发病机制及治疗方面的研究
400016 重庆 重庆医科大学神经科学研究中心(徐明亮、龙志敏、廖娥、钟国超、杜烨鸿),解剖学教研室(骆世芳、贺桂琼)
]贺桂琼,E-mail:guiqionghe@hotmail.com
猜你喜欢
中老年保健(2022年2期)2022-08-24
中老年保健(2022年1期)2022-08-17
浙江医学(2020年9期)2020-07-01
保健与生活(2019年17期)2019-11-08
养生阅刊(2019年7期)2019-09-10
海峡姐妹(2019年6期)2019-06-26
浙江中西医结合杂志(2019年4期)2019-05-05
浙江医学(2019年2期)2019-01-23
奥秘(2018年9期)2018-09-25
黄河黄土黄种人(2018年5期)2018-06-07
