
八线腹链蛇食物分析及其对中华青鳉的捕食
2016-04-13 06:12陈国柱
四川动物 2016年4期
陈国柱
(国家高原湿地研究中心,西南林业大学,昆明650224)
八线腹链蛇食物分析及其对中华青鳉的捕食
陈国柱
(国家高原湿地研究中心,西南林业大学,昆明650224)
在2014—2015年4—5月、2016年4月对云南省昆明市滇池流域中华青鳉Oryziassinensis模式标本产地的八线腹链蛇Hebiusoctolineatum肠道食物组成进行了分析,共检出包括鱼类、两栖类等11个食物种类或类型,并首次发现八线腹链蛇对中华青鳉的捕食现象。在15条受检八线腹链蛇标本中3条消化道内检出中华青鳉残体,出现频率达20%。本文对此现象进行了简要报道。
八线腹链蛇;中华青鳉;捕食
八线腹链蛇Hebiusoctolineatum属游蛇科Colubridae东亚腹链蛇属Hebius(Guoetal.,2014;蔡波等,2015),又名八线游蛇,中国特有种(周正彦等,2008),主要分布在云南、四川、贵州(杨大同,饶定齐,2008)及广西百色(曾小飚,2009)等海拔较高地区,垂直分布海拔700~2 460 m(赵尔宓,2006),成熟雄性个体体长170~824 mm、雌性203~845 mm(赵尔宓,2006)。在云南滇池流域,八线腹链蛇是优势蛇种(何晓瑞,1983a,1983b;何晓瑞等,2002),主要栖息在湖滨、池塘、沼泽及溪流等有水及湿润、半湿润生境,对鱼类、两栖类有较强的捕食作用(何晓瑞,1983a,1983b)。在贵州省,它甚至捕食国家Ⅱ级重点保护濒危物种——贵州疣螈Tylototritonkweichowensis(伍律等,1985)。数十年来,随着滇池流域湿地环境的剧烈改变、入侵物种的大量进入,滇池流域鱼类、两栖类群落发生了显著的改变,八线腹链蛇仍为常见蛇种(何晓瑞等,2002),尤其在滇池周边人类活动干扰较少的山区湿地生境较为活跃。
中华青鳉Oryziassinensis是一种小型鱼类,隶属于大颌鳉科Adrianichthyida青鳉属Oryzias,曾在云南地区广泛分布,近半个多世纪以来在栖息地被破坏、入侵物种特别是食蚊鱼Gambusiaaffinis入侵的影响下,其分布区域、种群数量在近年已显著下降,出现局部灭绝(陈银瑞等,1989)。在滇池流域,湖区的中华青鳉种群已经消失(陈自明等,2001),孑遗种群目前主要分布在滇池北部若干区域。早期世界自然保护联盟(IUCN)红色名录曾将中华青鳉列为易危级别(vulnerable,VU),近年的评估却因为缺乏足够数据,暂时将其列入较少关注级别(least concern,LC)(IUCN,2012)。在国内,中华青鳉已经被列入《中国动物红皮书 鱼类》红色名录,对其基础生态及保护生物学研究亟待开展。距离滇池仅5 km的云南省昆明市西山区团结乡花红洞山间盆地是中华青鳉模式标本产地(陈银瑞等,1989;杜丽娜等,2013),自1989年定名以后,该地种群延续至今将近30年。2012—2016年笔者对该地中华青鳉的种群生态进行了长期研究,初步结果显示,在食蚊鱼、克氏原螯虾Procambarusclarkii及其他入侵物种的影响下,该地种群数量剧烈下降。在这种情况下,任何对其种群数量产生不利影响的因素都可能加速其种群的衰亡速度。2014年4月在该地首次观察到土著捕食者——八线腹链蛇守候在因水分蒸发而形成的水洼处捕食鱼类的现象,通过解剖在其胃内发现了中华青鳉残体,这提示天敌捕食对中华青鳉种群数量的下降可能有重要影响。
为初步揭示八线腹链蛇捕食作用对中华青鳉种群的可能性影响,本文对八线腹链蛇在该区域的食物进行了分析,以了解现今滇池流域现存土著物种间可能存在的捕食关系,为进一步深入研究滇池流域土著物种-入侵物种间复杂的关系积累基础资料。
1 材料与方法
1.1 样品采集地
样品采集地位于云南省昆明市西郊团结乡花红园社区所在的山间盆地,距滇池草海的直线距离为5 km。盆地中心海拔约2 167 m,四周被海拔2 300~2 500 m山峰所群绕。年均气温14.5 ℃,年均降水量约1 031 mm,干湿季明显,一般情况下6—10月为雨季,11月至翌年5月为旱季,个别年份雨季会提前或延后。盆地西南侧为三家村水库,水库坝下为低缓的丘陵盆地,若干灌溉渠道纵横交错,出水口为地下溶洞,可能以地下河形式汇入滇池草海流域。拥有大村、小村、三家村等3座村庄。汇水面积约6 km2。近年该区低洼耕地在丢荒后出现众多的山地沼泽,形成水库、沼泽、沟渠等封闭的高原湿地生态系统,地形及湿地生境详细描述参见仇玉萍等(2015)。
1.2 样品采集与分析
根据每年雨季开始的时间及湿地水量情况不同,在2014—2015年4—5月、2016年4月分别进行了样品采集。采集时沿湿地区域巡视,当发现八线腹链蛇时即用网孔2 mm、网口直径30 cm的手抄网捕捉。样品用10%福尔马林溶液现场固定,然后带回实验室解剖检查。在实验室测量八线腹链蛇体长、体质量等指标。在体视显微镜下检视其消化道内容物。未消化个体鉴定其种类。对已消化分解残渣根据难分解物质如眼珠、耳石、平衡石等鉴定其种类。无法辨别的食物残渣记述其丰富程度。
2 结果
研究期间共采得八线腹链蛇15条,平均体长37.1 cm±16.5 cm,平均体质量24.8 g±24.7 g,主要为幼年个体(表1)。对所有样品分析显示,花红园区域八线腹链蛇的食物包括中华青鳉、小黄黝鱼Micropercopsswinhonis、蛙类(昭觉林蛙Ranachaochiaoensis)、蝌蚪、克氏原螯虾、摇蚊幼虫、其他昆虫及不能分辨种类的动物性残渣、脊椎骨等(表1)。
在3条八线腹链蛇消化道中发现中华青鳉残体,比例达20%。1号标本在被捕前正守候在浅水洼地水草内伺机捕捉因水分蒸发而汇集的鱼类(~200尾/m2)(图1:A),在其消化道内发现中华青鳉、小黄黝鱼各1尾,且未被消化(图1:B,C,D),2尾鱼体全长分别为30.87 mm和36.98 mm,体质量分别为265.4 mg和560.1 mg。在6号标本中发现桡足类1只,草梗1小段。7号标本消化道中的脊椎较小,约0.75 mm × 0.7 mm,且观察到眼珠1颗,直径0.5 mm,估计其残渣应为鱼类而不是两栖类。
此外,在水库漫滩区域观察到1条八线腹链蛇正在捕食华西雨蛙Hylaannectans(图2),该蛇体长约26.2 cm,体质量约4.5 g,而其捕食的华西雨蛙体质量则达2.7 g。
3 讨论
八线腹链蛇是高原蛇类(赵尔宓,2006),我国特有种,目前对其食物组成及摄食生态学特征的了解尚少。综合何晓瑞(1983b)、伍律等(1984,1985)及本文的观察资料,其可辨认、见于科学文献的食物种类近17种(表2),鱼类、两栖类是其主要捕食类型,这与游蛇科多数喜水栖种类的食物组成近似。对滇池流域的八线腹链蛇种群而言,何晓瑞(1983b)曾记载其在滇池流域大量捕食泥鳅Misgurnusanguillicaudatus、多鳞白鱼Anabariliuspolylepis、麦穗鱼Pseudorasboraparva、鲫鱼Carassiusauratus等,鱼类可占其食物总重量的62.9%;且常在养殖池内捕食鱼苗。本文还发现八线腹链蛇对中华青鳉等的捕食,进一步增加了对其食物组成种类的认识。特别是八线腹链蛇对入侵物种——小黄黝鱼、克氏原螯虾的捕食,提示八线腹链蛇对滇池水生生物群落的演变具有较强的适应性。
另一方面,游蛇科许多种类对鱼类有较强捕食作用(徐火弟,1974;Mushinskyetal.,1982)。如,西藏温泉蛇Thermophisbaileyi大量捕食异齿裂腹鱼Schizathoraxo’connori、高原鳅Triplophysasp.等(李键,2007);虎斑颈槽蛇Rhabdophistigrina大量捕食鲫鱼等(贾少波等,2000);棋斑游蛇Natrixtessellata栖息在水边时主要以鱼类为食(王国英等,1987);红纹滞卵蛇(旧称红点锦蛇)Oocatochusrufodorsatus嗜食泥鳅(黄美华等,1974)。八线腹链蛇也是一种嗜食鱼类的游蛇科蛇,它对鱼类的捕食是否能对滇池流域鱼类种群动态产生影响尚无系统深入的研究。

图1 A. 正守候捕食的八线腹链蛇Hebiusoctolineatum(红色箭头); B. 解剖前形态;
C. 解剖后形态, 白色箭头示意中华青鳉Oryziassinensis; D. 上: 中华青鳉O.sinensis, 下: 小黄黝鱼Micropercopsswinhonis
Fig. 1 A.H.octolineatum(red arrow) is waiting to attack the fish; B. Before anatomy;
C. After anatomy,O.sinensiswas detected (white arrow); D. Upper:O.sinensis, lower:M.swinhonis

图2 八线腹链蛇捕食华西雨蛙Fig. 2 Hebius octolineatum is preying on Hyla annectans表1 八线腹链蛇食物分析Table 1 Food analysis of Hebius octolineatum

年份Year序号ID体长Totallength/cm体质量Weight/g采集生境Habitats食物分析FoodcompositionsF1F2F3F4F5F6F7F8F9F10F112014122.43.8沼泽洼地11000000000217.91.7沟渠0000000000++348.826.0水库00010000000418.22.2沟渠1000043000++++522.04.5水库1000050010+623.13.3沼泽洼地0000000030++756.138.3水库000000800+++++2015856.070.8水库0002000120++936.212.2水库0000000000+1057.055.0水库0001000000+20161138.211.4沼泽洼地00+0000000++1231.550.0沟渠0000500000++1363.063.1沟渠0000300000++1419.02.1沟渠0000000000++1549.528.3沟渠00001000000平均值±标准差Mean±SD37.1±16.524.8±24.7频率Frequency/%20.06.76.720.020.013.313.36.720.06.780.0
注Notes: F1. 中华青鳉Oryziassinensis, F2. 小黄黝鱼Micropercopsswinhonis, F3. 鱼类残骸Fish debris, F4. 蛙类Frog, F5. 蝌蚪Tadpole, F6. 克氏原螯虾Procambarusclarkii, F7. 摇蚊幼虫Chironomidaesp. larval, F8. 蚱蜢类Grasshopper, F9. 其他昆虫Other insects, F10. 脊椎骨Vertebra, F11. 动物残渣Animal residue; +, ……, ++++. 不同符号表示动物性脊椎骨、食物残渣定性相对丰度, represented the qualitative abundance of vertebra and animal residue in the guts of samples, +. 稀有Rare, ++. 较多More, +++. 多Plenty, ++++. 很多Plenty more.
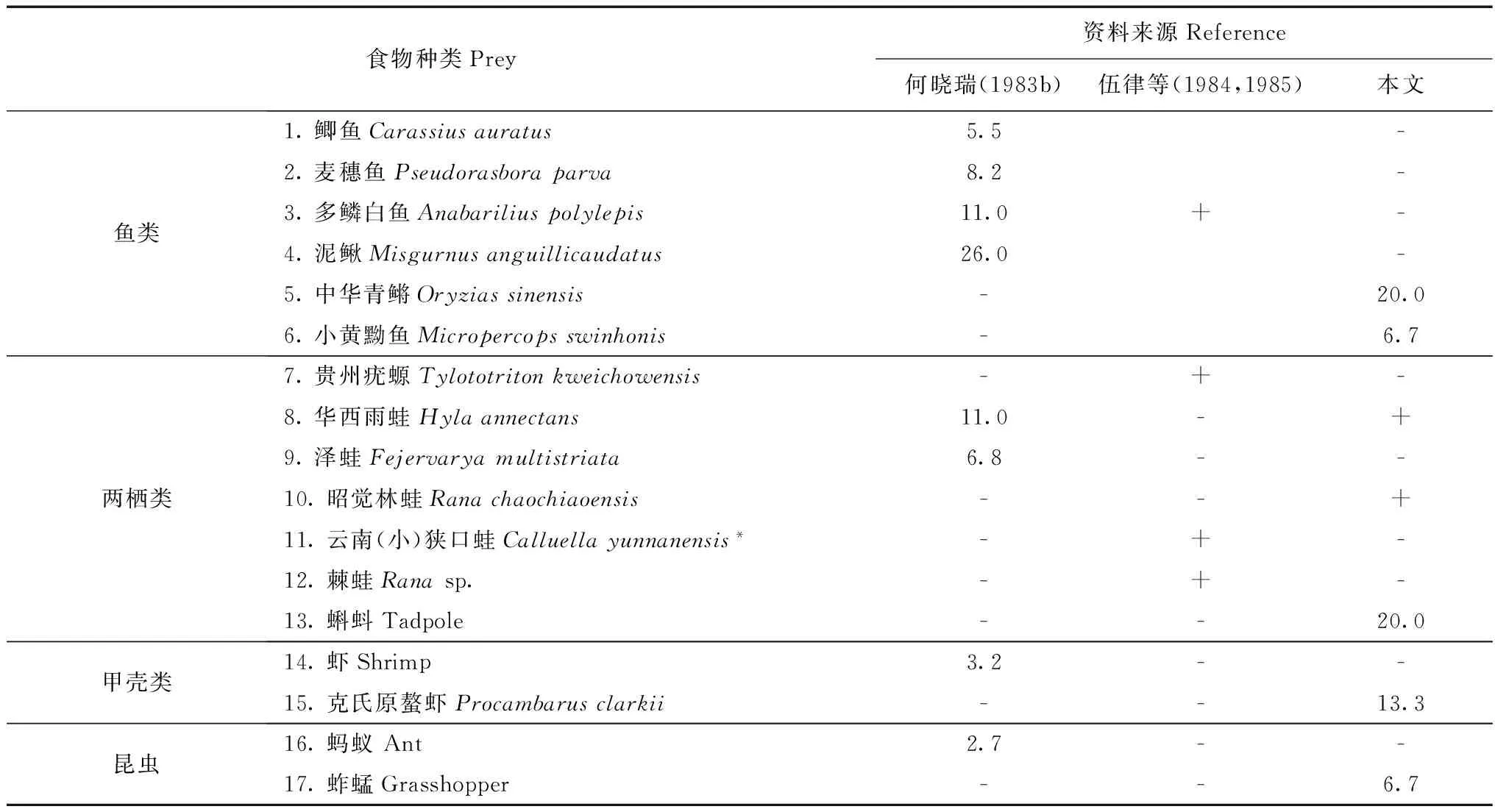
表2 八线腹链蛇食物组成Table 2 Food composition of Hebius octolineatum
注:数字为食物出现频率; -. 资料中无该食物记载, +. 资料中对该食物进行了记载, 但无具体的比例数据;*原文叙述为云南狭口蛙,后无拉丁学名, 查勘疑为云南小狭口蛙之误, 此处予以更正。
Notes: the numbers are the frequency of relative food items; -. no records of certain food items in the references; +. certain food items are reported in the references but no available frequency data;*the species noted in Chinese name without a scientific name in the reference, and we corrected it asCalluellayunnanensis.
许多研究认为,蛇类对鱼类的捕食作用受鱼类密度影响,例如,束带蛇Thamnophissirtalis在鱼类孵化场大量捕食鱼类,而在野外其他环境中对鱼类的捕食比例则较少(Gregory & Nelson,1991),菱斑水蛇Nerodiarhombifera也极喜欢进入鱼类密度极高的鱼类孵化场捕食鱼类(Plummer & Goy,1984)。每年4—5月是滇池流域的旱季末期,此时花红园地区沼泽、池塘等湿地生境由于水分的强烈蒸发而形成分离的浅水洼,水库区域则因退水形成大片低洼草甸、水洼交替生境,中华青鳉被迫聚集在狭小空间内,种群密度较高,容易吸引捕食者的注意,也增加了八线腹链蛇对其捕食的机会。另一方面,中华青鳉在该地直至雨季开始的6—8月才大量繁殖,在繁殖季节前的个体保有量将直接影响其当年的种群增长潜力(繁殖补充过低易导致种群数量的剧烈下降),八线腹链蛇在旱季结束前进入中华青鳉聚集的洼地生境并对其大量捕食,对中华青鳉种群数量影响不容忽视。特别是该地由于食蚊鱼的入侵,中华青鳉种群数量已显著下降的形势下,这种捕食压力更应当谨慎评估。如中国水蛇Enhydrischinensis对濒危物种唐鱼Tanichthysalbonubes的捕食被认为是唐鱼致危因素之一(易祖盛等,2004),这提示八线腹链蛇的捕食作用可能对中华青鳉在滇池流域的孑遗种群续存有负面压力,有待进一步深入研究。
蔡波, 王跃招, 陈跃英, 等. 2015. 中国爬行纲动物分类厘定[J]. 生物多样性, 23(3): 365-382.
陈银瑞, 宇和纮, 褚新洛. 1989. 云南青鳉鱼类的分类和分布[J]. 动物分类学报, 14(2): 239-246.
陈自明, 杨君兴, 苏瑞凤, 等. 2001. 滇池土著鱼类现状[J]. 生物多样性, 9(4): 407-413.
杜丽娜, 陈小勇, 杨君兴. 2013. 中国科学院昆明动物研究所鱼类模式标本名录[J]. 动物学研究, 34(4): 345-360.
何晓瑞, 王紫江, 吴金亮. 2002. 昆明地区爬行动物的研究[J]. 四川动物, 21(1): 8-11.
何晓瑞. 1983a. 八线游蛇生物学的初步研究[J]. 两栖爬行动物学报, 2(2): 11-17.
何晓瑞. 1983b. 昆明地区的蛇类[J]. 云南大学学报, 11: 40-47.
黄美华, 周世安, 谢占泰, 等. 1974. 红点锦蛇生态的观察[J]. 动物学杂志, 3: 23.
贾少波, 张树军, 马晓明. 2000. 山东聊城虎斑颈槽蛇生态学特性初步研究[J]. 两栖爬行动物学研究(第8辑): 161-164.
李键. 2007. 中国特有物种西藏温泉蛇的食性及取食行为[J]. 西藏大学学报, 22(2): 84-90.
仇玉萍, 刘勇, 孙大成, 等. 2015. 利用ISSR技术分析滇池流域中华青鳉(Oryziassinensis)遗传多样性(英文)[J]. 湿地科学, 13(5): 543-550.
王国英, 齐卫东, 马鸣, 等. 1987. 棋斑游蛇种群生态观察[J]. 干旱区研究, 2: 35-40, 28.
伍律, 李德俊, 刘积深. 1985. 贵州爬行类志[M]. 贵阳: 贵州人民出版社: 183-187, 435.
伍律, 刘积琛, 李德俊. 1984. 贵州省蛇类食性的初步分析[J]. 动物学杂志, 6: 22-25.
徐火弟. 1974. 清除吃鱼的水蛇[J]. 水产科技情报, 6: 16.
杨大同, 饶定齐. 2008. 云南两栖爬行动物[J]. 昆明: 云南科技出版社: 260-262.
易祖盛, 陈湘粦, 巫锦雄, 等. 2004. 野生唐鱼在广东的再发现[J]. 动物学研究, 25(6): 551-555.
曾小飚. 2009. 广西百色蛇类资源调查初报[J]. 四川动物, 28(5): 753-755.
赵尔宓. 2006. 中国蛇类[M]. 合肥: 安徽科学技术出版社: 167-168.
周正彦, 李丕鹏, 陆宇燕, 等. 2008. 中国特有蛇类资源及保护建议[J]. 四川动物, 27(1): 44-47.
Gregory PT, Nelson KJ. 1991. Predation on fish andintersite variation in the diet of common garter snakes,Thamnophissirtalis, on Vancouver Island[J]. Canadian Journal of Zoology, 69(4): 988-994.
Guo P, Zhu F, Liu Q,etal. 2014. A taxonomic revision of the Asian keelback snakes, genusAmphiesma(Serpentes: Colubridae: Natricinae), with description of a new species[J]. Zootaxa, 3873(4): 425-440.
IUCN. 2012. IUCN red list categories and criteria: version 3.1. 2nd ed[M]. Gland, Switzerland and Cambridge, UK: IUCN.
Mushinsky HR, Hebrard JJ, Vodopich DS. 1982. Ontogeny of water snake foraging ecology[J]. Ecology, 63(6): 1624-1629.
Plummer MV, Goy JM. 1984. Ontogenetic dietary shift of water snakes (Nerodiarhombifera) in a fish hatchery[J]. Copeia, 2: 550-552.
Food Analysis ofHebiusoctolineatumand its Predation onOryziassinensis
CHEN Guozhu
(National Plateau Wetland Research Center, Southwest Forestry University, Kunming 650224, China)
Food composition of the snake,Hebiusoctolineatum, was analyzed during April or May (before rainy season) from 2014 to 2016 in Dianchi Lake, Kunming city, Yunnan province. The results showed that 11 food types were detected. Especially, the endangered fish species, Chinese medaka (Oryziassinensis), was found for the first time in the guts of threeH.octolineatum(n=15). As the population ofO.sinensishad significantly declined since the invasive mosquitofish (Gambusiaaffinis) was introduced, the effects ofH.octolineatumpredation on the dynamics ofO.sinensispopulation need to be estimated.
Hebiusoctolineatum;Oryziassinensis; predation
2016-04-21 接受日期:2016-06-14 基金项目:国家自然科学基金青年项目(31400477)
陈国柱(1979—), 男, 博士, 讲师, 主要从事鱼类入侵生态学及濒危物种保护生物学研究, E-mail:chenguozhu79@163.com
10.11984/j.issn.1000-7083.20160093
Q959.6; Q959.4
A
1000-7083(2016)04-0588-05
猜你喜欢
今日农业(2022年15期)2022-09-20
海洋信息技术与应用(2022年1期)2022-06-05
文苑(2020年8期)2020-09-09
儿童时代·幸福宝宝(2020年9期)2020-09-08
红土地(2018年7期)2018-09-26
时代风采(2016年10期)2016-07-21
当代畜禽养殖业(2014年10期)2014-02-27
西南学林(2013年1期)2013-11-22
西南学林(2013年2期)2013-11-12
中学生物学(2008年6期)2008-08-29
