
切花非洲菊采后弯茎机制初探
2016-05-30 02:32祝小云王晰徐哲赖齐贤陈雯
热带作物学报 2016年5期
关键词:乙烯
祝小云 王晰 徐哲 赖齐贤 陈雯


摘 要 非洲菊(Gerbera jamesonii Bolus)是重要的切花材料,然而,切花非洲菊采后易发生弯茎现象,是导致采后损耗的主要原因。本研究以10个非洲菊切花品种为试材比较瓶插期间弯茎进程,发现弯茎现象在品种之间存在显著差异,并从中筛选出易弯茎品种和不弯茎品种。对非洲菊各品种进行外源乙烯和乙烯作用抑制剂1-MCP处理,发现非洲菊切花对于乙烯敏感性也存在品种特异性,并且乙烯对敏感型品种的弯茎发生有延迟或促进的不同影响效果。同时,对瓶插期间各品种花茎不同区域干、鲜重情况分析,发现非洲菊花茎距离花盘底部10~15 cm区域的干重、鲜重和水分损失均较高,可能是导致该区域发生弯茎的主要原因之一。此外,本文还发现非洲菊花茎直径和茎段的再伸长与弯茎现象的发生没有联系。
关键词 非洲菊;弯茎;乙烯;水分损失
中图分类号 S682.1 文献标识码 A
Abstract Gerbera jamesonii Bolus is one of the most important cut-flowers. However, stem bending often occurs during the postharvest period of Gerbera flowers, and is a major reason for postharvest loss of Gerbera flowers. In this study, ten cultivars of Gerbera flower were chosen as the materials. By measuring the occurrence time of stem bending during the vase life, we found that the phenotypes of stem bending were quite different among cultivars, and we screened stem-bending liable cultivars and stem-bending resistant ones. Treatments of exogenous ethylene and 1-MCP, which is an ethylene action inhibitor, indicated that the responses of Gerbera flowers to ethylene were cultivar-dependent. And to the ethylene-sensitive cultivars, ethylene treatment had either positive or negative effect on the occurrence time of stem bending. Measurement of fresh weight and dry weight of segments along stems in tested Gerbera cultivars showed that the loss of fresh weight, dry weight and water of 10-15 cm region beneath the flower disk was higher than that of other stem regions, and it might lead to stem bending on this region. In addition, no relationship was found between stem diameter and stem elongation with the occurrence time of stem bending.
Key words Gerbera;Stem bending;Ethylene;Water loss
doi 10.3969/j.issn.1000-2561.2016.05.012
非洲菊(Gerbera jamesonii Bolus)是世界上最受欢迎的鲜切花之一。非洲菊切花品质包括花型、花盘大小、花茎质量和瓶插寿命等。其中,瓶插寿命是衡量切花商品价值的重要因素之一。通常,切花非洲菊瓶插寿命决定于两个方面,即花瓣萎蔫状况及花茎弯曲程度。非洲菊切花在瓶插阶段发生的花茎不规则弯折现象,称为弯茎现象[1-2]。非洲菊切花的弯茎现象在采后发生普遍,并且往往先于花瓣的萎蔫而发生,是切花瓶插寿命的限制性因素,也是导致采后损耗的重要原因。
非洲菊切花的弯茎常发生在花茎的特定区域,集中在距花冠基部8~15 cm的区段;对于不同品种,弯茎发生区域略有不同[2-3]。同时,切花非洲菊的弯茎具有品种特异性,不同品种在弯茎发生时间和弯茎率方面往往存在较大差异。Ferrante等[4]对20个非洲菊切花品种在瓶插期的弯茎情况进行了比较,发现‘Dame blanche,‘Olina,‘Dino,‘Sensation和‘Venice5个品种弯颈率高达100%,而‘Zembla, ‘Dalma, ‘Mammut和‘Lindessa4个品种的弯颈率为0,即不发生弯茎,后者的瓶插寿命仅由舌状花瓣的萎蔫程度决定。在瓶插寿命方面,‘Miria发生弯茎的时间最短,约为5 d,而‘Lindessa最长,约为23 d。吕长平等[5]对国内13个非洲菊切花品种进行了瓶插寿命比较,也得到品种间弯茎现象差异显著的相似结论。然而,造成品种间弯茎现象差异的原因仍不清楚。
近年来的研究表明,非洲菊切花弯茎的发生是一个复杂的生理过程,受到环境因素和植物内在因素的综合调控,包括微生物侵染及花茎死亡细胞的分泌物导致的导管堵塞[6-8],花茎组织结构和机械支持力[9-12],弯茎发生区域水分平衡、细胞壁物质代谢和活性氧代谢[2,13-14],及植物激素乙烯[15]和赤霉素[16]的调控等。迄今为止,关于非洲菊弯茎发生机制的解析依然不够深入,尤其是乙烯对于非洲菊切花衰老的影响,还没有统一的定论。一些学者认为非洲菊对乙烯不敏感,为非跃变型切花,外源乙烯处理对非洲菊切花的衰老影响不大[14,17-18]。有的学者则认为非洲菊切花对乙烯的敏感性与品种有关,不同的品种对乙烯的敏感性存在显著差异[19]。
为了进一步研究非洲菊切花采后弯茎发生的机制,本研究通过弯茎现象观察及生理指标测定,对市场上常见非洲菊品种中易弯茎和不易弯茎品种进行筛选;同时,探究弯茎发生与乙烯、干鲜重变化、水分平衡、茎伸长和花茎直径等因素之间的联系,初步明确非洲菊切花弯茎的主要原因,为开发新型非洲菊切花保鲜技术提供了理论依据。
1 材料与方法
1.1 试验材料
试验花材取自浙江海盐花卉基地。试验所选取的10个非洲菊(Gerbera jamesonii Bolus)品种依次为: ‘北极星、 ‘玫红、 ‘白马王子、 ‘玲珑粉、‘老橘黄、 ‘金太阳、 ‘黄绿心、 ‘鸿运、 ‘水粉、 ‘向日葵(图1)。挑选处于Ⅵ-Ⅶ期(外层舌状花完全开放、内层管状花开放1~2 轮),花茎长度为50 cm左右,花茎健壮,无病虫害的花材进行采收[20],采摘时用手抓住花茎,顺着花茎的生长方向往外拉动,使花茎从根颈处断离。采收后应立即将花茎基部浸入水中,并快速运至实验室。以每支切花花茎长度为 40 cm为标准,用刀片在水中进行45°斜切。切花瓶插于含 250 mL蒸馏水的塑料瓶中,每瓶中放置3支切花。瓶插室温度22 ℃,湿度60%。每隔1天将花茎在水中斜剪去约 1 cm长度,每天更换蒸馏水。
1.2 试验方法
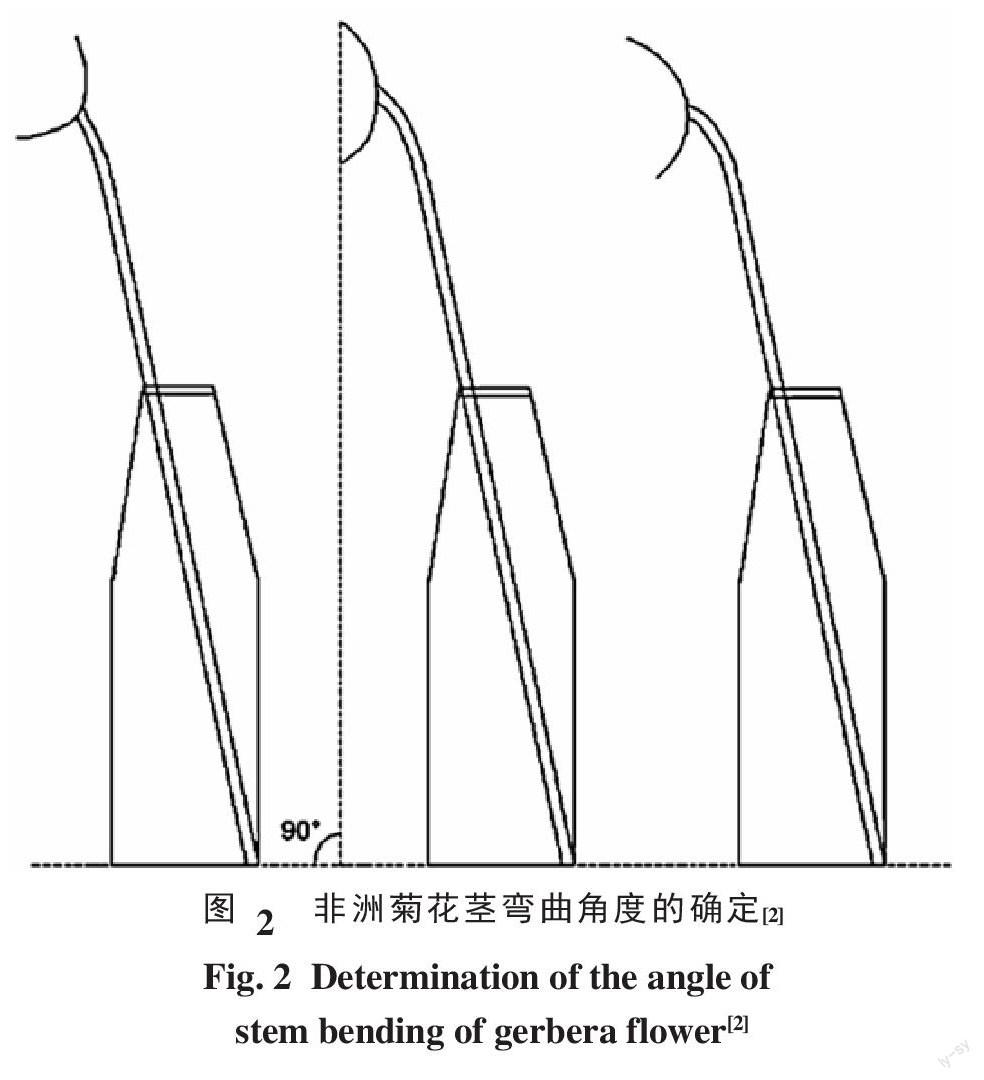
1.2.1 花茎弯茎级数确定 非洲菊瓶插试验中,花茎放置于瓶中,花茎相对于竖直面约为20°。测量时确保花茎底部固定于瓶子下端,测出花盘平面与水平面的角度即为弯颈角度。当该角度为0°~90°范围内判定为未发生弯颈(图2左);当角度为90°时判定为弯茎发生的零界点(图2中);角度超过90°的进程则判定为弯茎发生进程,此时花茎弯曲程度不断加剧(图2右)。非洲菊切花弯茎级别参考Perik等[2]并作以下修改:1级,花盘平面与水平方向夹角在0°~30°:2级,花盘平面与水平方向夹角在30°~60°:3级,花盘平面与水平方向夹角在60°~90°:4级,花盘平面与水平方向夹角在90°~120°:5级,花盘平面与水平方向夹角在120°~150°;6级,花盘平面与水平方向夹角在150°~180°。弯茎发生时间:弯茎程度已达4级及4级以上且不再恢复,当日计为弯茎发生日。弯茎发生部位:花茎弯曲的中心点到花盘底部的距离。非洲菊切花弯茎程度每天测量。每一品种15枝。
1.2.2 乙烯和1-MCP处理 试验共设有乙烯处理组、1-MCP处理组和对照组,每组10个品种,每个品种20枝。将各组花材放置于盛有蒸馏水的容器中,密封于纸箱内。乙烯气体通过注射器注入纸箱内,处理浓度为100 μg/mL[4];1-MCP粉末置于培养皿中放置于纸箱内,待纸箱密封后将蒸馏水通过注射器注入纸箱内的培养皿中,使1-MCP溶于水并蒸发,处理浓度为2 μg/mL。注射后迅速用胶带将注射口密封。对照组为同样密闭的纸箱。同时,每个封箱内装有 25 mL的 1 mol/L NaOH溶液吸收CO2气体。将纸箱置于温度为22°C环境中处理 24 h。处理后将各组切花置于瓶插室中,温度为22 ℃,湿度60%。每日观察并确定弯茎级别和发生时间。
1.2.3 花茎不同区域干、鲜重损失和失水率测定 在瓶插期间第0天和第8天随机选取各品种花盘大小和花茎直径一致的10支切花,以 5 cm 为单位截取茎杆0~5 cm、5~10 cm、10~15 cm、15~20 cm、20~25 cm、25~30 cm茎段,测量鲜重;随后放置于65 ℃烘箱中,96 h后测量干重。计算各品种茎段在瓶插过程中干、鲜重损失率和失水率。
鲜重损失率=(0 d鲜重-8 d鲜重)/0 d鲜重×100%
干重损失率=(0 d干重-8 d干重)/0 d干重×100%
失水率=[(0 d鲜重-0 d干重)-(8 d鲜重-8 d干重)]/(0 d鲜重-0 d干重)×100%
1.2.4 花茎伸长量测定 瓶插进行之前,在所有供试非洲菊品种花茎上从花盘底部开始标记0 、5 、10 、15 cm、20 cm长度标记点。瓶插前4 d每天测量标记之间的长度,计算每个茎段的伸长量。
1.3 数据处理
采用SPSS 16.0进行数据统计和分析; 利用Excel软件进行作图。
2 结果与分析
2.1 非洲菊切花易弯茎和不易弯茎品种筛选
2.1.1 非洲菊不同品种弯茎发生时间分析 对非洲菊10个切花品种统计了弯茎发生的具体时间,如图3、表1所示,非洲菊不同切花品种的弯茎发生时间存在显著差异。在发生弯茎的品种中,弯茎最早的是‘黄绿心,约2.5 d, 最晚的是‘玲珑粉,约8.2 d。弯茎发生时间居于两者之间的品种为‘玫红(4.2 d), ‘金太阳(4.6 d), ‘老橘黄(5.3 d), ‘向日葵(6.6 d), ‘北极星(7.9 d), ‘玲珑粉(8.2 d)。‘白马王子和‘鸿运在瓶插阶段始终不发生弯茎。按照各品种非洲菊弯茎发生时间将其划分为: 1)易弯茎品种(< 4 d): ‘黄绿心、 ‘水粉; 2)较易弯茎品种(4 d~9 d): ‘玫红、 ‘金太阳、 ‘老橘黄、 ‘向日葵、 ‘极星'、 ‘玲珑粉; 3)不弯茎品种: ‘白马王子、 ‘鸿运。
在非洲菊切花弯茎过程中,弯茎发生部位与花茎折断部位不相同。由表1所示,非洲菊弯茎发生部位多位于距花冠基部6~9 cm茎段处。其中,弯茎发生部位距离花盘最近的是‘北极星,约6.9 cm处;最远的是‘老橘黄,约8.8 cm处,两者相差近2 cm。另外,非洲菊各品种间花茎折断位置差异较小,多位于距花冠基部11~13 cm茎段处。其中,‘老橘黄折断位置距花盘最远,约13.0,而‘金太阳最近,约11.0。综上所述,在非洲菊切花弯茎发生至花茎折断过程中,弯茎区域并不是固定不变的,弯茎中心逐渐远离花冠基部。因此,距花冠基部约7~13 cm的茎段是发生弯茎进程的主要区域,可能与该茎段的生理特性相关。
2.1.2 非洲菊花茎直径对于弯茎发生的影响 由表1所示,各品种非洲菊弯茎部位花茎直径范围约0.5~0.6 cm,直径由大到小依次为‘玫红、‘向日葵、 ‘黄绿心、 ‘玲珑粉、 ‘鸿运、 ‘老橘黄、‘水粉、 ‘白马王子、 ‘北极星、 ‘金太阳。其中花茎直径最大的是‘玫红0.62 cm,最小的是‘金太阳0.48 cm,前者约为后者的1.3倍。通过比较非洲菊花茎直径与弯茎发生时间,并未发现花茎越细,则越容易发生弯茎的趋势。如不弯茎品种‘白马王子‘花茎直径仅为0.55 cm,而易弯茎品种‘黄绿心花茎为0.59 cm,说明非洲菊品种间花茎直径大小与弯茎发生没有必然联系。
2.2 乙烯和乙烯抑制剂1-MCP处理对于非洲菊切花弯茎发生的影响
如表2所示,大部分品种,包括‘金太阳、‘老橘黄、‘向日葵、‘北极星、‘白马王子和‘鸿运的弯茎发生时间不受乙烯和1-MCP处理的影响,即这6个品种为表现为乙烯非敏感性。然而, ‘黄绿心、 ‘水粉、‘ 玫红和‘玲珑粉4个品种的弯茎发生时间在乙烯和/或1-MCP处理后发生了不同程度的变化。其中,‘黄绿心的弯茎发生时间在乙烯处理后推迟至对照的1.5倍,而1-MCP处理对弯茎发生没有显著影响。‘水粉的弯茎发生时间在乙烯处理后推迟到对照的1.3倍,在1-MCP处理后相比对照显著提前约52.6%,为典型的乙烯敏感型品种。‘玫红的弯茎发生时间不受乙烯处理影响,但在1-MCP处理后延长约23.3%。‘玲珑粉的弯茎发生时间则在乙烯处理后提前约0.5 d,达到显著差异水平,而1-MCP处理对弯茎发生没有显著影响。这些结果说明非洲菊切花对乙烯的反应较为复杂,大部分品种对乙烯不敏感,仅少数品种存在不同程度的乙烯敏感性,这个现象可能与非洲菊本身的遗传背景相关。同时,乙烯和1-MCP对于非洲菊切花乙烯敏感型品种的弯茎发生具有促进或延缓的不同效果。
2.3 非洲菊切花瓶插进程中茎干损耗对于非洲菊切花弯茎发生的影响
由图4可知瓶插0 d时,花茎干、鲜重总体趋势为靠近花盘基部的茎段,尤其是0~5 cm茎段的数值较低;干重在远离花盘基部的区域数值较高;大部分品种的鲜重在5~20 cm区间出现最大值。瓶插8 d时,从数值上可以看出干重相比0 d略微下降,但是变化较小,但是在5~20 cm区域有明显的下降趋势;鲜重下降幅度较干重更明显,特别是在5~20 cm区域。同时,干、鲜重仍然在花茎远离花盘的区域,特别是25~30 cm保持较高值。干、鲜重在花茎上的这种不均匀分布可能导致了花茎的支持力存在区域差异性,导致弯茎往往发生在花茎的特定区域。
瓶插期间非洲菊的各个茎段的鲜重均呈现不同程度的损失,鲜重损失率根据不同品种和不同区域存在差异。供试10个非洲菊品种鲜重损失率较高的茎段主要集中在距花盘基部5~20 cm的范围。其中,10~15 cm区域的鲜重损失率最高,从该区域向两侧逐渐递减(表3),该区域也和结果1中弯茎发生位置7~13 cm相吻合。同时,各非洲菊品种花茎的干物质含量也发生了轻微的变化(表4)。总体而言,在瓶插过程中干物质的变化较小,呈不变或轻微减少的趋势。干重损失率最高的区域仍然集中在距花盘基部5~20 cm区域,除了‘金太阳,其余品种在10~15 cm区域的干重损失率最高,并从两侧递减。由于水分占据了茎干鲜重的85%~90%左右,鲜重的减少说明在瓶插过程中水分损失较多。对各个品种失水率的检测发现,失水率在不同茎段间的变化趋势和鲜重变化类似(表5)。由此可得,水分的损失可能导致茎干5~20 cm区域,特别是10~15 cm区域膨压下降,从而造成该区域的机械支持力度减少,发生弯茎现象。
然而,不弯茎品种‘白马王子和‘鸿运,在茎干5~20 cm区域也出现了较高的干、鲜重和水分损失,尤其是‘鸿运的干、鲜重和水分损失甚至超过很多易弯茎品种。该结果说明品种间弯茎现象的差异并非由茎干的水分代谢决定,但是至少在一定程度上决定了弯茎发生的区域。
2.4 非洲菊花茎伸长量对弯茎的影响
非洲菊切花在采后瓶插过程中花茎会出现再伸长的现象,为了探讨花茎伸长对非洲菊切花弯茎的影响,挑选易弯茎品种‘水粉和‘黄绿心,及不弯茎品种‘白马王子和‘鸿运,分别测定了各品种瓶插前4 d花茎的伸长量。结果发现,瓶插前4 d,非洲菊各品种距离花盘基部10~20 cm区域伸长量较少,而在0~10 cm区域均存在一定程度的再伸长,伸长程度根据品种和茎段区域的不同而存在一定差异。其中,伸长最多的是0~5 cm段,其次是5~10 cm段。同时,通过易弯茎与不弯茎品种各茎段伸长量的比较,发现易弯茎与不弯茎品种0~5 cm,5~10 cm茎段伸长量只与品种相关,与弯茎发生不存在联系(图5),说明花茎的再伸长和弯茎现象没有必然联系。
3 讨论
非洲菊是重要的切花材料,然而其采后常常发生弯茎现象,是非洲菊切花采后保鲜的世界性难题。探索非洲菊采后弯茎的发生机制,对于延长非洲菊切花采后瓶插寿命,提高观赏价值,具有重要的研究意义。本试验研究结果表明,切花非洲菊的弯茎现象存在品种差异性,不同品种的弯茎发生时间和弯茎率存在较大差异,这与国内外有关研究结论类似[4-5]。同时,弯茎发生的时间也受到气候、季节、切花批次和切花状态的影响,如比较表1和表2各品种的弯茎发生时间,发现同一个品种的弯茎发生时间略有不同,但是这种差异只在一定范围内变化。此外,供试非洲菊弯茎发生位置与折断位置不一样,弯茎开始发生位置一般在距花冠基部7~9 cm茎段处,而弯茎最终折断位置在11~13 cm处,这个现象可能是由于花茎不同区域具有不同的组织结构,以及花盘重量引起的机械力相关。有关学者也提出不同非洲菊品种弯茎的发生区域略有不同,通常集中在8~15 cm 的区段[21]。另外,供试10个非洲菊品种中花茎直径大小并不影响弯茎现象的发生。与Perik等[2]研究结论类似。
目前,关于乙烯是否影响非洲菊切花的衰老进程仍然存在分歧。部分研究认为非洲菊对乙烯不敏感,外源乙烯处理对非洲菊切花的衰老影响不大[14,17-18]。也有报道指出非洲菊切花对乙烯的敏感性与品种有关,并分为敏感,略敏感和不敏感品种[19]。同时,另一些研究发现,乙烯对于非洲菊弯茎具有抑制作用,可能与促进木质素合成相关[4,22],增加了花茎的坚固性,减少弯茎的发生。本试验结果表明非洲菊不同品种对乙烯也表现为敏感与不敏感,对于乙烯敏感型非洲菊品种乙烯对其弯茎进程存在促进或抑制作用。类似的结果在月季中也有发现[23]。因此,针对特定品种的乙烯敏感性和响应机制仍然需要进一步研究来探索。
弯茎现象的产生,与花茎的组织结构[1-2,5]、花茎中物质和水分的新陈代谢密切相关[2,13,14]。有研究表明,在非洲菊花茎中,花头以下的0~15 cm的区域几乎不含厚壁组织,木质素的含量从花头到花茎基部逐渐增加,与厚壁组织的分布一致[2];廖立新等[3]也发现非洲菊弯茎区域和上部茎段粗纤维含量无显著差异,但都远低于下部,并且弯茎发生部位的水分净损失会显著高于其他区段;另外,已有报道显示,非洲菊切花在瓶插阶段的水分净损失几乎只限于花茎,也是弯茎现象产生的原因之一[9,24]。本试验研究结果显示,瓶插期间,非洲菊各品种花茎的干、鲜重和水分变化在不同区域并不相同。干重含量整体变化较小,但是在弯茎区域有明显的下降趋势,鲜重和水分损失变化类似,均表现为10~15 cm处茎段损失率最高,也与弯茎区域吻合。由此可见,非洲菊切花瓶插期间茎干弯茎区域的干物质损耗和水分散失均高于其它区域,可能是导致弯茎发生的主要原因之一。
然而,研究结果并没有发现干、鲜重损耗与失水率在弯茎品种和不弯茎品种之间存在显著差异,Perik等[2]研究发现,同一个品种内弯茎个体和不弯茎个体的花茎鲜重损失率,在所有区域都存在显著差异,即弯茎个体的花茎失水率高于不弯茎个体。结合这个观点,推测非洲菊的弯茎现象,特别是品种间的弯茎差异是一个由多种因素共同协作影响的生理现象,包括花茎组织结构特点,花茎中水分和相关物质代谢,内外因素引起的花茎导管堵塞和植物激素调控等。因此,结合以上研究结果,下一步研究可以集中在其他植物激素,如赤霉素、细胞分裂素、脱落酸对于非洲菊弯茎发生的作用;导管堵塞的成因,如细菌性还是生理性堵塞;探讨各影响因素之间的互作关系等。不同的研究结果是否和品种特异性有关,还需要进一步研究证明。此外,关于非洲菊弯茎发生的分子机理仍然不清楚,今后的研究方向在非洲菊弯茎发生的生理研究基础上,逐步开展相关因素的分子机理研究,对于开发非洲菊切花保鲜剂具有重要的理论价值和实际生产意义。
参考文献
[1] 冯 会. 非洲菊弯茎机理及保鲜技术研究[D]. 福州: 福建农林大学, 2006.
[2] Perik R R J, Razé D, Harkema H, et al. Bending in cut Gerbera jamesonii flowers relates to adverse water relations and lack of stem sclerenchyma development, not to expansion of the stem central cavity or stem elongation[J]. Postharvest Biol Tech, 2012, 74: 11-18.
[3] 廖立新, 彭永宏, 叶庆生. 非洲菊鲜切花弯茎部位及有关原因[J]. 园艺学报, 2003, 30(1): 110-112.
[4] Ferrante A, Alberici A, Antonacci S, et al. Effect of promoter and inhibitors of phenylalanine ammonia lysase enzyme on stem bending of cut gerbera flowers[J]. Acta Hortic, 2007, 755: 471-476.
[5] 吕长平, 陈海霞, 陈晨甜. 切花非洲菊花茎解剖构造对瓶插寿命的影响研究[J]. 中国农学通报, 2011, 27(31): 99-104.
[6] Penningfeld F, Forchthammer L. Silbernitrat verbessert die Haltbarkeit geschnittener Gerbera[J]. Gartenwelt, 1966, 66: 226-228.
[7] van Meeteren U. Water relations and keeping quality of cut Gerbera flowers.I. The cause of stem break[J]. Scientia Hortic, 1978(8): 65-74.
[8] van Meeteren, U. Water relations and keeping quality of cut Gerbera flowers. II. Water balance of ageing flowers[J]. Scientia Hortic, 1978(9): 189-197.
[9] Steinitz B. The role of sucrose in stabilization of cut gerbera flower stalks[J]. Gartenbauwissenschaft, 1982, 47: 77-81.
[10] Steinitz B. The influence of sucrose and silver ions on dry weight, fiber and lignin content, and stability of cut Gerbera flower stalks[J]. Gartenbauwissenschaft, 1983, 48: 821-837.
[11] Dubuc-Lebreux M A, Vieth J. Histologie du pédoncule inflorescentiel de Gerbera jamesonii[J]. Acta Bot Neerl, 1985, 34: 171-182.
[12] Marousky F J. Vascular structure of the Gerbera scape[J]. Acta Horticulture, 1986, 181: 399-406.
[13] Redgwell R J, MacRae E, Hallett I, et al. In vivo and in vitro swelling of cell walls during fruit ripening[J]. Planta, 1997, 203(2): 162-173.
[14] 吴岚芳, 黄绵佳, 蔡世英. 非洲菊切花活性氧代谢的研究[J]. 园艺学报, 2003, 30(1): 69-73.
[15] Mencarelli F, Agostini R, Botondi R, et al. Ethylene production, ACC content, PAL and POD activities in excised sections of straight and bent gerbera scapes[J]. J Hortic Sci Biotech, 1995 70(3): 409-416.
[16] Emongor V E. Effects of gibberellic acid on postharvest quality and vaselife life of gerbera cut flowers(Gerbera jamesonii). Journal of Agronomy, 2004, 3(3): 191-195.
[17] 高俊平. 切花衰老和乙烯. 园艺学年评[M]. 北京: 科学出版社, 1995: 82-106.
[18] 王凤兰, 张昭其. 切花花瓣衰老机理研究进展[J]. 浙江农业学报, 2011, 23(5): 1 063-1 068.
[19] 盛爱武, 谢晓蓉, 曹 丽. 非洲菊切花品种的乙烯敏感性及保鲜研究[J]. 仲恺农业技术学院学报, 2003, 16(2): 26-31.
[20] 陈丹生, 李娘辉, 王精明, 等. 氯化钙对非洲菊切花的保鲜作用[J]. 云南植物研究, 2004, 26(3): 345-348.
[21]李金泽, 熊 丽, 李 鹏. 非洲菊的瓶插寿命及其保鲜技术[J]. 西南园艺, 2003(1): 19.
[22] Gerasopoulos D, Chebli B. Effects of scape-injected 1-aminocyclopropane-1-carboxylic acid(ACC)on the vase life of 'Testarossacut gerberas[J]. J Am Soc Hortic Sci, 1998, 123: 921-924.
[23] 蔡 蕾, 张晓红, 沈红香, 等. 乙烯对不同切花月季品种开花和衰老的影响[J]. 园艺学报, 2002, 29(5): 467-472.
[22] Bleecker A B, Kende H. Ethylene: A gaseous signal molecule in plants[J]. Annu Rev Cell Dev Biol, 2000, 16: 1-18.
[24] 王 晰, 徐 哲,赖齐贤, 等. 非洲菊切花弯茎影响因素研究进展[J]. 园艺学报, 2015, 42(9): 1 771-1 780.
猜你喜欢
中国石油石化(2021年8期)2021-07-20
水泵技术(2021年4期)2021-01-22
石油化工技术与经济(2021年4期)2021-01-13
石油化工建设(2018年2期)2018-07-11
流程工业(2017年4期)2017-06-21
中国塑料(2016年12期)2016-06-15
合成化学(2015年2期)2016-01-17
焊接(2015年3期)2015-07-18
中国氯碱(2014年7期)2014-08-15
自动化博览(2014年9期)2014-02-28
