
水分胁迫对金露梅叶片水势、光合特性和水分利用效率的影响
2016-06-18 07:12刘颖贺康宁徐特王辉刘玉娟
中国水土保持科学 2016年1期
刘颖,贺康宁,徐特,王辉,刘玉娟
(北京林业大学 水土保持与荒漠化防治教育部重点试验室, 100083, 北京)
水分胁迫对金露梅叶片水势、光合特性和水分利用效率的影响
刘颖,贺康宁†,徐特,王辉,刘玉娟
(北京林业大学 水土保持与荒漠化防治教育部重点试验室, 100083, 北京)
摘要:为解决青海高寒区绿化植物的栽培驯化问题,并为金露梅栽植的科学管理及高寒区水资源有效利用提供理论基础,以2年生金露梅幼苗为研究对象,通过盆栽方法人为控制土壤水分条件,测定不同土壤水分条件下金露梅苗木的光合生理特征,研究金露梅光合生理特性及其与土壤水分的相互关系。结果显示:1)金露梅凋萎系数为4.02%左右;2)土壤水分对金露梅叶水势及光合作用的影响具有阈值现象,净光合速率最大时土壤含水量为20.83%,水合补偿点为4.38%,水分利用效率最大时土壤含水量为13.82%;3)在砂壤土条件下,金露梅生长最适宜的土壤水分环境为8.33%~12.71%,此范围内既可以维持植物基本的生长所需,又可以最大程度提高水分利用效率;4)青海地区在金露梅盛花期(7月),在无降雨的情况下,每2~3周补充一次水分能维持金露梅较好生长。2~3周的持续干旱所造成的伤害在灌水后可逐渐恢复,但连续1个月无任何供水会使金露梅死亡。
关键词:水分胁迫; 叶片水势; 光合作用; 水分利用效率; 土壤含水量
我国城市化建设的快速发展以及新农村建设的推进,对各地道路绿化、园林景观都提出了更高的要求[1];但青海地区纬度高、气温低以及干旱问题[2]使得许多低海拔地区的花卉植物无法在当地恶劣的气候中生长与繁殖[3]。许多学者开始针对当地优势野生物种进行研究,试图从当地花卉中挑选出适合园林美化与道路绿化的植物品种进行驯化培养,以满足青海地区对绿化植物的迫切需求。
金露梅(Potentillafruticosa)具有抗寒性和耐旱性,是青海高海拔区一种野生优势物种,花期可持续5个月,开黄色小花,花型与颜色俱佳,可引种为城镇绿化植物。金露梅属蔷薇科委陵菜属灌木,C3植物,产于青海、甘肃、四川及云南,东北、华北也有分布,生于海拔2 500~4 000 m的山坡草地、砾石坡、灌丛及林缘或河滩上[4],观赏性良好,被誉为“花寿之王”,是极具开发价值的野生绿化花卉资源。目前金露梅由野生向城市道路绿化的引种已在甘肃等地取得进展[5],已有金露梅用于绿篱或草坪、花境边缘,以及配植于高山园、岩石园的报道[6],在青海柴达木地区的道路绿化中也可见金露梅的分布。但现阶段金露梅对干旱胁迫及复水的响应规律尚不明晰,造成管理水平低下,不利于金露梅在干旱半干旱地区的灌溉管理及推广,且以往研究多集中在干旱期间植物的忍耐程度上,对胁迫后复水条件下植物的恢复与补偿现象认识有限[7]。笔者以金露梅幼苗为研究对象,通过盆栽方法人工模拟土壤干旱条件,测定不同土壤水分含量、不同干旱及复水历时条件下叶水势和光合参数的变化,揭示金露梅干旱耐受能力和供水后的恢复能力,为提高金露梅的管理水平以及青海高寒区植被建设提供参考。
1研究区概况
本试验地点位于青海省中部海西蒙古族藏族自治州都兰县境内的香日德农场(E 97°48′12″,N 36°04′39″ )。该农场位于柴达木盆地东南边缘,海拔2 997 m,属于高原大陆性气候,空气干燥,辐射强烈,干旱、霜冻、风沙等灾害性气候居多。年平均气温4 ℃,年较差26.1~28.0 ℃。此地区降雨稀少,年降水量166 mm,年蒸发量2 227 mm(20 cm蒸发器),无霜期90 d左右。土壤多数为薄土层或沙砾石滩,以沙壤土为主,质地为壤质。当地野生植被大多数属于藜科、菊科、豆科、蒺藜科,且都是耐旱、耐盐碱的物种,如梭梭(Haloxylonammodendron(C.A. Mey.) Bunge)、柽柳(TamarixchinensisLour.)、白刺(NitrariatangutorumBobr.)等。
2研究方法
2.1试验设计
2012年4月,将供试金露梅苗木(1年生)植入上口径30.5 cm,下口径28.5 cm,高32.0 cm的侧向打孔排水的塑料圆筒中,每盆1株。苗木栽植后,充分浇水,使之成活并培养1年。试验盆内土壤为24.6 kg砂壤土,土壤密度1.42 g/cm3,田间持水量24.1%,pH值8.2。
7月为金露梅在青海柴达木地区的盛花期,故于2013年7月1日开始,选取株高(45.0~50.0 cm)、冠幅(48.0 cm×40.0 cm左右)、枝条数(13~15)基本一致的17盆苗木进行干旱及复水试验。试验期间,晴天时,使盆栽暴露在自然环境中,阴天和雨天时,将花盆移入透明塑料防雨棚来阻碍自然降水。将试验苗木分为对照CK组(5盆)和处理W组(12盆)。利用TDR土壤水分测定仪每日测量各盆植物土壤体积含水量(在花盆中部接近苗木根部位置的不同方向上测量3次),之后换算为质量含水量(本研究讨论的土壤含水量均为花盆内土壤的质量含水量)。
干旱胁迫试验:控制CK组土壤含水量维持在23.0%~24.0%,W组在充分供水后自然干旱。因为叶片水势在清晨最为稳定,净光合速率在11:00时处于上升阶段;故分别于胁迫试验开始后每3 d对W组中生长状态良好的6盆植物和CK中生长状态良好的3盆植物测量清晨叶水势及11:00—12:00间的光合生理参数,并每6 d对W组中生长状态良好的6盆植物测量叶水势及光合生理参数的日变化过程。
复水试验:当处理组植物出现净光合速率趋近于零的时候,对W组中3盆植物进行复水,使其土壤水分含量维持在对照组水平,观察复水后植物的生长状况,其余9盆继续干旱胁迫试验;3 d后再对W组中3盆植物进行复水(设为D组),并以此类推每3 d对余下的3盆植物进行复水,至所有植物都复水完毕,复水组分别设置为D、E、F。植物复水后每3 d测量1次W组植物清晨叶水势及11:00—12:00间的光合生理参数,并于试验第30 d测定W组植物的叶水势及光合生理参数的日变化过程。试验测定时均为晴朗天气,日照充足。
2.2测量方法
使用PSYPRO露点水势仪于06:00—07:00间测定植物清晨叶水势(ψw),选择发育良好的枝条上部功能叶片,放入C-52样品室,每5 min记录1次数据,维持2 h,测量结果取平均值。
利用LI-COR 6400便携式光合仪(LICOR, Lincoln, NE,USA)对植物叶片进行光合参数的观测,时间为11:00—12:00。每盆植物选中上部5~6片健康叶子,每次连续记录10个稳定数据,测量结果取平均值。可得被测植物的净光合速率、蒸腾速率、气孔导度和胞间CO2浓度等光合生理参数。
W组与CK叶水势和光合参数日过程的观测时间为07:00—19:00,每隔2 h观测1次数据。日过程试验第1、9、18、30天的W组的数据分别为W1、W2、W3、Wf。
使用SPSS18.0 和Excel 软件进行数据分析。
3结果与分析
3.1干旱和复水过程中土壤含水量、叶片水势、蒸腾作用的变化

CK is the comparison group; W is the test group; D, E and F refer to the the rewatering group in the 21th, 24th, 27th day. 图1 土壤含水量变化Fig.1 Changes of soil moisture content
如图1所示在试验过程中CK组的土壤含水量始终保持在24.1%左右;而随着自然干旱的进行,W组的土壤含水量急剧下降,至第15天时植物叶片出现发黄、萎蔫现象;到试验的第18天土壤含水量降为5.97%,降幅达78.58%,一半以上的叶片已萎蔫与干枯。在第18天晚上对W组进行复水,使其土壤含水量基本达到CK组水平,复水12 d后5盆植物均有新叶芽长出;第21天晚上将W组中的D组进行复水,复水前约80%的叶片干枯,复水后9 d仅有1盆出现新的叶芽;第24天晚上将W组中的E组进行复水,复水前所有小枝和叶片干枯,复水后6 d生长状况无恢复;第27天晚上将W组中的F组进行复水,复水前土壤含水量降至3.27%,复水至CK组水平多日后生理活性无恢复。
如图2(a)所示,CK组与W组起始清晨叶水势均为-0.76 MPa左右,当自然干旱进行到第3天时,W组的清晨叶水势并未大幅下降;继续干旱至第6天开始明显下降,最大降幅出现在第6~9天之间;从第9天开始叶水势平稳下降至第18天达最低点-3.78 MPa。可知当水分下降至阈值19.48%左右时叶水势才会开始下降。干旱18 d后因叶片干枯萎蔫现象严重,故D、E、F组苗木未能继续进行叶水势及光合参数的测定。复水后第1天W组叶水势迅速大幅回升,此后随复水历时的增长趋于稳定,其值略低于CK,且没有超过自身起始值,未出现补偿现象。
如图2(b)所示,叶水势在清晨最高,此后不断降低至15:00出现最低值,之后回升但仍低于清晨叶水势。水分胁迫下,叶水势不断下降,在土壤含水量14.37%~17.03%之间,清晨叶水势下降幅度最大速度最快;但日变化规律保持稳定,当土壤水分含量下降至5.97%时,叶水势全天变化范围极小,变化规律紊乱,生理活性受到严重抑制,复水后日变化规律恢复。
如图2(c)所示,干旱处理前6 d蒸腾速率略有上升,至第9天开始大幅下降,第18天达最低值,降幅为80.55%。复水后1 d蒸腾速率即出现明显回升,复水后12 d恢复到CK水平。如图2(d)所示,蒸腾速率的日变化规律表现为“双峰型”,2个高峰值出现在11:00与15:00,13:00出现暂时降低的“午休现象”。随土壤水分含量降低,蒸腾速率也不断下降,至第18天降至最低值,蒸腾微弱,蒸腾曲线的变化幅度极小;复水后蒸腾曲线的变化幅度加大,变化规律恢复。
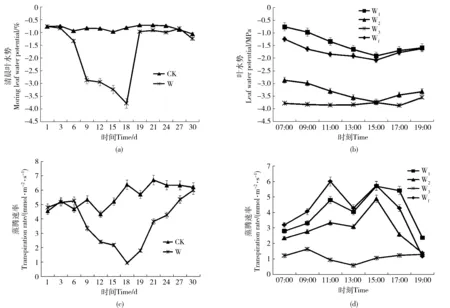
W1,W2,W3 and Wf refers respectively to the data of the darly experiment in group W on the 1st, 9th, 18th and 30th day图2 叶水势、蒸腾速率变化Fig.2 Changes of leaf water potential and transpiration rate
苗木蒸腾量的变化和凋萎湿度之间有着密切的关系,当土壤含水量降低至苗木吸水受到胁迫时,就会使苗木失去水分平衡,因而代谢作用失常,出现紊乱状态,开始永久凋萎,同时可用叶水势校核苗木的凋萎点[8]。金露梅在土壤含水量为5.97%时,蒸腾速率与叶水势的日变化规律都出现紊乱;土壤含水量降至4.34%时,复水后半数苗木无法恢复生理活性,已经死亡;土壤含水量低至4.02%左右时,复水后所有苗木无恢复,已经死亡,可得金露梅凋萎含水量为4.02%左右。
3.2干旱和复水过程中光合特性的变化
从图3(a)可看出:试验过程中CK组净光合速率因环境因素变化影响略有起伏;干旱处理前3 d,净光合速率呈上升趋势;W组第9天净光合速率仍高于CK组,未出现大幅下降,此后随土壤含水量下降净光合速率不断减小,即土壤含水量14.37%是净光合速率出现大幅改变的阈值。复水后1 d净光合速率出现微弱回升,其值与复水前基本持平,说明光合作用的恢复有一定的滞后;复水后3 d回升明显;至复水后9 d净光合速率恢复为CK水平,12 d时W组净光合速率超过CK组与自身起始值,出现一定的补偿现象。
由图3(b)可知,金露梅净光合速率也呈“双峰型”日变化规律,于11:00上升至峰值,13:00降至最低值,15:00再次升至峰值。水分含量降至5.97%时,净光合速率全天维持在极低水平,变化曲线起伏微弱,日变化幅度较小。说明此时光合作用已经受到严重抑制,生理活动出现紊乱,复水后日变化曲线恢复“双峰型”。

图3 净光合速率、胞间CO2浓度、气孔导度变化Fig.3 Changes of net photosynthetic rate, intercellular CO2 concentration and stomatal conductance
气孔是连接植物体内与外部大气的通道,在一定程度上控制着水分与CO2的进出;同时气孔的开闭反之也受CO2浓度的影响:高CO2浓度使叶片气孔关闭,低CO2浓度促使气孔开启[9]。从图3(c)和图3(d)可知试验过程中CK组气孔导度受多方面因素影响出现起伏现象,干旱W组气孔导度在前9 d都高于CK组,12 d时气孔导度才开始大幅下降。第18天,由于土壤水分含量已降至最低,为了最大程度减少蒸腾耗水,气孔导度降至0.01 mol/(m2·s),接近于完全关闭。复水12 d后气孔导度持续上升至对照水平,但未超过自身起始值。气孔导度在叶水势降低至阈值时下降,且气孔导度近于零时的叶水势值越低,抗旱性越大[10]。图2(a)所示金露梅气孔导度开始降低的土壤含水量阈值为14.37%,水势阈值为-2.87 MPa;气孔接近完全关闭时土壤含水量阈值为5.97%,水势阈值为-3.78 MPa。
3.3干旱和复水过程中水分利用效率的变化
植物的水分利用效率是由净光合速率与蒸腾速率的比值计算而得。在干旱环境下,水分利用效率的大小决定了植物节水能力和水分生产力水平[11]。由图4可知:干旱胁迫前6 d水分利用效率变化不大,W组与对CK组相差无几;随土壤含水量的继续下降,第9天时W组水分利用效率大幅增长,涨幅达35.17%,比CK组高95.0%。第12天用水效率再次下降,至复水后第3天降至最低,复水后3 d仍保持在较低水平,说明水分利用效率的恢复有较长的滞后性;复水后6 d水分利用效率回升超过CK组15.97%,出现补偿现象,可见金露梅对水分胁迫有较强的忍耐度,且经过干旱锻炼后其用水效率可一定程度地提高。
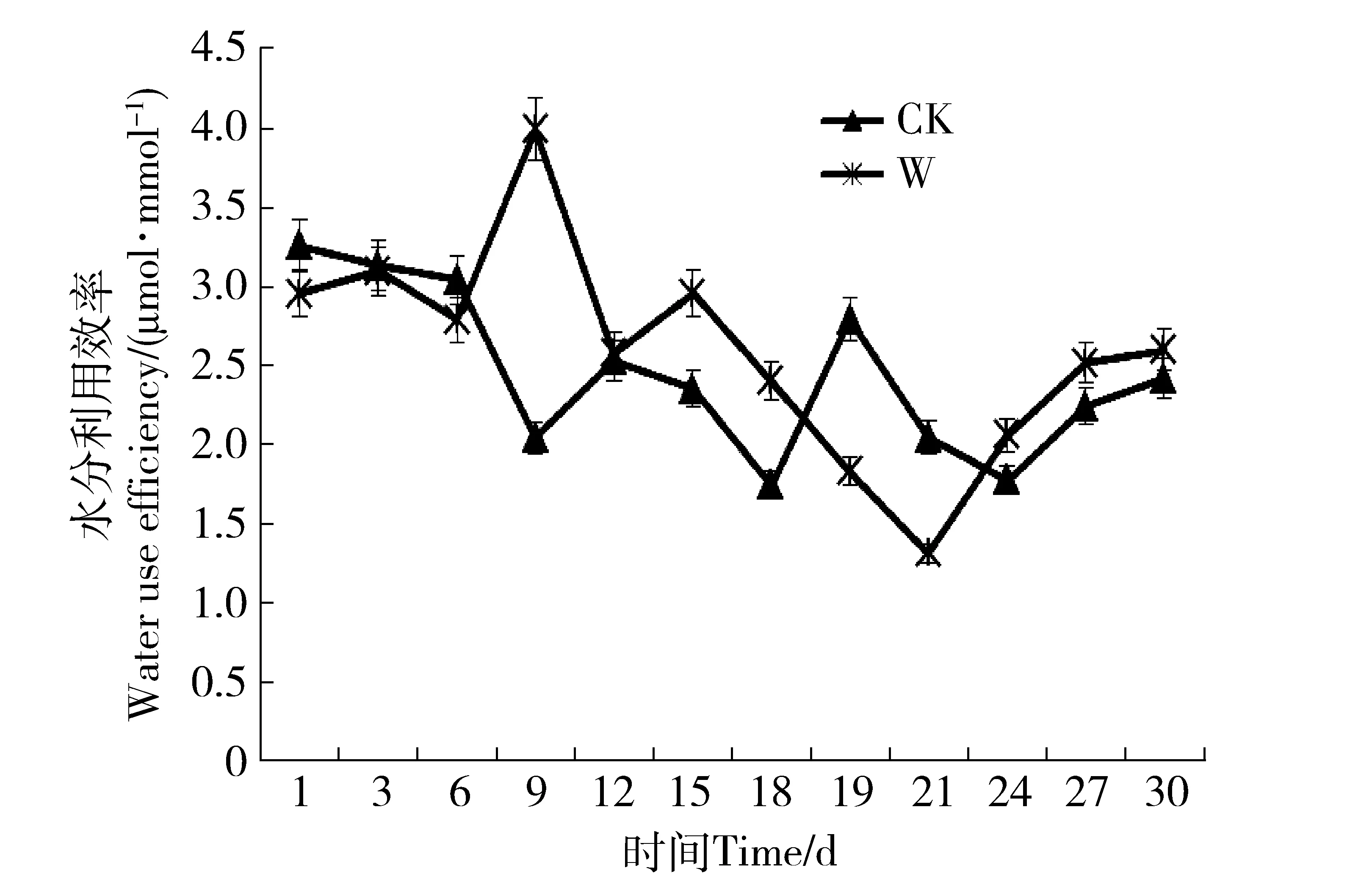
图4 水分利用效率变化Fig.4 Changes of water use efficiency
3.4净光合速率、蒸腾速率、水分利用效率与土壤水分的关系
图5(a)、图5(b)所示干旱胁迫期间金露梅净光合速率和蒸腾速率与土壤含水量均成二次曲线关系。由计算可知:金露梅净光合速率最大值对应的土壤含水量为20.83%,水合补偿点为4.38%,维持最高净光合速率70%以上的土壤含水量应在12.71%以上;蒸腾速率最大值对应的土壤含水量为25.52%,蒸腾速率为0时对应的土壤含水量为2.81%,维持最高蒸腾速率70%以上的土壤含水量应在13.09%以上。
如图5(c)所示在设定土壤水分范围内,金露梅水分利用效率随土壤含水量上升先增大后减小,最后趋于稳定。在土壤水分含量13.82%时出现峰值,7.20%~21.9%之间可维持水分利用效率的70%以上,变化速率由快变缓的拐点为含水量8.33%。已有研究证明,山西方山主要树种刺槐(Robiniapseudoacacia)、侧柏(Platycladusorientalis)和苹果(Maluspumila)的水分利用效率随土壤含水量的变化呈三次四项式的关系[12],具有一定的阈值现象,与笔者研究结果一致。
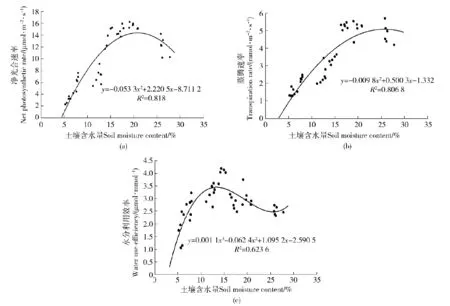
图5 净光合速率、蒸腾速率、叶片水分利用效率与土壤水分的相互关系Fig.5 Relationships between net photosynthetic rate, transpiration rate, water use efficiency and soil moisture content
因此,选取维持水分利用效率较快增长的土壤含水量阈值8.33%和维持较高净光合速率的土壤含水量临界值12.71%,作为干旱区金露梅生长适宜的土壤水分环境的低限和高限值。
4讨论和结论
叶水势反应出叶片水分状况的变化,其值越低说明植物吸水能力越好,抗旱性就越强[13]。在一定阈值下,金露梅叶水势随土壤水分降低开始下降,而净光合速率、蒸腾速率和气孔导度随干旱历时延长和土壤水分降低均表现为先升高后下降的趋势。
水分变动条件下,植物的生长存活和产量不仅仅取决于对阶段干旱的忍耐能力,还决定于对旱后复水的适应和恢复能力[14]。植物对干旱缺水有一定的适应范围,在该范围内的缺水,往往在复水后,可产生水分利用和生长上的补偿效应,对终产量无害甚至有利,这就是植物的有限缺水效应[15]。郭贤仕等[16]发现,前期干旱锻炼使复水后谷子叶片的净光合速率超过一直充分供水的对照,并认为这与叶片的叶绿素含量增加和渗透调节能力增强有关。同样有研究[17]证明复水后各小麦品种净光合速率、气孔导度、蒸腾速率都迅速向正常供水水平接近,气孔导度恢复最迅速、超补偿效应明显,净光合速率次之,蒸腾速率最慢;但笔者得出了不同的结果,蒸腾速率、气孔导度、净光合速率在复水1 d后开始回升,蒸腾速率恢复地最迅速,气孔导度次之,净光合速率表现为相对滞后。这进一步说明金露梅蒸腾作用对水分改变的敏感性比光合作用高,这可能是因为蒸腾作用受气孔导度影响更大。
很多研究[11]认为水分利用效率并非在水分充足时最高,由于蒸腾作用对水分胁迫的反应比光合作用敏感,使得光合速率的降低比蒸腾速率滞后;因此适度的水分胁迫,可使水分利用效率有所提高。特别是在临界发育阶段,中期产量可能受水分亏缺影响的时候,这种适应性的调节方式对植物的节水与生存是有利的[18]。
通过分析模拟土壤干旱条件下及复水后盆栽金露梅叶水势与光合参数的变化特征,基于笔者试验的土壤类型以及气候类型,得出如下结论:
1)金露梅凋萎系数为4.02%左右。
2)土壤含水量对金露梅叶水势及光合作用的影响具有一定的阈值现象,过高与过低的水分均会抑制金露梅的各项生理活性。金露梅叶水势随土壤水分降低开始下降的阈值为19.48%左右;在土壤含水量14.37%~17.03%之间,清晨叶水势下降幅度最大速度最快,说明此范围内土壤水分含量会明显提高金露梅的吸水性及抗旱性。
金露梅气孔导度开始降低的土壤含水量阈值为14.37%,水势阈值为-2.87 MPa;干旱历时18 d气孔接近完全关闭时土壤含水量阈值为5.97%,水势阈值为-3.78 MPa。
净光合速率最大时土壤含水量为20.83%,水合补偿点为4.38%,维持最高净光合速率70%以上的土壤含水量最低为12.71%;水分利用效率最大时土壤含水量为13.82%,维持水分利用效率70%以上且变化速率由快变缓的拐点为土壤含水量8.33%。
3)在砂壤土条件下,金露梅生长最适宜的土壤水分环境为8.33%~12.71%,此范围内既可以维持植物基本的生长所需,又可以最大程度提高水分利用效率,对干旱区水资源的最大化利用极具意义。
4)青海地区在金露梅盛花期(7月),无降雨的情况下,2~3周浇1次透水能维持金露梅较好的生长状况,2~3周的持续干旱所造成的伤害在灌水后可逐渐恢复,但连续1个月无任何供水会使金露梅死亡。
5参考文献
[1]孙飞. 西北干旱区城市森林道路绿化植物配置模式研究[D].兰州:甘肃农业大学, 2013:37-38.
Sun Fei.Research on the mode of plant configurationof urban forest road in Northwest arid areas[D].Lanzhou: Gansu Agriclutural University, 2013:37-38. (in Chinese)
[2]李芳兰, 包维楷, 吴宁. 白刺花幼苗对不同强度干旱胁迫的形态与生理响应[J]. 生态学报, 2009,30(19):5140.
Li Fanglan, Bao Weikai, Wu Ning. Morphological and physiological responses of current Sophora davidii seedlings to drought stress[J]. Acta Ecologica Sinica, 2009, 30(19):5140. (in Chinese)
[3]董梅. 柴达木地区主要树种抗旱耐盐生理研究[D]. 北京:北京林业大学, 2013:23-25.
Dong Mei. Drought and Salinity Resistant Physiology of main tree speciesin Qaidam of China[D]. Beijing: Beijing Forestry University, 2013:23-25. (in Chinese)
[4]祁如雄. 青南牧区野生金露梅、银露梅人工驯化研究[J]. 现代农业科技, 2008(18):32.
Qi Ruxiong. Study on wild Potentilla fruticosaandPotentilla glabrain pasturing area[J].Modern Agricultural Science and Technology, 2008(18):32. (in Chinese)
[5]王东武. 金露梅在甘肃地区园林绿化中的应用[J]. 中国园艺文摘, 2013(9):83.
Wang Dongwu. The application of Potentilla fruticosa in landscaping in Gansu area[J]. Chinese Horticulture Abstracts, 2013(9):83. (in Chinese)
[6]方正, 王占朝, 陈段芬,等. 不同激素处理对火鹤开花的影响初探[J]. 河北农业大学学报, 2004,27(2):51.
Fang Zheng, Wang Zhanchao, Chen Duanfen, et al. Effect of different hormone on blossom of Anthurium scherzerianum[J]. Journal of Agricultural University of Hebei, 2004,27(2):51. (in Chinese)
[7]王磊, 张彤, 丁圣彦. 干旱和复水对大豆光合生理生态特性的影响[J]. 生态学报, 2006,26(7):2073.
Wang Lei, Zhang Tong, Ding Shengyan. Effect of drought and rewatering on photosynthetic physioecological characteristics of soybean[J]. Acta Ecologica Sinica, 2006,26(7):2073. (in Chinese)
[8]王斌瑞. 晋西黄土高原主要造林树种凋萎湿度的研究[J]. 北京林业大学学报, 1988,10(4):17.
Wang Binrui. Study on the loess of wilting moisture of main afforestation tree species[J].Journal of Beijing Forestry University, 1988,10(4):17. (in Chinese)
[9]Kaufmann M R. Leaf conduce ance as a function of photosynthetic photon flux density and absolute humidity difference from leaf to air[J]. Plant physiology, 1982,69(4):1018.
[10] 刘友良. 植物水分逆境生理[M].北京:中国林业出版社, 1992: 79-83.
Liu Youliang.Plantphysiology under water stress[M]. Beijing:Chinese forestrypublishing company, 1992: 79-83. (in Chinese)
[11] 陈静, 秦景, 贺康宁,等. 水分胁迫对银水牛果生长及光合气体交换参数的影响[J]. 西北植物学报, 2009,29(8):1649.
Chen Jing, Qin Jing, He Kangning, et al. Influences of water-stress on shepherdia argentea seedlingsgrowth and photosynthetic gas-exchange parameters[J].Acta Botanica Boreali-Occidentalia Sinica, 2009,29(8):1649. (in Chinese)
[12] 贺康宁, 张光灿, 田阳,等. 黄土半干旱区集水造林条件下林木生长适宜的土壤水分环境[J]. 林业科学, 2003,39(1):10.
He Kangning, Zhang Guangcan, Tian Yang, et al. The suitable soil moisture condition for forest growthin catchment afforestation in semi-arid regionon loess plateaus[J].Scientia Silvae Sinicae, 2003,39(1):10. (in Chinese)
[13] 李继文, 王进鑫, 张慕黎. 干旱及复水对刺槐叶水势的影响[J]. 西北林学院学报, 2009,24(3):33.
Li Jiwen , Wang Jinxin , ZhangMuli. Effect of drought and rewater on leaf water potential of Robinia pseudoacacia[J].Journal of Northwest Forestry University, 2009,24(3):33. (in Chinese)
[14] McCree K J. Whole-plant carbon balance during osmotic adjustment to drought and salinity stress[J]. Australian Journal of Plant Physiology, 1986,13(8):33.
[15] 厉广辉, 万勇善, 刘风珍. 苗期干旱及复水条件下不同花生品种的光合特性[J]. 植物生态学报, 2014,38(7):729.
Li Guanghui, Wan Yongshan, Liu Fengzhen.Photosynthetic characteristics in different peanut cultivars under conditions of drought andre-watering at seedling stage[J].Chinese Journal of Plant Ecology, 2014,38(7):729. (in Chinese)
[16] 郭贤仕, 山仑. 前期干旱锻炼对谷子水分利用效率的影响[J]. 作物学报, 1994,20(3):352.
Guo Xianshi,Shan Lun.The influence of early drought exercise on millet water use efficiency[J]. Acta Agronomica Sinica, 1994,20(3):352. (in Chinese)
[17] 罗爱花, 郭贤仕, 柴守玺. 水分胁迫对不同品种春小麦的水分利用率及生理指标的影响[J]. 2008,25(5):683.
Luo Aihua, Guo Xianshi, Chai Shouxi. Effects of water stress on water use efficiency and physiological traits of different varieties of spring wheat[J].2008,25(5):683. (in Chinese)
[18] 许大全. 光合作用“午睡”现象的生态、生理与生化[J]. 植物生理学报, 1990(6):5.
Xu Daquan. Ecology, physiology and biochemistry of midday depression of photosynthesis[J]. Plant Physiology Journal, 1990(6):5. (in Chinese)
(责任编辑:郭雪芳)
Impact of water stress on leaf water potential, photosynthetic parameters and water use efficiency ofPotentillafruticosa
Liu Ying, He Kangning, Xu Te, Wang hui, Liu Yujuan
(Beijing Forestry University, Key Lab. of Soil & Water Conservation and Desertification Combating, Ministry of Education, 100083, Beijing, China)
Abstract:[Background] The rapid development of urbanization have put forward higher requirements on the country road greening and landscape. But the high latitudes, low temperature and drought problem restrain the growth and breeding of many kinds of flowers in the cold and arid Qinghai Province. So a number of scholars began to study the advantages of the local wild species, trying to pick out plant species domesticated from local flowers for landscaping and road greening. [Methods] In order to solve the problem of the cultivation and domestication of greening plants in the cold and arid Qaidam regions, Qinghai Province, explore the photosynthetic physiological characteristics of Potentilla fruticosa and its quantitative relations with soil moistures, provide a scientific basis to the scientific irrigation management of P. fruticosa, and improve the water use efficiency of alpine and arid regions in addition, 17 P. fruticosa seedlings (2 years old) were taken as the research object under a pot experiment to study its rules of response to drought stress and rewatering. Leaf water potential, photosynthetic parameters and photosynthetic diurnal variation process were investigated. Quantitative relations between net photosynthetic rate, transpiration rate, water use efficiency and soil water content were explored to determine the suitable soil moisture thresholds of photosynthesis and water use efficiency of P. fruticosa. [Results & Conclusions] The main results are as follows: 1) With the lowering of soil moisture to 5.97%, more than half of the leaves dried up, and the wilting moisture content of P. fruticosa was about 4.02%. 2)The effects of soil moisture content on leaf water potential and photosynthetic parameters had threshold, too high and too low soil moisture content would inhibit the physiological activity of P. fruticosa. The morning leaf water potential, net photosynthetic rate, and transpiration rate of P. fruticosa had obvious threshold response to the changes of soil moisture. The morning leaf water potential would decrease if the soil moisture was lower than 19.48%, and when the soil moisture varied from 17.03% to 14.37%, the morning leaf water potential would come down at the maximum speed and amplitude, indicating that this range of soil moisture content would significantly improve the water absorption and drought resistance of P. fruticosa. When the net photosynthetic rate reached its maximum the soil moisture content was 20.83%, the soil moisture content of highest water use efficiency was 13.82%, and the hydration compensation point was 4.38%. 3)Under the condition of sandy loam, the net photosynthetic rate could keep above 70% of its maximum unless the soil moisture went down to 12.71%. When the soil moisture content was about 8.33% the water use efficiency could keep 70% of its maximum, and the change speed of water use efficiency would slow down. So soil water content varing from 8.33% to 12.71% could not only satisfy the basic need for the growth of P. fruticosa, but also improve the efficiency of water use to the largest extent. 4) During the growing season of P. fruticosa in Qinghai Province (July) without any rain, the irrigation should be conducted once every two weeks for good growth. P. fruticosa is able to recover from 2-3 weeks’ drought after irrigation, but will die of the drought stress which lasts over one month.
Keywords:water stress; leaf water potential; photosynthesis; water use efficiency; soil moisture content
收稿日期:2015-03-20修回日期: 2015-10-17
第一作者简介:刘颖(1988—),女,硕士研究生。主要研究方向:水土保持和林业生态工程。E-mail: liuyingxiaoxi@163.com †通信 贺康宁(1962—),男,教授,博士生导师。主要研究方向:水土保持和林业生态工程。E-mail:hkn@bjfu.edu.cn
中图分类号:S714
文献标志码:A
文章编号:1672-3007(2016)01-0106-08
DOI:10.16843/j.sswc.2016.01.013
项目名称: 2014年青海省重大科技专项“柴达木盆地盐碱地造林技术研究与示范”(2014-NK-A4-4)
猜你喜欢
学苑创造·A版(2018年5期)2018-05-28
江苏农业科学(2016年10期)2017-02-05
江苏农业科学(2016年4期)2016-06-14
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
江苏农业科学(2015年10期)2015-12-23
少儿科学周刊·儿童版(2015年10期)2015-11-07
长江蔬菜·学术版(2015年1期)2015-07-10
中学生物学(2008年2期)2008-07-07
