
基于ITS序列分析的部分榕属植物系统树构建
2016-09-15 15:46李升星陈家林张太奎刘小珍张汉尧
福建林业科技 2016年1期
李升星,陈家林,张太奎,贺 笑,刘小珍,张汉尧
(1.西南林业大学云南省高校林木遗传改良与繁育重点实验室,云南 昆明 650224;2.西南林业大学国家林业局西南地区生物多样性保育重点实验室,云南 昆明 650224)
基于ITS序列分析的部分榕属植物系统树构建
李升星1,陈家林2,张太奎2,贺 笑2,刘小珍1,张汉尧1
(1.西南林业大学云南省高校林木遗传改良与繁育重点实验室,云南 昆明 650224;2.西南林业大学国家林业局西南地区生物多样性保育重点实验室,云南 昆明 650224)
以榕属中黄金榕、橡皮树、小叶榕3个树种为研究对象,以其叶片为试材提取基因组DNA,进行ITS-PCR扩增。对扩增产物测序结果进行分析,在NCBI上选取同源率较高的27个树种的ITS序列与其进行同源比较,并建立系统树分析它们之间的亲缘进化关系。结果表明:30种榕属植物基于ITS序列分析与传统分类学略有差异,通过结合ITS鉴定与形态分类相结合可以为以后榕属的分类提供借鉴。
榕属;ITS序列分析;系统树构建
榕属(FicusLinn.)隶属桑科,常绿、稀落叶,乔木或灌木,有时匍匐或攀援状,在生态圈中,是包含乔木、灌木和藤本的形态变化较大的一属。本属约1000种,主要分布于热带、亚热带地区。我国约98种3亚种43变种2变型,广泛分布于西南部至东部。榕属植物为低维护性景观植物,韧皮纤维可作麻类代用品,其中有些种类的榕果可药用或食用,在诸多医学著作中均有记载,经济价值较高。榕属植物在文化方面或是实用性上有相当关联,根据相关资料显示榕属植物并没有很明确的化石留存,然而现存的主要树种的辐射适应已经发生在最近的20~40万a。根据榕属植物的生活习性、繁殖系统、花序果的着生位置将其分为4个亚属,制定了早期的榕属分类系统。由于该属植物具有形态特征变异幅度大,种间界限难以界定的特点导致该属植物分类比较困难,如无花果为隐头花序,难以观察其花器官,目前国内关于该属植物系统进化方面的报道尚少[1-4]。
内转录间隔区ITS(Internal Transcribed Spacer)由保守的18S、5.8S 和28S 基因编码区以及非编码的各种间隔区组成。ITS 片段在绝大多数真核生物中表现出了极为广泛的序列多态性,因此在ITS 序列上即使是亲缘关系非常接近的2 个种也能表现出较为明显的差异,显示出最近的进化特征。由于ITS区具有较丰富的信息位点和变异位点,目前已有文献证实ITS是研究许多被子植物类群系统与进化的重要分子标记,例如ITS序列分析对竹种进行系统分类的研究及对各种中药材植物的鉴定等[5-6]。由于ITS能较好地反映出科内、属间、种间的亲缘关系,所以被广泛应用于植物的系统演化及亲缘关系的研究[7-17]。
本研究以云南省昆明市内3种不同榕属树种为试材,通过提取DNA,选取ITS-PCR产物直接测序方法获得其序列,从NCBI上下载同源率较高的序列,并利用MEGA 5.05软件构建系统发育树,对榕属植物进行分析,从分子水平上对榕属植物进行系统分类,以期对该属植物的系统分化和系统分类提供依据,为种质资源的利用和对其遗传改良工作提供参考。
1 试验材料与方法
1.1 榕属植物材料
以昆明市金色俊园小区的3种榕属植物黄金榕(F.microcarpacv.GoldenLeaves)、橡皮树(F.elastica)、小叶榕(F.concinna)新鲜叶片为试验材料。
1.2 叶片DNA提取
将采集的叶片在硅胶中干燥7 d后,用振荡机振荡成粉末状,用BioTeKe 新型快速植物基因组DNA提取试剂盒提取DNA,并对提取的DNA进行琼脂糖凝胶电泳检测。
1.3 ITS序列扩增及测序
根据GenBank中榕属的ITS基因序列选取ITS2和4组合特异性引物扩增样品ITS基因,上游引物ITS2的序列为:5’—AGAAGTCGTAACAAGGTTTCCGTAGG—3’,下游引物ITS4的序列为:5’—TCCTCCGCTTATTGATATGC—3’。以10个树种的DNA为模板,采用25 μL反应体系即1 μL DNA模板、1 μL ITS2、1 μL ITS4、12.5 μL 2×Taq PCR MasterMix和9.5 μL dd H2O,进行PCR扩增;反应程序为:94 ℃预变性3 min;94 ℃变性30 s,50.8 ℃退火30 s,72 ℃延伸90 s,循环30次;72 ℃终延伸5 min;产物于4 ℃保存。取5 μL PCR产物和1 μL上样缓冲液混匀后于1.2%琼脂糖凝胶进行电泳检测,用凝胶成像仪拍照[7]。剩余的PCR产物(约)20 μL委托昆明硕阳科技有限公司,采用双向测序方法进行测序。
1.4 ITS序列分析及形态标记聚类分析
将测序得到的榕属3个树种的ITS序列用ContigExpress软件进行拼接后,提交NCBI公用数据库Genbank,利用NCBI软件的BLAST进行网上对比。对所测得的3个树种和从GenBank得到的树种用系统分析和系统发育分析软件MEGA 5进行系统发育分析。对建树的30种榕树植物的叶形、叶质、果形、果实着生方式通过查阅文献[18]进行记录并作为聚类指标。
2 结果与分析
2.1 DNA提取结果
以榕属10种植物的叶片为材料,使用试剂盒提取的DNA,经1.2%琼脂糖凝胶电泳检测,结果显示所提取的DNA质量较高,条带单一且无拖带现象。
2.2 ITS-PCR扩增结果
以榕属3种植物的基因组DNA为模版,使用引物ITS1与ITS4进行PCR扩增,产物经1.2%琼脂糖凝胶电泳检测,结果(图1)显示使用该引物扩增效果较好,可以得到稳定的条带。根据电泳结果可知,扩增出来的DNA片段的长度约为700 bp。
2.3 ITS序列分析
将扩增获得的榕属3种植物的ITS序列与GenBank的ITS序列进行比对找到与样本同源种的ITS序列27个,其所对应的GenBank序列登记号、长度和G+C含量见表1。供试榕属3种植物扩增获得的ITS序列全长位于757~782 bp之间,通过在GenBank对比找到的27种序列,ITS序列全长均在684~804 bp之间。
每个树种DNA中G+C含量的数值是恒定的,不会因外界环境条件的改变而发生变化,且在同属不同种间,G+C含量的数值也不会有很大差异。由表1可知,供试榕属3种植物的ITS序列中,黄金榕、橡树G+C含量均为62%,小叶榕G+C含量为63%,平均G+C含量为62.3%;在全部53种榕属植物中,整体G+C含量在61%~66%范围内,整体平均G+C含量为62.9%,因此整体含量差异不明显,适于作遗传分析。
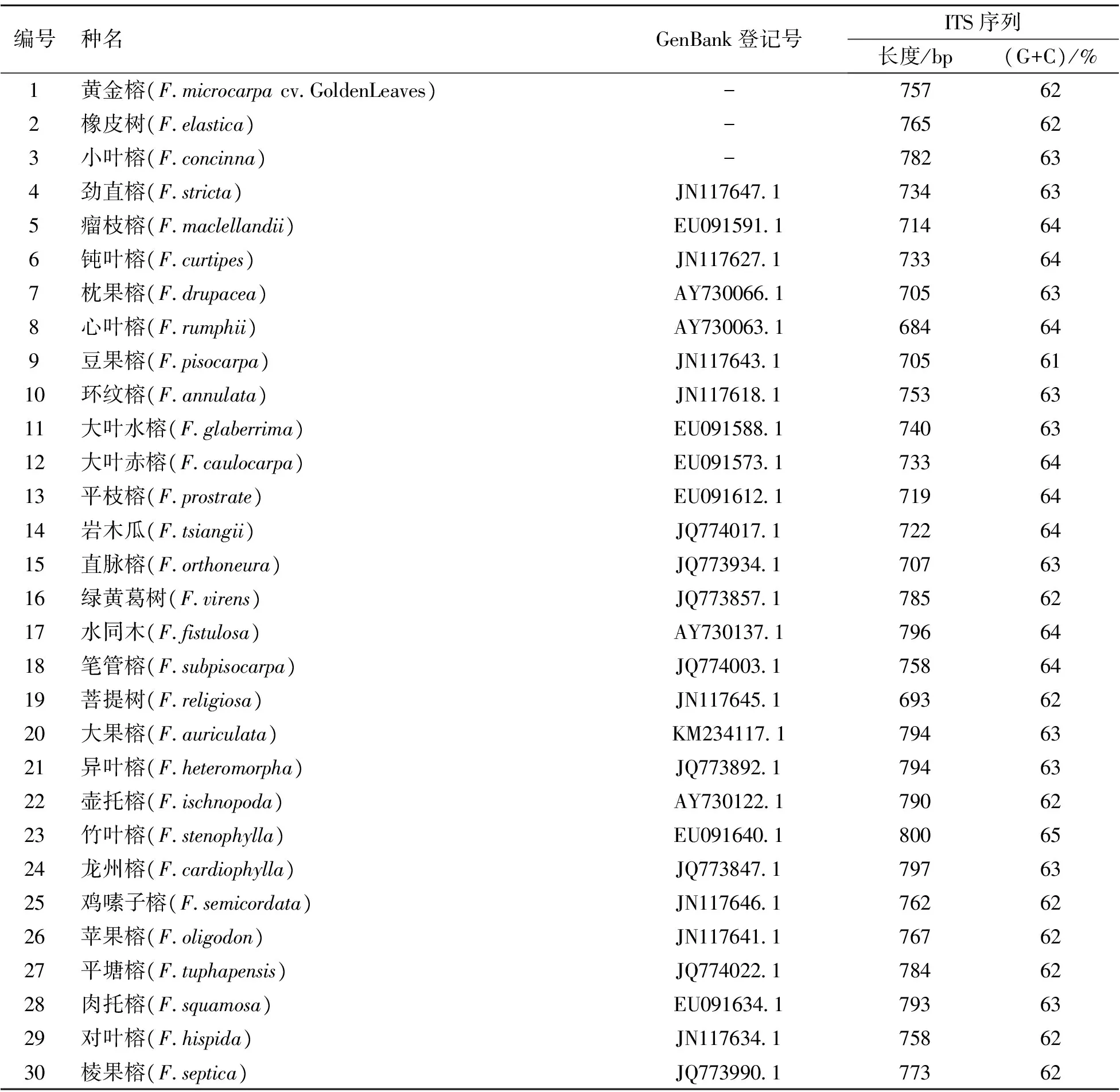
表1 榕属植物的GenBank序列登记号、ITS序列长度、G+C含量
2.4 系统发育与形态标记聚类分析
将供试的榕属10种植物与在GenBank中同源搜索获得的43个近缘种的ITS序列,用MEGA 5.05进行完全比对,再采用邻接距离矩阵法(Neighbor-joining)构建系统发育树并进行形态标记聚类分析(图2)。选用Kimura2-parameter 参数模型,计算所得的遗传距离(由于图表过大,在此只统计遗传距离数据)。
由榕属30种植物的系统树及形态标记热图可以看出:榕属的30个种以100%的支持率被分为2大支。其中供试的3种榕属植物全部聚在一支上,而聚类分析色块图也与所建的系统发育树大部分相吻合。该结果与传统分类基本相符,但并不完全一致。如:小叶榕与岩木瓜、绿黄葛树、笔管榕、菩提树、龙州榕同为榕亚属下榕组,但从图中可以看出:小叶榕与其他几种不聚在一支上,系统发育树和聚类图表明小叶榕与这几种榕属植物关系较远。而聚在一大支上的岩木瓜、绿黄葛树、笔管榕、菩提树、龙州榕理应聚在一小支上,但是岩木瓜单独与无花果亚属的竹叶榕、壶托榕、异叶榕及平塘榕聚为一小支上,从系统发育树上看岩木瓜应归为无花果亚属。
榕属中30个种的遗传距离在0.01~0.08之间,绝大多数的遗传距离<0.6,表明30个榕属植物的亲缘关系较近。其中小叶榕与黄金榕、劲直榕的遗传距离分别是0.005、0.003,表明小叶榕与这2种榕树的亲缘关系非常近。
3 结论与讨论
传统分类学上主要以花果为特征进行分类,榕属植物现有的分类系统亦然,但由于榕属植物为隐头花序,雌雄同株或异株,繁殖器官材料难观察,在分类中困难重重,其中存在争议问题颇多。榕属植物复杂多样的变异通过传统的形态分类学很难揭示榕属性状的演化特点,单就叶的形状而言,就有椭圆形、长圆形、卵形、心形等,如果过分的依赖某一个或几个形态标记进行分类,可能会出现误差,准确性和可靠性不高。本文通过ITS序列分析的方法可以为榕属的分类提供很好的借鉴。从构建的系统树与形态标记热图联合来看,虽然与传统分类大体一致但也出现了一些不相符之处,如小叶榕属于榕属榕组,但是它与榕属环纹榕组的黄金榕、劲直榕亲缘关系非常近。原因可能是传统分类学上过分突出某一个形态特征。
植物学家们一直在努力寻找从多学科手段上可用于系统分类的证据。近年来,ITS序列分析已广泛应用于植物科、亚科、族、属和种间系统学研究[19—22],但在不同的分类等级上,ITS序列所具有的价值是不一样的。对大多数被子植物来说,ITS序列具有十分明显的同步进化,其众多拷贝已变得高度相似或一致化,因此在一定程度上克服了非同源的障碍,另外在许多类群内ITS序列的替代速率是基本稳定的[23]。这样通过ITS构建的系统分支树就能够反映植物的系统发育关系。陈纪云[24]针对榕属植物63种,228个样本,基于5个叶绿体基因片段和1个核基因片段(ITS),计算了遗传距离并构建了系统发育树来检验所选取基因片段对树种鉴定的能力,王双[25]利用MP、ML和BI 3种方法构建系统发育树。根据田婕[26]2006年从ITS序列矩阵的 MP和Bayesian分析结果来看,ITS在榕属系统发育学中的适用性基本得到了肯定。Weiblen[27]2000年就使用ITS作为研究桑科榕属植物的分子标记应用到榕属植物的系统学研究中,对Corner的系统提出了质疑,并证明对于榕属这样一个广泛分布于世界各地的大类群,是比较合适使用ITS 作为研究其整个类群的系统发育关系的分子标记。
本研究表明,供试的3个榕属树种(黄金榕、橡皮树、小叶榕)及同属27个树种ITS序列分析与传统分类学略有差异,通过这种差异可以更好地对榕属系统分类提供一种可行的分类方法借鉴。笔者认为在对榕属植物进行系统发育研究时,可以结合传统分类和ITS提供的信息对榕属植物系统进化做更为精确划分,能使我们对榕属系统进化的认识又向前跨了一步,为榕属植物的分类和系统发育研究提供参考。
[1]张秀实,吴征镒,曹子余.中国植物志[M].北京:科学出版社,1998:23.
[2]尧金燕,赵南先.中国榕属榕亚属植物的系统发育初探[J].广西植物,2009,29(1):7-12.
[3]杨大荣,彭艳琼,张光明.西双版纳热带雨林榕树种群变化与环境的关系[J].环境科学,2002,23(5):29-35.
[4]钱钧碧.福建园林常用榕属乔木栽培管理与大树移苗技术[J].福建农业科技,2009(2):56-58.
[5]张汉尧,刘小珍,龚秀会,等.竹子DNA提取方法改良及ITS-RFLP分析的初步研究[J].江西林业科技,2006(3):3-5.
[6]许亮,谷丽艳,赵丹玉,等.基于ITS序列的植物类中药材鉴定研究及展望[J].中华中医药学刊,2010(4):737-738.
[7]刘峥,张汉尧.旋花科植物ITS序列分析[J].西部林业科学,2012(4):70-74.
[8]刘明航,朱芳明,刘峥,等.柳属植物ITS序列分析[J].贵州林业科技,2012,40(4):30-35.
[9]张道远,陈之端,孙海英,等.用核糖体DNA的ITS序列探讨中国柽柳科植物系统分类中的几个问题[J].西北植物学报,2000,20(3):421-431.
[10]张露,蔡友铭,诸葛强,等.新疆石竹属野生种核糖体DNA的ITS序列与亲缘关系[J].遗传学报,2002,29(6):549-554.
[11]Hsiaoc,Chatterton N J,Asaykh.Phylogenetie relationships of 10 grass species:an assessment of phylogenetic utility of the internal transcribed spacer region in nuclear ribosomal DNA in monocots[J].Genome,1994(37):112-120.
[12]Alvarez I,Wendel J F.Ribosomal ITS sequences and plant phylogenetic inference[J].Molecular Phylogenetics and Evolution,2003,29(3):417-434.
[13]黄海,王安仙,刘杨,等.贵州地区蜘蛛抱蛋属植物ITS1区序列分析及其亲缘关系研究[J].贵州科学,2010,28(2):50-52.
[14]康云,张明理,陈之端,等.基于ITS序列对中国簇毛黄耆亚属(黄耆属)系统学问题的初步研究[J].植物学报,2003,45(2):140-145.
[15]史全良,诸葛强,黄敏仁,等.用ITS序列研究杨属各组之间的系统发育关系[J].植物学报,2001,43(3):323-325.
[16]赵一之,成文连,尹俊,等.用rDNA的ITS序列探讨绵刺属的系统位置[J].植物研究,2003,23(4):402-406.
[17]朱惠芬,杨俊波,张长芹,等.应用ITS序列分析探讨偏花报春的系统位置[J].植物分类学报,2002,40(2):133-138.
[18]刘凤红,刘国俊,韩芳桥,等.结合科研需求自建专业特色电子资源[C]//中国图书馆学会专业图书馆分会2011年学术年会,2011.
[19]Thompson J D,Gbson T J,Plewniak F,et al.The clustal—X windows interface:flexible strategies for mμL tiple sequence alignment aided by quality analysis tools[J].Nucleic Acids Research,1997(25):4876-4882.
[20]Yuan Y M,Kupfer P,Doyle J J.Infrageneric phyIogeny of the genus Gentiana inferrred from nucleotide sequences of the internal transcribed spacers of the nuclear ribosomal DNA[J].American Journal of Botany,1996(83):641-652.
[21]汪小全,邓峥嵘,洪德元,等.铁破锣属的系统位置——ITS(nrDNA)序列证据[J].植物分类学报,1998,36(5):403.
[22]宋葆华,陈之端,汪小全,等.中国苋属nrDNA的ITS序列分析及其系统学意义[J].植物学报,2000,42(11):1184.
[23]张文驹,瞿礼嘉,高巍,等.用ITS序列确定小麦B基因组的可能供体间的关系[J].植物分类学报,1999,37(5):417.
[24]陈纪云.中国榕属植物六个候选DNA条形码的评价[D].上海:华东师范大学,2012.
[25]王双.国产无花果组(Ficussect.Ficus,Moraceae) 的分子系统学研究[D].上海:华东师范大学,2012.
[26]田婕.桑科榕属植物的系统发育学及其在果实性状进化系统学检验中的应用[D].西双版纳:中国科学院研究生院(西双版纳热带植物园),2006.
[27]Weiblen G D.Phylogenetic relationships of functionally dioeciousFicus(Moraceae) based on ribosomal DNA sequences and morphology[J].American Journal of Botany,2000,87(9):1342-1357.
Construction of Phylogenetic Tree Based on ITS Sequence Analysis of someFicusSpecies
LI Sheng-xing1,CHEN Jia-lin2,ZHANG Tai-kui2,HE Xiao2,LIU Xiao-zhen1,ZHANG Han-yao1
(1.KeyLaboratoryforForestGeneticandTreeImprovement&PropagationinUniversitiesofYunnanProvince,SouthwestForestryUniversity,Kunming650224,Yunnan,China;2.KeyLaboratoryofBiodiversityConservationinSouthwestChina,StateForestAdministration,SouthwestForestryUniversity,Kunming650224,Yunnan,China)
Using the leaves ofFicusmicrocarpa,F.elastic,F.concinna,F for ITS-PCR amplification,and then sequencing analysis was carried on.The ITS sequences (27 ITS sequences which had high scores with homology comparison were downloaded from NCBI) was used to build a phylogenetic tree to analyze their genetic evolutionary relationships.Resμlts showed that:It is different between based on ITS sequence analysis and traditional taxonomy,By combining the ITS identification and morphological classification can provide a reference for future classification ofFicus.
Ficus;ITS sequence analysis;Phylogenetic tree
2015-06-02;
2015-09-01
国家林业局948项目(2012-4-62);国家自然科学基金项目(31360404)
李升星(1989—),女,江西宜春人,西南林业大学云南省高校林木遗传改良与繁育重点实验室在读硕士研究生,从事林业生物技术研究。E-mail:shengxing_555@126.com。
张汉尧(1975—),男,福建永定人,西南林业大学云南省高校林木遗传改良与繁育重点实验室教授,博士,硕士生导师,从事植物和微生物分子遗传研究。E-mail:hanyaoz@163.com。
10.13428/j.cnki.fjlk.2016.01.002
Q949
A
1002-7351(2016)01-0009-05
猜你喜欢
现代仪器与医疗(2022年3期)2022-08-12
科学与财富(2021年36期)2021-05-10
民用飞机设计与研究(2020年4期)2021-01-21
少儿科学周刊·儿童版(2017年5期)2017-06-29
少儿科学周刊·儿童版(2017年5期)2017-06-29
红领巾·萌芽(2017年5期)2017-06-23
爆笑show(2016年7期)2017-02-09
山东工业技术(2016年15期)2016-12-01
少儿科学周刊·儿童版(2015年10期)2015-11-07
少儿科学周刊·儿童版(2015年1期)2015-07-07
