
遮光对松属3个树种幼树光合特性和荧光参数的影响
2016-09-26 09:01刘秀梅王华田孟宪鹏曹桂萍黄昌豹山东农业大学山东省高校森林培育重点实验室山东泰安271018国家林业局泰山森林生态系统定位研究站山东泰安271018泰山风景名胜区管理委员会山东泰安271000
浙江农林大学学报 2016年4期
王 巧,聂 鑫,刘秀梅,王华田,孟宪鹏,曹桂萍,李 健,黄昌豹(1.山东农业大学 山东省高校森林培育重点实验室,山东 泰安 271018;2.国家林业局 泰山森林生态系统定位研究站,山东 泰安 271018;.泰山风景名胜区管理委员会,山东 泰安271000)
遮光对松属3个树种幼树光合特性和荧光参数的影响
王巧1,2,聂鑫1,2,刘秀梅1,2,王华田1,2,孟宪鹏3,曹桂萍3,李健3,黄昌豹3
(1.山东农业大学山东省高校森林培育重点实验室,山东 泰安 271018;2.国家林业局 泰山森林生态系统定位研究站,山东 泰安 271018;3.泰山风景名胜区管理委员会,山东 泰安271000)
通过搭建遮光棚模拟空旷地(不遮光对照)、林隙(46%遮光)、林下(81%遮光)等3种光照环境,研究了黑松Pinus thunbergii,油松Pinus tabuliformis,赤松Pinus densiflora等幼树叶片光合特性和叶绿素荧光动力学特征随梯度光照强度的变化规律,以期了解3个树种对不同光环境的适应性对策,为华北地区人工林的天然更新及长期经营提供理论依据。结果表明:随着光照强度的降低,黑松幼树净光合速率(Pn),蒸腾速率(Tr),水分利用效率(EWUE),性能指数(Iabs),质体醌库的面积(Sm),PSⅡ捕获的能量从QA传递到QB的效率(ΨO),PSⅡ捕获的能量从QB传递到PSⅠ的效率(ΨRE),单位反应中心吸收(ABS/RC)、捕获(TRo/RC)、用于电子传递(ETo/RC)的能量均减小。油松幼树净光合速率(Pn),水分利用效率(EWUE),性能指数(Iabs),PSⅡ捕获的能量从QA传递到QB的效率(ΨO),PSⅡ捕获的能量从QB传递到PSⅠ的效率(ΨRE),单位反应中心吸收(ABS/RC)、捕获(TRo/RC)、用于电子传递(ETo/RC)的能量在46%遮光条件下达到最大值。赤松幼树光合特征参数及叶绿素荧光参数在不同遮光条件下差异不显著(P>0.05)。因此,赤松对弱光的利用能力最强,能够适应不同的光照环境;黑松在光照充足的环境中生长状况较好;油松适宜在轻度遮光的林隙环境中生长。图2表2参28
植物生理学;遮光;松属;光合特性;叶绿素荧光
进入21世纪以来,随着中国经济社会的发展,对森林的功能需求逐步由传统的木材生产转为生态效益、经济效益和社会效益的多功能需求。实行森林分类经营以后,对纳入生态公益林的现有人工林如何经营,一直是中国林业主管部门、科技工作者和森林经营技术人员思考和关注的重大问题[1-2]。人工林近自然经营理论的提出,为中国人工营造的生态公益林实现长期经营提供了有力的理论支撑[3-5]。通过合理抚育森林以促进森林天然更新,或通过人工林下播种或植苗方式实现林下更新,逐步将现有单层同龄纯林转化为异龄复层混交林,以达到人工林天然化经营的目的,从而实现对森林系统的长期经营。林下幼树更新是人工林近自然化过程中的关键。幼树阶段是个体生命周期中对环境影响最为敏感和最为脆弱的时期[6-7]。光作为影响植物生长的重要生态因子[8],因冠层结构和郁闭度的差异,导致森林群落下层出现大小不等的光斑和光强度,甚至光谱成分亦有差异,并且随时间呈现出动态变化。光辐射量的变化制约着幼树的更新、存活和生长[9-10],在树木生理特征上引起直接而迅速的响应。林下幼树的暗呼吸速率、光补偿点和光饱和点均随着光强的降低而降低[11-13]。光合原初反应和叶绿素荧光存在着密切关系,叶绿素荧光技术可以快速检测植株在胁迫下光合作用的真实行为,评价光合机构的功能和环境胁迫对它的影响。慢相荧光动力学参数主要揭示了光合暗反应启动后的光能利用和分配情况[14],而快相荧光动力学JIP测定技术可以分析光合作用的光反应过程和能量流程,并因其方便、快捷、无损、信息丰富等优点开始应用于植物光合功能的研究当中[15-16]。植物在弱光环境中叶绿素的慢相荧光动力学参数PSⅡ光化学效率、非光化学猝灭系数增加,光化学猝灭系数和电子传递速率逐渐降低[11,17-19]。针对松属Pinus树种光合作用和叶绿素荧光动力学特征对梯度光强的响应尚缺乏必要研究,快相荧光动力学技术在光逆境下的研究还未见报道。本研究以华北地区山地森林生态系统中的主要针叶树种油松Pinus tabuliformis,黑松Pinus thunbergii,赤松Pinus densiflora等3个树种为研究对象,采用人工遮光的方法模拟旷地、林隙、林下3种不同的光照强度,对不同光强下3个树种幼树的光合生理特性和快相叶绿素荧光动力学特性进行了对比研究,探讨它们在不同光辐射下的光保护机制和光合机构的光反应过程,研究3个松属树种的天然更新能力与近自然森林经营的关系,揭示3个树种对梯度光环境的适应对策,从而为华北地区山地人工林的抚育和长期经营提供理论依据。
1 材料与方法
1.1试验地概况
试验在山东农业大学南校区林学实验站(36°16′N,117°11′E)进行。该地区属于温带季风大陆性气候,四季分明,年均温为12.8℃,极端高温40.0℃,极端低温-20.0℃,≥10℃积温4 283.1℃,无霜期186.6 d,年降水量600~700 mm,降水分布不均匀,相对湿度65%。
1.2试验材料与试验设计
试验材料为生长健壮的2年生实生油松、黑松、赤松营养袋苗,于2013年11月栽植于山东农业大学南校区林学试验站,2014年5月用黑色遮光网搭建遮光棚对供试材料进行遮光处理。遮光棚为东西向,高为2.0 m,南北开敞以便于通风透气。设置不遮光、1层遮光网覆盖、2层遮光网覆盖,形成不遮光对照,46%,81%不同遮光强度3种处理,分别模拟空旷地、林隙和林冠下生境光照条件。栽植苗木12株·小区-1,株距为30 cm,行距为50 cm,重复3次。遮光处理3个月后,于8月中旬进行光合和快相叶绿素荧光参数的测定。
1.3测定方法
1.3.1光合作用参数的测定在不遮光处理的油松、黑松、赤松幼树中,各选择3株·小区-1长势健壮均一的幼树,在当年新梢中部选择5束针叶,用配备针叶叶室[PP SYSTEMS PLC(C)]的便携式光合测定系统(CIRAS-2)对光响应曲线进行测定。控制大气二氧化碳摩尔分数400 μmol·mol-1,叶室温度24~26℃,相对湿度 40%±5%,发光二极管(LED)光源控制光合有效辐射强度梯度为 1 400,1 200,1 000,800,700,600,500,400,300,200,100,0 μmol·m-2·s-1。每个光照强度下设定数据采集时间为2 min,于晴天9:00-11:30进行测定。同时在不遮光处理、46%遮光处理和81%遮光处理的油松、黑松、赤松小区内各选择3株长势良好健壮均一的幼树,测定针叶叶片的净光合速率(Pn),蒸腾速率(Tr),胞间二氧化碳摩尔分数(Ci)和水分利用效率(EWUE),重复测定3次·株-1,LED光源控制光合有效辐射强度为1 200 μmol·m-2·s-1。用于光合作用参数测定的针叶采集后,放于自封袋,置于冰盒中带回,用EPSON PERFECTION V700 PHOTO根系扫描仪扫描针叶特征影像,应用根系分析软件WINzip获取针叶面积数据。净光合速率-光响应曲线在低光强(0~200 μmol·m-2·s-1)下呈直线部分的斜率即表观量子效率(EAQY),将测得的净光合速率(y)与相应的光量子(x)做直线回归y=a+bx,其中a为暗呼吸速率,b为表观光量子效率,a/b为光补偿点,即该直线与X轴的交点为其光补偿点(PLCP),与Y轴的交点则为光下的暗呼吸速率(Rd),光饱和点(PLSP)由抛物线模型y=ax+bx+c,以Pnmax时的光合有效辐射(RPAR)值计算。水分利用效率(EWUE)的计算公式为:EWUE=Pn/Tr。
1.3.2快相叶绿素荧光参数测定采用Handy PEA(Plant Efficiency Analyser;Hansatech Instrument Ltd.,英国)测定3种松树在不同遮光处理下的快相叶绿素荧光参数。将针叶平铺在荧光夹中,覆盖住4 mm2的测试孔。暗适应20 min后,由650 nm的红光诱导,在3 000 μmol·m-2·s-1的饱和光强下,测定1 s,选取3株·小区-1长势一致的健壮幼树,重复测定3次·株-1。用PEA Plus V1.10专业软件对快相叶绿素荧光参数性能指数(Iabs),质体醌库的大小(Sm),PSⅡ捕获的能量从QA传递到QB的效率(ΨO),PSⅡ捕获的能量从QB传递到PSⅠ的效率(ΨRE),单位反应中心吸收的能量(ABS/RC),单位反应中心捕获的能量(TRo/RC),单位反应中心用于电子传递的能量(ETo/RC)进行整理。
2 结果与分析
2.1全光照条件下油松、黑松、赤松的光响应
分析图1和表1可知:在全光照条件下,油松、黑松、赤松等3个树种的净光合速率均随着光合有效辐射量的增加呈现出大幅度增加的规律,到达一定程度后趋于平稳,达到光饱和点之后,随着光强的增加,净光合速率开始下降。最大净光合速率(Pnmax)反映了植物的生长速度。赤松最大净光合速率(Pnmax)最大,黑松次之,油松最小;暗呼吸速率(Rd)反映出在没有光照条件下的呼吸速率,暗呼吸速率呈现出油松>黑松>赤松的规律;光饱和点(PLSP)和光补偿点(PLSP)反映了植物利用强光和弱光能力的大小,黑松光饱和点最高,赤松光补偿点最低;表观量子效率(EAQY)反映了植物的耐荫性以及在弱光下吸收、转化和利用光能的能力,表观量子效率呈现出赤松>油松>黑松的规律。在3种松树中,赤松最大净光合速率最大,暗呼吸速率最小,光补偿点最低,表观量子效率最高,表明赤松对弱光的利用能力最强。
2.2遮光对油松、黑松、赤松光合参数的影响
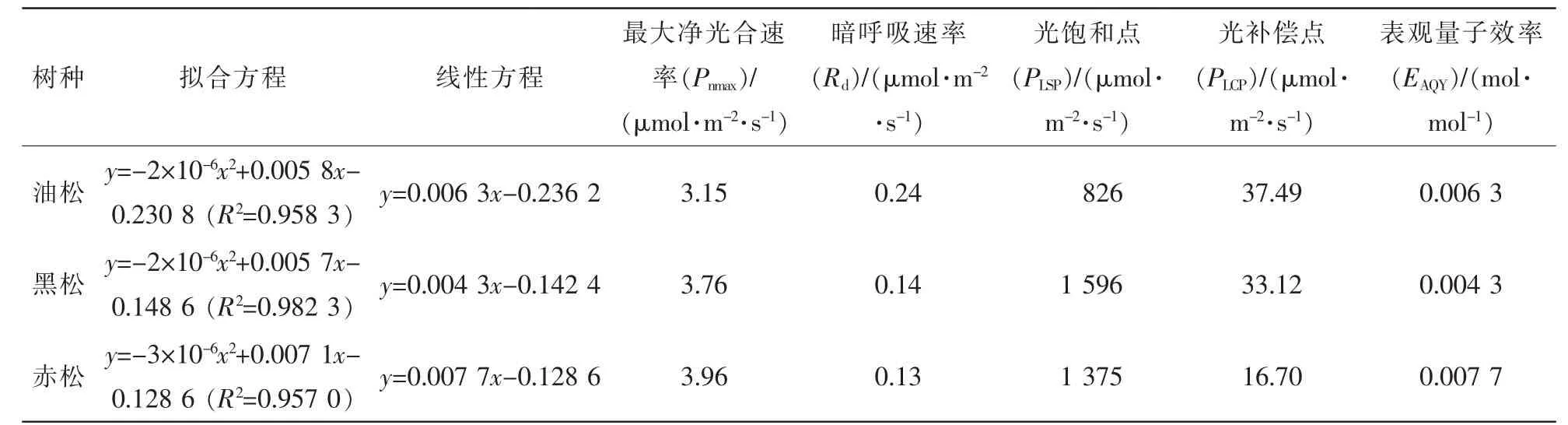
表1 3个松属树种全光照条件下光合参数Table 1 Photosynthetic parameters of 3 pine species under 100%light regimes

图1 松属3个树种全光照条件下光响应曲线Figure 1 Photosynthetic light response curves of3 pine species under 100%light regimes

图2 不同遮光条件对3个松属树种光合参数的影响Figure 2 Photosynthetic parameters of 3 pine species under different light regimes
分析图2可知:光照条件的差异对黑松、油松、赤松的光合作用能力产生了不同的影响。3种松树幼树净光合速率(Pn),蒸腾速率(Tr),胞间二氧化碳浓度(Ci)及水分利用效率(EWUE)在不同光照条件下差异显著(P<0.05)。在全光照条件下净光合速率呈现出黑松>赤松>油松的规律,而在遮光条件下,油松>赤松>黑松;赤松蒸腾速率最大,随着遮光度的增加,油松的蒸腾速率大于黑松;全光照条件下水分利用效率呈现出黑松>油松>赤松的规律,46%遮光条件下赤松>油松>黑松,81%遮光条件下黑松和油松水分利用效率大于赤松。遮光度越高,黑松幼树的净光合作用能力越低,蒸腾速率与水分利用效率也随之降低,而胞间二氧化碳摩尔分数在46%遮光条件下>81%遮光>对照,净光合速率、蒸腾速率、胞间二氧化碳摩尔分数及水分利用效率均具有显著差异(P<0.05);油松幼树的净光合速率、蒸腾速率、胞间二氧化碳摩尔分数分别呈现出46%遮光>81%遮光>对照、81%遮光>46%遮光>对照、46%遮光>对照>81%遮光的规律,不同遮光条件下的水分利用效率差异不显著(P>0.05),46%遮光条件下利用率较高;赤松幼树的净光合速率和蒸腾速率在不同遮光条件下差异不明显(P>0.05),水分利用效率在46%遮光条件下利用率最高。从黑松、油松、赤松在不同光照条件下的光合特征参数的变化可以看出,黑松幼树对光强变化的适应能力强,但是不耐阴;油松幼树有一定的耐阴能力,在46%遮光条件下光合作用能力较强;赤松幼树耐阴性强,能够很好地在光合有效辐射量低的条件下生存。
2.3遮光对油松、黑松、赤松快相叶绿素荧光参数的影响
叶绿素荧光可以从一定程度上反映环境因子对植物的影响,快相叶绿素荧光参数能够深入地揭示以PSⅡ为主的植物光合机构对环境的适应机制。遮光条件下PSⅡ反应中心及受体侧的变化及能量流动情况如表2所示。性能指数Iabs反应光合机构的状态,黑松和油松的性能指数优于赤松,在0.05水平上差异显著。不同遮光条件下黑松的性能指数差异不显著(P>0.05),但随着遮光度的增加性能指数降低;油松的性能指数有显著的差异(P<0.05),在46%遮光状况下最大;赤松的性能指数差异不明显(P>0.05)。3种松树在遮光条件下,PSⅡ受体侧电子受体库(PQ)的容量(Sm)均变小,从QA-进入电子传递链的电子减少,黑松和油松变化差异明显,赤松无明显变化。黑松、油松和赤松PSⅡ捕获的能量从QA传递到QB的效率(ΨO)及PSⅡ捕获的能量从QB传递到PSⅠ的效率(ΨRE)差异显著(P<0.05),赤松PSⅡ捕获的能量从QA传递到QB的效率低于油松和黑松,但是PSⅡ捕获的能量从QB传递到PSⅠ的效率高于油松和黑松。遮光程度越大,黑松PSⅡ捕获的能量从QA传递到QB的效率和PSⅡ捕获的能量从QB传递到PSⅠ的效率越小;油松在46%遮光条件下效率最高;赤松的PSⅡ捕获的能量从QA传递到QB的效率和PSⅡ捕获的能量从QB传递到PSⅠ的效率不受遮光条件的影响。
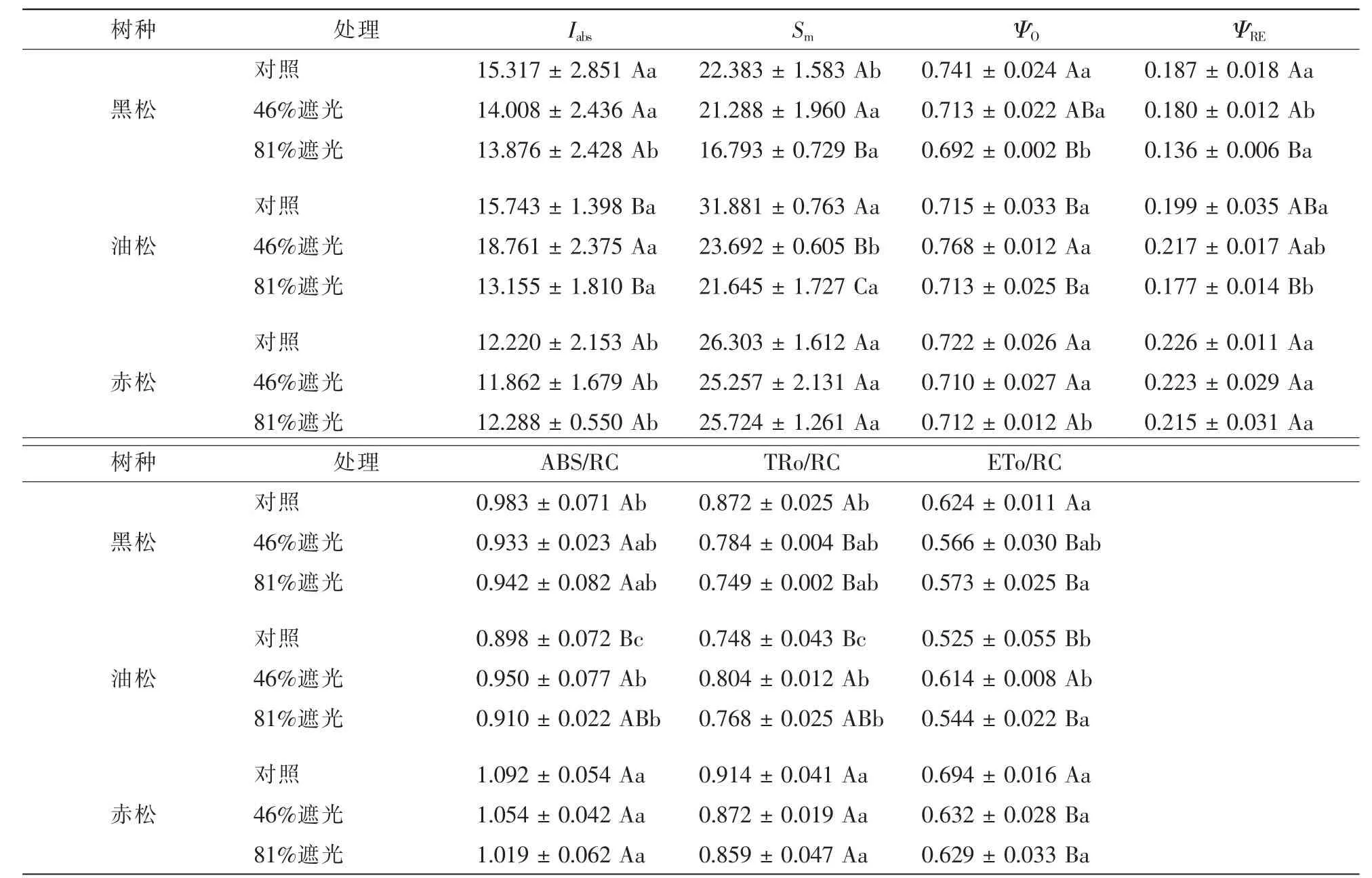
表2 不同遮光条件对3个松属树种快相叶绿素荧光参数的影响Table 2 Effects on snapshot chlorophyll fluorescence parameters of 3 Pine species under different light regimes
3种松树在遮光条件下单位反应中心吸收的能量(ABS/RC),捕获的能量(TRo/RC)以及用于电子传递的能量(ETo/RC)有显著的差异(P<0.05)。赤松单位反应中心吸收、捕获及电子传递的能力均大于黑松和油松,PSⅡ光合机构对光能的利用能力最强。黑松单位反应中心对光能的吸收、捕获及利用能力在全光照条件下最强,随着光合有效辐射量的减小对光能的利用能力降低;油松在46%遮光条件下单位反应中心对光能的利用能力最强;光合有效辐射量的变化对赤松单位反应中心的光能利用效率无显著的影响(P>0.05)。快相叶绿素荧光参数结果表明:油松光合机构的状态以及PSⅡ反应中心捕获的能量在QA与QB间的传递效率优于黑松和赤松,但是赤松单位反应中心吸收及用于电子传递的能量均显著大于黑松和油松,能量利用效率高,致使PSⅡ捕获的能量从QB传递到PSⅠ的效率高。
3 讨论与结论
植物最大净光合速率、暗呼吸速率、光饱和点、光补偿点以及表观量子效率的高低能够直接反映植物对弱光的适应性和捕获光量子用于光合作用的能力。植物的光补偿点越低,越有利于适应弱光环境,在弱光下进行光合作用,并且低光补偿点、高光饱和点的植物对复杂的光环境有更强的适应性。植物光合机构每吸收1 mol光量子进行光合作用后释放出的氧气的摩尔数或者同化的二氧化碳的摩尔数越高,则表观量子效率越高,植物吸收与转换光能的色素蛋白复合体越高,植物耐阴性越强[22-23]。最大净光合速率大则植物的净光合潜能大,生长速度快。暗呼吸速率小则代谢和生理活动消耗的光合产物少。本研究中,赤松幼树具有最低的光补偿点和暗呼吸速率、较高的光饱和点、最高的表观量子效率和最大净光合速率,因此,在黑松、油松和赤松3种树种当中,赤松苗木耐荫性强并且能够适应各种不同的光环境。
光照是影响植物净光合速率、蒸腾速率、胞间二氧化碳摩尔分数及水分利用效率的重要因素,不同植物对光强变化的响应特征不同。经过3个月的遮光处理,不同遮光条件下黑松、油松和赤松对光环境的反映有明显的不同。黑松的净光合速率、蒸腾速率与遮光程度呈现出负相关的关系,遮光强度越大,光合参数值净光合速率、蒸腾速率越小,体现植物在长期适应过程中为达到光合作用和蒸腾作用的最佳状态而形成的适应策略的水分利用效率也减少,弱光条件下气孔限制值降低,胞间二氧化碳摩尔分数升高,有利于二氧化碳的固定和植物对光能的利用率[24-25],然而净光合速率仍然下降,则捕获光合有效辐射量不足是遮光条件下光合速率下降的主要原因。油松的净光合速率、水分利用效率在46%遮光条件下最大,而蒸腾速率、胞间二氧化碳摩尔分数则随遮光程度增大而增大,可能是因为在全光照条件下,光抑制对光合反应中心造成一定的损伤,并且强光下细胞壁加厚影响气体交换速率以及呼吸速率增强[26],导致全光照条件下油松幼树光合能力并未明显高于46%遮光条件下的光合能力。赤松幼树因其光补偿点低,表观量子效率高,对弱光利用能力强,所以赤松的净光合速率、蒸腾速率在不同光环境中无显著差异。徐飞等[25]对麻栎Quercus acutissima,刺槐Robinia pseudoacacia光合特性对不同光照强度的响应结果表明,随遮光程度的增加,麻栎的净光合速率、蒸腾速率、气孔导度逐渐降低,刺槐的净光合速率则在适度遮光下最大。温达志等[27]对黧蒴Castanopsis fissa和九节Psychotria rubra的研究表明:与全光照相比,遮光下植物叶片净光合速率、气孔导度、胞间二氧化碳摩尔分数与大气二氧化碳摩尔分数之比变化不明显。因此,在不同光照条件下生长的植物,光合速率的变化与植物的种类及其所处的环境有关,并且呈现出不同的变化规律。
光环境的变化不仅影响植物的光合与蒸腾,还引发植物光合机构的能量流动和电子传递的变化,从而导致植物光合机构功能异常。天线色素(Chl)吸收的能量(ABS)一部分以热能和荧光(F)的形式耗散掉,另一部分被反应中心(RC)捕获(TR),反应中心的激发能转化成还原能之后将QA还原成QA-,QA-被重新氧化产生电子传递(ET),传递的电子用于固定二氧化碳或其他途径。黑松、油松和赤松幼树在遮光条件下单位反应中心吸收的能量(ABS/RC),捕获的能量(TRo/RC)以及用于电子传递的能量(ETo/RC)的不同,引起3种松属树种在弱光下对光响应机制的差异。赤松幼树在全光照、46%遮光及81%遮光条件下单位反应中心吸收、捕获以及用于电子传递的能量大于黑松和油松,并且差异显著(P<0.05),赤松的电子传递效率(ETR)高,利用弱光的能力强。随着生长光强的减弱,黑松单位反应中心吸收、捕获以及用于电子传递的能量值降低,表明遮光降低了黑松叶片PSⅡ光合电子传递活性,遮光条件下黑松单位反应中心吸收的能量没有明显减少,但是单位反应中心捕获的能量以及用于电子传递的能量值显著降低(P<0.05),因此,热耗散掉的能量比例升高,为碳同化积累的能量减少,这可能是黑松对弱光适应的一种保护机制,与刘建锋等[11]对遮光条件下崖柏Thuja sutchuenensis的叶绿素荧光动态变化情况相同。油松在全光照条件下过剩光能导致PSⅡ光反应中心关闭,而在46%遮光条件下单位反应中心吸收、捕获以及用于电子传递的能量值增大,说明遮光导致PSⅡ光反应中心开放度增加。遮光处理能够导致PSⅡ反应中心活性下降,电子传递过程受到抑制,同化力供应不足,植物的光合碳同化能力被限制。PSⅡ和PSⅠ在电子传递体PQ、细胞色素b6f复合体与质蓝素的协同作用下完成植物光合作用[28],PSⅡ捕获的能量从QA传递到QB的效率(ΨO)及PSⅡ捕获的能量从QB传递到PSⅠ的效率(ΨRE)的变化表明了电子在QA,QB及PSⅠ间的传递状况。黑松幼树在弱光条件下PSⅡ捕获的能量从QA传递到QB的效率及PSⅡ捕获的能量从QB传递到PSⅠ的效率减小,PSⅡ受体侧功能破坏,QB向PSⅠ的电子传递受到抑制,而QB作为光合电子传递过程中含量最多的电子传递体,因它们既传递电子,又传递质子的特异性,成为光合电子传递的关键步骤和限速环节,因而QB的活性被抑制,影响到PSⅠ正常的生理功能,最终致使光合作用能力下降[15-16]。
综上所述,黑松、油松和赤松幼树在弱光下的光响应和叶绿素荧光响应状况不同。赤松幼树光补偿点低、表观量子效率高、净光合速率高、PSⅡ反应中心对电子的传递能力强,对弱光的利用能力强,在旷地、林隙、林下不同光照强度条件下均能较好地生长。黑松幼树在弱光条件下净光合速率和PSⅡ反应中心活性均降低,电子传递过程受抑制,光合作用能力减弱。黑松在林隙及林下照光不充足的环境中难以保持良好的生长状态,因此,黑松幼树适宜在旷地生存。强光抑制油松PSⅡ反应中心的开放度,在轻度遮光条件下光合作用能力最强,油松适宜在林隙环境中生存。在黑松、油松、赤松等3种松属树种人工林近自然化经营过程中,应当分别选取不同的择伐方式,为其提供适宜幼树天然更新的光环境条件,以保持森林最基本的自然结构特征,从而使森林的生态功能最大程度地发挥出来,为国家的社会发展奠定坚实的生态环境基础。
[1]周建云,李荣,何景峰,等.近自然经营对辽东栎林优势乔木更新的影响[J].林业科学,2013,49(8):15-20. ZHOU Jianyun,LI Rong,HE Jingfeng,et al.Regeneration of the dominant arbors after close-to-natural management of Quercus wutaishanica forest[J].Sci Silv Sin,2013,49(8):15-20.
[2]李婷婷,陆元昌,庞丽峰,等.杉木人工林近自然经营的初步效果[J].林业科学,2014,50(5):90-100. LI Tingting,LU Yuanchang,PANG Lifeng,et al.Initial effect of close-to-nature management of Chinese fir plantation [J].Sci Silv Sin,2014,50(5):90-100.
[3]KUIJPER D P J.Lack of natural control mechanisms increases wildlife-forestry conflict in managed temperate European forest systems[J].Euro J For Res,2011,130(6):895-909.
[4]MORADI M,MOHADJER M R M,SEFIDI K,et al.Over-mature beech trees(Fagus orientalis Lipsky)and close-tonature forestry in northern Iran[J].J For Res,2012,23(2):289-294.
[5]KOHSAKA R,HANDOH I C.Perceptions of“close-to-nature forestry”by German and Japanese group:inquiry using visual materials of‘cut'and‘dead'wood[J].J For Res,2006,11(1):11-19.
[6]HAMRICK J L,LEE J M.Effect of soil surface topography and litter cover on the germination survival and growth of musk thistle(Garduus nutans)[J].Am J Bot,1987,74(3):451-457.
[7]刘长成,刘玉国,郭柯.4种不同生活型植物幼苗对喀斯特生境干旱的生理生态适应性[J].植物生态学报,2011,35(10):1070-1082. LIU Changcheng,LIU Yuguo,GUO Ke.Ecophysiological adaptations to drought stress of seedlings of four plant species with different growth forms in karst habitats[J].Chin J Plant Ecol,2011,35(10):1070-1082.
[8]ROZENDAAL D M A,HURTADO V H,POORTER L.Plasticity in leaf traits of 38 tropical tree species in response to light;relationships with light demand and adult stature[J].Funct Ecol,2006,20(2):207-216.
[9]PORTSMUTH A,NIINEMETS Ü.Structural and physiological plasticity in response to light and nutrients in five temperate deciduous woody species of contrasting shade tolerance[J].Funct Ecol,2007,21(1):61-77.
[10]林波,刘庆.4种亚高山针叶林树种的表型可塑性对不同光照强度的响应[J].生态学报,2008,28(10):4665-4675. LIN Bo,LIU Qing.Plasticity responses of 4 tree species in subalpine-coniferous forest to different light regimes[J].Acta Ecol Sin,2008,28(10):4665-4675.
[11]刘建锋,杨文娟,江泽平,等.遮荫对濒危植物崖柏光合作用和叶绿素荧光参数的影响[J].生态学报,2011,31(20):5999-6004. LIU Jianfeng,YANG Wenjuan,JIANG Zeping,et al.Effects of shading on photosynthetic characteristics and chlorophyll fluorescence parameters in leaves of the endangered plant Thuja sutchuenensis[J].Acta Ecol Sin,2011,31 (20):5999-6004.
[12]刘柿良,马明东,潘远智,等.不同光强对2种桤木幼苗光合特性和抗氧化系统的影响[J].植物生态学报,2012,36(10):1062-1074. LIU Shiliang,MA Mingdong,PAN Yuanzhi,et al.Effects of light regimes on photosynthetic characteristics and antioxidant system in seedlings of two alder species[J].Chin J Plant Ecol,2012,36(10):1062-1074.
[13]MACEDO M F,DUARTE P,FERREIRA J G.The influence of incubation periods on photosynthesis-irradiance curves[J].J Exp Mar Bio Ecol,2002,274(2):101-120.
[14]邱念伟,周峰,顾祝军,等.5种松属树种光合功能及叶绿素快相荧光动力学特征比较[J].应用生态学报,2012,23(5):1181-1187. QIU Nianwei,ZHOU Feng,GU Zhujun,et al.Photosynthetic functions and chlorophyll fast fluorescence characteristics of five Pinus species[J].Chin J Appl Ecol,2012,23(5):1181-1187.
[15]STIRBET A,GOVINDJEE.On the relation between the Kautsky effect(chlorophyll a fluorescence induction)and photosystem(Ⅱ)basics and applications of the OJIP fluorescence transient[J].J Photochem Photobiol B Biol,2011,104(1/2):236-257.
[16]李鹏民,高辉远,STRASSER R J.快速叶绿素荧光诱导动力学分析在光合作用研究中的应用[J].植物生理与分子生物学学报,2005,31(6):559-566. LI Pengmin,GAO Huiyuan,STRASSER R J.Application of the fast chlorophyll fluorescence induction dynamics analysis in photosynthesis study[J].J Plant Physiol Molec Biol,2005,31(6):559-566.
[17]王建华,任士福,史宝胜,等.遮荫对连翘光合特性和叶绿素荧光参数的影响[J].生态学报,2011,31(7):1811-1817. WANG Jianhua,REN Shifu,SHI Baosheng,et al.Effects of shades on the photosynthetic characteristics and chlorophyll fluorescence parameters of Forsythia suspensa[J].Acta Ecol Sin,2011,31(7):1811-1817.
[18]吕铖香,张明如,邹伶俐.模拟酸雨与光强处理对芒萁叶绿素及荧光特性的影响[J].浙江农林大学学报,2015,32(1):52-59. LÜ Chengxiang,ZHANG Mingru,ZOU Lingli.Chlorophyll fluorescence characteristics with simulated acid rain and light intensity treatments for Dicranopteris dichotoma[J].J Zhejiang A&F Univ,2015,32(1):52-59.
[19]DENG Yanming,LI Congcong,SHAO Qingsong,et al.Differential responses of double petal and multi petal jasmine to shading(Ⅰ)Photosynthetic characteristics and chloroplast ultrastructure[J].Plant Physiol Biochem,2012,55:93 -102.
[20]COLEY P D.Herbivory and defensive characteristics of tree species in a low land tropical forest[J].Ecol Monogr,1983,53(2):209-233.
[21]伍维模,李志军,罗青红,等.土壤水分胁迫对胡杨、灰叶胡杨光合作用—光响应特性的影响[J].林业科学,2007,43(5):30-35. WU Weimo,LI Zhijun,LUO Qinghong,et al.Effects of soil water stress on light response curves of photosynthesis of Populus euphratica and Populus pruinosa[J].Sci Silv Sin,2007,43(5):30-35.
[22]LEE D W,BONE R A,TARSIS S L,et al.Correlates of leaf optical properties in tropical forest sun and extremeshade plants[J].Am J Bot,1990,77(3):370-380.
[23]RICHARDSON A D,BERLYN G P.Spectral reflectance and photosynthetic properties of Betula papyrifera(Betulaceae)leaves along an elevational gradient on Mt.Mansfield,Vermont,USA[J].Am J Bot,2002,89(1):88-94.
[24]方江保,殷秀敏,余树全,等.光照强度对苦槠幼苗生长与光合作用的影响[J].浙江林学院学报,2010,27 (4):538-544. FANG Jiangbao,YIN Xiumin,YU Shuquan,et al.Growth and photosynthesis of Castanopsis sclerophylla seedlings with three light intensity treatments[J].J Zhejiang For Coll,2010,27(4):538-544.
[25]徐飞,郭卫华,徐伟红,等.不同光环境对麻栎和刺槐幼苗生长和光合特征的影响 [J].生态学报,2010,30 (12):3098-3107. XU Fei,GUO Weihua,XU Weihong,et al.Effects of light intensity on growth and photosynthesis of seedlings of Quercus acutissima and Robinia pseudoacacia[J].Acta Ecol Sin,2010,30(12):3098-3107.
[26]BLACK K,DAVIS P,McGRATH J,et al.Interactive effects of irradiance and water availability on the photosynthetic performance of Picea sitchensis seedlings:implications for seeding establishment under different management practices[J].Ann For Sci,2005,62(5):413-422.
[27]温达志,叶万辉,林植芳,等.全光和遮阴下2种亚热带木本植物的光合作用对光的响应[J].热带亚热带植物学报,2001,9(3):248-255. WEN Dazhi,YE Wanhui,LIN Zhifang,et al.Photosynthetic responses of two subtropical woody plants growing under sun and shade environments[J].J Trop Subtrop Bot,2001,9(3):248-255.
[28]李利,潘响亮,李宏.模拟干旱和盐分胁迫对沙枣幼苗PSⅡ活力的影响[J].西北植物学报,2001,31(4):768-775. LI Li,PAN Xiangliang,LI Hong.Comparative effects of NaCl and polyethylene glycol on PSⅡactivity of Elaeagnus angustifolia[J].Acta Bot Boreal-Occident Sin,2011,31(4):768-775.
Photosynthetic characteristics and chlorophyll fluorescence of three Pinus tree species with shading
WANG Qiao1,2,NIE Xin1,2,LIU Xiumei1,2,WANG Huatian1,2,MENG Xianpeng3,CAO Guiping3,LI Jian3,HUANG Changbao3
(1.Key Laboratory of Silviculture of Shandong Province,Shandong Agriculture University,Tai'an 271018,Shandong,China;2.Taishan Forest Ecosystem Research Station,State Forestry Administration,Tai'an 271018,Shandong,China;3.Taishan Scenic Area Management Committee,Tai'an 271000,Shandong,China)
Seedling stage is the most sensitive and vulnerable period of the individual life cycle on the environment.The update,survival and growth of young trees influenced larger by the change of light radiation.This paper focused the study on the change rule of young tree leaf photosynthetic characteristics and chlorophyll fluorescence kinetics characteristics along with the change of light intensity,which were three species,Pinus thunbergii,Pinus tabuliformi,Pinus densiflora.The goal was to investigate the three species'adaptive strategy to different light environments through building shading to mimic the open place,gap,understory(i.e.0%,46% and 81%sun-shading)and provide theoretical for natural regeneration and long-term management of plantations in north China.Variables including Pn(net photosynthetic rate),Tr(transpiration rate),EWUE(water use efficiency),Iabs(performance index on absorption basis),Sm(normalized total complementary area above the OJ-I-P transit),ΨO(probability that a trapped exciton moves an electron into the electron transport chain beyondQA-(electron transfer mediators)),ΨRE(probability that a trapped exciton moves an electron into the electron transport chain beyond QB-(electron transfer mediators)),ABS/RC(absorption flux per RC(per QAreducing PSⅡreaction center)),TRo/RC(trapped energy flux per RC),ETo/RC(electron transport flux per RC)had been measured.The results showed that:For the photosynthetic parameters,the photosynthetic parameters of three Pinus trees were significantly different(P<0.05)and Pnwas the largest with P.densiflora seedlings.Pn,Trand EWUEof P.thunbergii decreased with the shading degree increasing;Pnof P.tabuliformis reached maximum under 46%shading degree with Trand EWUEreaching maximum under 81%shading degree;Pn,Trand EWUEof P.densiflora showed no obvious differences.For the chlorophyll fluorescence kinetics parameters,the Iabs,Sm,ΨO,ΨRE,ABS/RC,TRo/RC,ETo/RC under different shading degree of the three Pinus trees were significantly different(P<0.05).Iabs,Sm,ΨO,ΨRE,ABS/RC,TRo/RC and ETo/RC of P.thunbergii had the same rule with the photosynthetic parameters;Iabs,ΨO,ΨRE,ABS/RC,TRo/RC and ETo/RC of P.tabuliformis reached maximum under 46%shading degree with Smreaching maximum under 81%shading degree;The chlorophyll fluorescence kinetics parameters of P.densiflora wasn't significantly different when the shading degree changed (P>0.05).In general,P.densiflora had the strongest ability of using weak light and could adapted to various light environments;P.thunbergii grew well with enough light while P.tabuliformis grew well in light shaded gap.[Ch,2 fig.2 tab.28 ref.]
plant physiology;shade;Pinus;photosynthetic characteristics;chlorophyll fluorescence parameters
S718.3;S791.24
A
2095-0756(2016)04-0643-09
10.11833/j.issn.2095-0756.2016.04.013
2015-07-12;
2015-10-16
国家林业公益性行业科研专项(201104002-6);山东省泰安市科技计划项目(2011-40)
王巧,从事风景林与城市林业研究。E-mail:1029379692@qq.com。通信作者:王华田:教授,博士生导师,从事森林培育与森林生态等研究。E-mail:wanght@sdau.edu.cn
猜你喜欢
北华大学学报(自然科学版)(2022年5期)2022-11-21
美食(2022年1期)2022-02-18
新农民(2020年2期)2020-12-08
农业与技术(2020年12期)2020-07-04
河北果树(2020年1期)2020-02-09
美食(2019年7期)2019-09-10
农学学报(2019年1期)2019-02-21
科学大众·小诺贝尔(2018年12期)2018-02-22
新课程·下旬(2017年7期)2017-08-14
北京文学(2007年5期)2007-05-23
