
鹊鸲冬季和春季取食生态位初步研究
2016-12-09 08:27原宝东闫永峰
四川动物 2016年3期
原宝东, 闫永峰
(1.商丘师范学院生命科学学院,河南商丘476000;2.河池学院化学与生物工程学院,广西宜州546300)
鹊鸲冬季和春季取食生态位初步研究
原宝东1, 2, 闫永峰1*
(1.商丘师范学院生命科学学院,河南商丘476000;2.河池学院化学与生物工程学院,广西宜州546300)
2013年3—5月,2014年12月—2015年2月,在广西宜州利用直接观察法对鹊鸲Copsychussaularis的取食生态位进行分析,记录鹊鸲的数量、取食时间、食物种类、取食方式、取食生态位、取食方向等,共记录38 d,收集数据6 407个。结果表明:鹊鸲为杂食性鸟类,以果实和昆虫为主,冬季取食的植物有23种,取食生态位宽度为11.24;春季取食植物有22种,取食生态位宽度为12.19;春季和冬季的取食生态位重叠度为0.359。卡方独立性检验表明:春季和冬季鹊鸲的取食时间和食物种类的差异有统计学意义(P<0.05);取食高度、取食水平和取食方向等的差异无统计学意义(P>0.05)。鹊鸲的取食方式为啄食。鹊鸲的食物种类和取食时间受季节的影响较大,人为干扰是影响鹊鸲取食和活动的主要因素。
鹊鸲;食性;生态位宽度;生态位重叠度
鹊鸲Copsychussaularis是主要分布于我国长江以南地区的常见留鸟,共有2个亚种分化(赵正阶,2001)。国外见于欧亚大陆、印度次大陆、中南半岛、华莱士区(Wallace)和太平洋诸岛(蒋爱伍等,2009)。鹊鸲主要栖息于海拔2 000 m以下的低山、丘陵和山脚平原地带的次生林、竹林、林缘疏林灌丛和小块丛林等开阔地,尤其喜欢村寨和居民点附近的小块丛林、灌丛、果园以及耕地、路边和房前屋后的树林与竹林,甚至出现在城市公园和庭院树上(蒋仕仁,2003)。目前关于鹊鸲的研究相对较少,有学者在杭州对鹊鸲的声行为进行了观察和记录,并与印度哈里瓦地区的鹊鸲进行了比较分析,有学者分析了四川省东部峨城山区鹊鸲数量不断减少的原因(王协义,1999;刘绘,2003)。乱砍滥伐、猎捕和农药的使用以及土地使用性质的改变使鹊鸲的栖息地丧失,造成了鹊鸲的数量锐减(郑光美, 王岐山,1998)。
生态位是一个生物单位(个体、种群或物种)生存条件的总集合体,由于食物资源的质量直接影响鸟类的分布模式、种群密度、繁殖成功率和雏鸟存活率等,因而食物生态位研究至今仍然是生态位研究中非常活跃的一个领域(Thomas,2008)。动物的取食生态位研究对评估生境质量、预测栖息地负载量、制定合理的保护策略和资源管理方案等均具有重要意义。通过对鸟类在不同季节取食生态位的重叠度、相似性及宽度的计算,可深入认识鸟类对资源波动性的行为适应机制(杨晓君等,1994;王维禹等,2005;赵欣如等,2010)。因此,通过对广西宜州地区鹊鸲的取食行为进行直接观察,初步分析其春季和冬季取食生态位的宽度和重叠度,为以后开展鹊鸲的行为研究工作提供基础性资料,并对今后的保护和管理工作提出合理建议。
1 研究地区概况

2 研究方法
2.1 研究方法
2013年3—5月,2014年12月—2015年2月在野外调查采用样线法和样方法,发现鹊鸲后利用8×42倍的双筒望远镜采用随机取样法和扫描取样法定点观察鹊鸲的数量和取食行为,每2 d观察一次,观察时间段为:06∶00—09∶00、09∶00—12∶00、12∶00—15∶00、15∶00—18∶00。记录食物种类、取食方式、取食生态位及取食方向,每次观察可选取不同的时间段,也可以进行连续观察或全天观察。其中取食方式分为整吞、啄食、拾取及其他;取食生态位分为地面、灌丛及乔木树冠,其中乔木树冠的垂直取食生态位由上至下依次分为上层、中层及下层,水平取食生态位由外向内依次分为外侧、中侧及内侧;取食方向以正北方向为0°,正东方向为90°,正南方向为180°,正西方向为270°,共分为4个范围:315°~45°、45°~135°、135°~225°、225°~315°。将春季、冬季的各生态位参数进行卡方独立性检验。
所有数据处理均在Excel 2010和SPSS 13.0中进行。
2.2 种群分布模式
鹊鸲种群的个体在其生活空间中的位置状态和布局,采用统计样方结果中的方差/平均数(S2/m)来判断。S2/m=0为均匀分布,S2/m=1为随机分布,S2/m>1为集群分布。其中:
m=∑fx/N
S2=[∑fx2-(∑fx)2/N]/(N-1)
式中x为样本中含有的鹊鸲数量,f为出现频率(含有不同个体数样本的出现频率),N为样本总数。方差反映了样本与平均数的差异程度,方差越大,样方间鹊鸲种群密度的差异越大。
2.3 数据处理方法
2.3.1 取食生态位宽度 取食生态位宽度(feeding niche breadth)用Simpson多样性指数(B)表示(吴诗宝等,2005)。
B=1/∑Pi2
式中,Pi为实际利用的第i种食物在所有被利用的食物中所占的比例。
2.3.2 取食生态位重叠度 取食生态位重叠度(feeding niche overlap)用Levins(Ojk或Okj)指数表示。
Ojk=Okj=∑PijPik/(∑Pij2∑Pik2)1/2
式中,Pij和Pik分别为被j和k组鹊鸲利用的第i种食物在所有被利用的食物中所占比例。
3 结果及分析
3.1 鹊鸲种群的分布型
鹊鸲以单独觅食为主,偶尔有雌雄2只共同觅食,超过3只共同觅食时,它们之间会发生食物竞争的冲突,春季2只共同觅食发现比例明显超过冬季。将数据运用格局指数进行计算,结果表明,研究地区冬季的鹊鸲种群分布格局指数为0.99,为随机分布;春季的为1.13,为集群分布(表1)。

表1 鹊鸲种群分布的模式统计表
3.2 鹊鸲冬季、春季取食生态位宽度和重叠度
根据观察记录的数据,鹊鸲的取食行为数据见表2,并对不同季节的取食生态位宽度和重叠度进行分析。从结果可以看出鹊鸲为杂食性鸟类,以果实和昆虫为主,冬季取食的植物有23种,取食生态位宽度为11.24;春季取食的植物有22种,取食生态位宽度为12.19;春季和冬季的取食生态位重叠度为0.359。取食都是用喙啄食。鹊鸲喜欢在高大乔木、灌丛、草地和地面上独自取食,取食植物上的果实及地面上的落果,还取食植物枝条上和地面上的昆虫,有时也会在生活垃圾中寻找食物。鹊鸲春季和冬季的取食生态位宽度都较高,这与不同季节可供取食的果实的种类不同有关。
3.3 鹊鸲春季和冬季取食生态位比较分析
通过对鹊鸲春季和冬季的取食时间、食物种类、取食方式、取食高度、取食水平和取食方向等进行分析,结果见表3。
卡方独立性检验表明:鹊鸲春季和冬季的取食时间和食物种类的差异有统计学意义(P<0.05),取食高度、取食水平和取食方向等的差异无统计学意义(P>0.05),取食方式为啄食。取食果实种类不同可能与不同季节果实的成熟期不同有关;春季的主要取食时间在06∶00—09∶00,比例为50.13%,冬季的主要取食时间在09∶00—12∶00,比例为32.12%,取食时间受季节的影响较大,冬季早上寒冷,鹊鸲为了减少能量的散失会推迟取食时间,这是对环境的行为适应。鹊鸲春季和冬季在取食高度、取食水平和取食方向等的差异无统计学意义,这可能与研究区植物种类组成单一和挂果在外侧并朝阳有关。
4 讨论
4.1 鹊鸲的取食生态位
食物是影响鸟类生境利用和选择的主要因素,鸟类取食行为的适应机制是对食物丰富程度的响应。鸟类取食地选择的问题,不仅与鸟类本身的食性相关,也与栖息地结构以及食物资源状况相关。它可以揭示鸟类在其生存环境中的地位和作用,以及鸟类与环境之间的关系(杨晓君等,1994)。鹊鸲
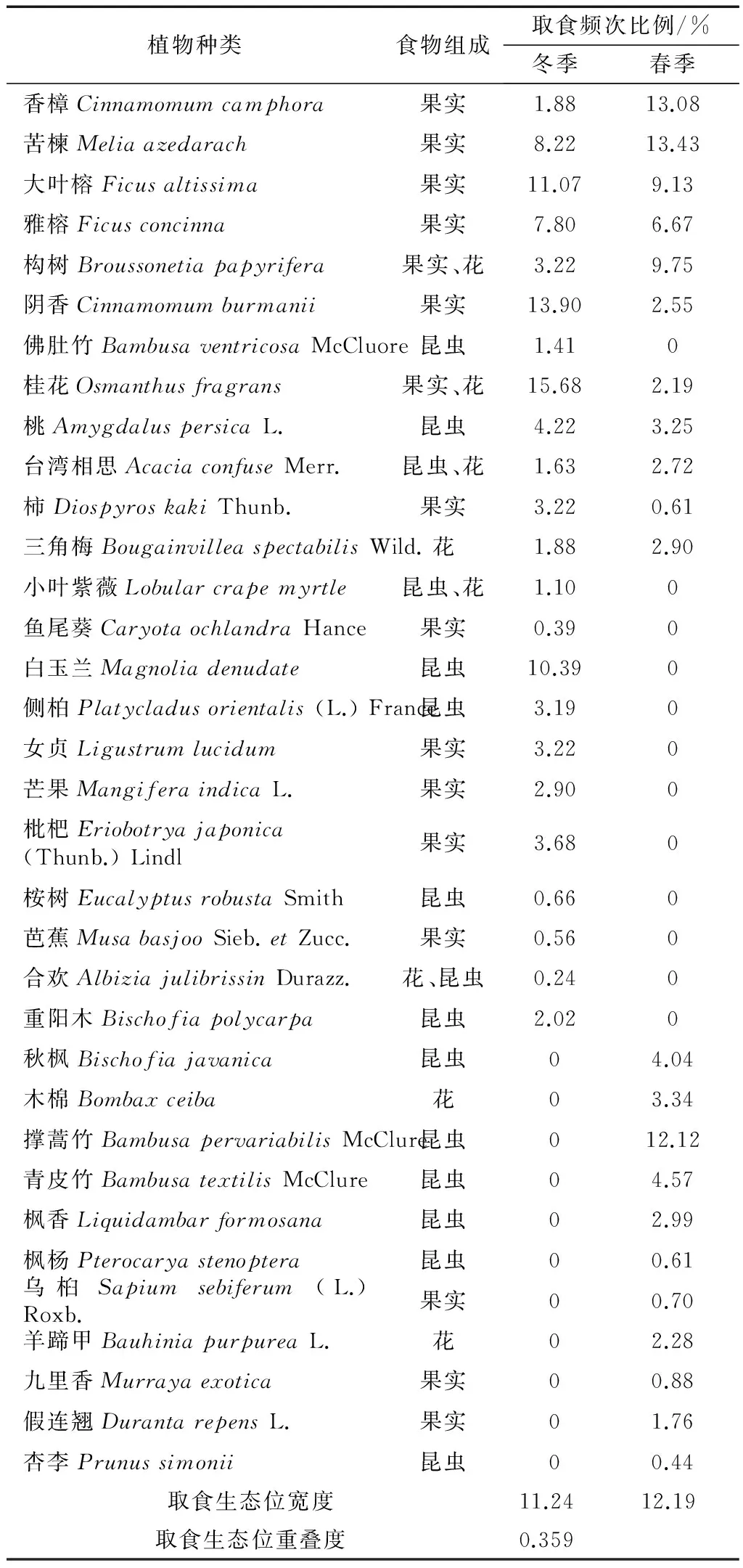
表2 鹊鸲冬季、春季取食生态位宽度和重叠度

表3 鹊鸲春季和冬季取食生态位比较分析(卡方独立性检验)
不同时段取食频次的差异可能与以下几方面原因有关:经过一晚的休息,鹊鸲消化道内的食物已经消化完毕;早晨人类的活动较少,对鸟类的干扰较少,这是造成早上取食频次相对较多的主要原因。以往的研究指出,大多数昼行性动物都具有早上进食的行为,以补充夜晚能量的消耗(庞秉璋,1981;韦明思等,2004)。鸟类的取食行为并不是一成不变的,它随着环境条件、季节、个体发育阶段不同以及新情况的出现而发生变化,是对环境适应的一个重要方面。然而随着季节的变化,冬季与春季相比温度要低一些,鹊鸲会推迟外出活动取食的时间,在其他鸟类的研究中也得出了类似的结果(Yagihashietal.,1998)。
生态位宽度是物种利用或趋于利用所有可利用资源状态,一个物种的生态位宽度越宽,其特化程度越小,也就是说该物种更倾向于一个泛化种;相反,一个物种的生态位宽度越窄,该种的特化程度越大,即更倾向于一个特化种。生态位重叠是个体与生态因子联系上的相似性,当物种利用同一资源(食物、营养成分、空间等)时,就会出现生态位重叠现象,生态位重叠程度能体现种群对共同资源的利用状况(van Den Wyngaertetal.,2003)。在研究地区,冬季挂果的植物相对较少,春季植物开始冒出新芽,有些春季结果实的植物为鹊鸲提供了更多的食物选择,这使得春季的取食生态位宽度略大于冬季。春季和冬季的取食生态位重叠度很低可能与植物果实的成熟季节不同有关(Kwitetal.,2004)。
在进化过程中,动物选择了适应本身条件发展的最佳取食策略。鹊鸲属于小型鸟类,2个季节能够取食的植物种类达到30多种,这与早期报道中小型鸟类食物选择性较小,通过提高觅食活动率、增加食物量、拓宽取食生态位相符(Barneaetal.,1991)。研究过程中发现,由于研究地区冬、春季昆虫资源相对较少(邓维安,2003),可以提供的肉食性食物有限,鹊鸲会更多地取食果实,这与早期报道杭州地区鹊鸲以肉食性为主的结果有差异(蒋仕仁,2003),同时从另一方面表明鹊鸲是一种适应能力较强的鸟类,能够根据食物资源的可利用性调整取食策略。一年之中夏季是昆虫多样性丰富的季节,秋季是果实成熟的主要季节,下一步开展季节变化对鹊鸲取食行为的影响研究是理解鹊鸲生活史策略的关键。
4.2 鹊鸲对种子的潜在传播作用
通过对2个季节鹊鸲的取食可以看出,鹊鸲主要以果实为主,也会取食昆虫、垃圾中的食物,这对抑制虫害起到了一定的作用。鹊鸲喜欢在树冠高处取食,这与植物树冠层采光充足、果实发育相对饱满、食物营养更多有关。许多植物通过鸟类传播果实,Barnea等(1991)研究表明,鸟类的消化过程能够明显提高种子的萌发率,甚至使种子由不能萌发变为能够萌发,原因是鸟类消化了果肉,而果肉中可能存在萌发抑制物质(Cipollini & Levey,1997)。鹊鸲在啄食果实的过程中会将种子附带吞入体内,未消化的种子以粪便的形式排出体外,从而成为了种子的传播者。在调查中也发现,在离母树很远的距离发现了苦楝Meliaazedarach、桂花Osmanthusfragrans等幼苗,这些幼苗很可能是借助食果动物的传播才得以在适宜的生境萌发。动物对种子传播的生态学是值得深入研究的重要领域(申仕康等,1998)。
研究过程中可见成对的鹊鸲在房顶上、空中打斗或嬉戏,停靠在电线杆上鸣叫,或者互相鸣叫。对于鹊鸲如何筑巢、繁殖,目前还没有相关的研究。研究期间曾发现鹊鸲在长穗鱼尾葵Caryotaochlandra和桂花上筑巢,深入研究鹊鸲的繁殖生物学才能更好地了解该物种的进化适应机制。
致谢:感谢河池学院本科生雷志栋和张小娟同学在野外数据收集过程中给予的帮助。
邓维安. 2003. 广西宜州市蝗虫名录[J]. 河池师专学报, 23(2): 97-98.
蒋爱伍, 于家捷, 杨浪, 等. 2009. 河池学院校园鸟类初步观察[J]. 河池学院学报, 29(5): 69-75.
蒋仕仁. 2003. 杭州鹊鸲声行为[J]. 四川动物, 22(3): 144-145.
刘绘. 2003. 拯救乖鸟鹊鸲[J]. 大自然探索, 22(6): 6.
庞秉璋. 1981.白头鹎的食性[J]. 动物学杂志, (4): 75-76.
覃勇荣, 覃宝山. 2008. 河池学院校园寄生植物的危害及其防治[J]. 河池学院学报, 28(2): 47-52.
申仕康, 胡晓立, 王跃华. 2008. 鸟类对猪血木种子的散布及在其保护中的意义[J]. 广西植物, 28(5): 650-654.
王维禹, 郭延蜀, 胡锦矗, 等. 2005. 白头鹎春季食性及取食空间生态位的初步研究[J]. 四川动物, 24(4): 466-468.
王协义. 1999. 鹊鸲缘何在减少[J]. 野生动物, 20(3): 5.
韦明思, 陈章和, 任海, 等. 2004. 鸟类和蚂蚁对桃金娘种子传播的初步研究[J]. 生物多样性, 12(5): 494-500.
吴诗宝, 刘迺发, 李有余, 等. 2005. 中国穿山甲的食性与觅食行为初步观察[J]. 应用与环境生物学报, 11(3): 337-341.
杨晓君, 文贤继, 杨岚. 1994. 笼养白腹锦鸡觅食活动的观察[J]. 动物学研究, 15(1): 49-54.
赵欣如, 肖雯, 张瑜. 2010. 野外观鸟手册[M]. 北京: 化学工业出版社: 6-10.
赵正阶. 2001. 中国鸟类志 下卷 雀形目[M]. 长春: 吉林科学技术出版社: 260-262.
郑光美, 王岐山. 1998. 中国濒危动物红皮书 鸟类[M]. 北京: 科学出版社.
Barnea A, Yom-Tov Y, Friedman J. 1991. Does ingestion by birds affects seed germination[J]. Functional Ecology, 5: 394-402.
Cipollini ML, Levey DJ. 1997. Secondary metabolites of fleshy vertebrate-dispersed fruits: adaptive hypotheses and implications for seed dispersal[J]. The American Naturalist, 150: 346-372.
Kwit C, Levey DJ, Greenberg CH,etal. 2004. Fruit abundance and local distribution of wintering hermit thrushes (Catharusguttatus) and yellow-rumped warblers (Dendroicacoronata) in south Carolina[J]. Auk, 121: 46-57.
Thomas K. 2008. Spatial smoothing techniques for the assessment of habitat suitability[J]. Environmental and Ecological Statistics, 15: 343-364.
van Den Wyngaert IJJ, Wienk LD, Sollie S,etal. 2003. Long-term effects of yearly grazing by moulting greylag geese (Anseranser) on reed (Phragmitesaustralis) growth and nutrient dynamics[J]. Aquatic Botany, 75: 229-248.
Yagihashi T, Hayashida M, Miyamoto T. 1998. Effects of bird ingestion on seed germination ofSorbuscommixta[J]. Oecologia, 114: 209-212.
传播科学信息的媒介 开展学术交流的平台

●主要栏目:研究报告、基础资料、野生动物保护与自然保护区、实验动物与动物实验、教学探索、综述与进展。
●读者对象:广大从事动物学、生物学和野生动物保护方面的科研、教学、管理、医卫等科技工作者,有关院校师生和业余爱好者。
●双月刊,大16开,每期 164页,精印彩色封面,逢单月末出版,2016年全年6期国内定价300元(每期50元),国外为每期20美元,全年120美元。
●订阅办法:从邮局汇款至本刊编辑部,写清订阅人姓名、地址、邮编,附言订阅数量即可。
电话/传真:028-85410485 电子邮件:scdwzz001@163.com scdwzz@vip.163.com
邮局汇款:四川省成都市望江路29号四川大学生命科学学院内《四川动物》编辑部
邮政编码:610064
银行汇款:开户银行:中国工商银行四川分行营业部东大支行 户名:四川省动物学会
帐号:4402298009000012596
Feeding Niche ofCopsychussaularisbetween Spring and Winter
YUAN Baodong1, 2, YAN Yongfeng1*
(1.College of Life Sciences, Shangqiu Normal University, Shangqiu, Henan Province 476000, China;2.Department of Chemistry and Life Sciences, Hechi University, Yizhou, Guangxi Zhuang Autonomous Region 546300, China)
Feeding niche of magpie robin (Copsychussaularis) was investigated from March to May 2013 and December 2014 to February 2015.In the fieldwork, number ofC.saularis, feeding time, food types, feeding behaviors, feeding niche and feeding direction were recorded.The results showed thatC.saulariswas omnivorous, but mainly fed on fruits and insects.Analysis of feeding niche breadth and overlap suggested that the value of feeding niche breadth was 11.24 in winter and 12.19 in spring, respectively; the value of feeding niche overlap was 0.359 between spring and winter.The feeding time and food types had significant differences between spring and summer (P<0.05), however, no significant difference was observed in other parameters (P>0.05).The feeding behaviors ofC.saulariscould be affected by food resources in different seasons.Human disturbance might be the main reason to influence its activity and feeding.
Copsychussaularis; feeding habit; niche breadth; niche overlap
2015-11-09 接受日期:2016-01-15 基金项目:河南省动物营养代谢与遗传改良创新型科技团队项目; 广西高校科学技术研究重点项目(KY2015ZD105); 商丘师范学院高层次人才科研启动项目(500014001)
原宝东(1981—), 男, 博士, 副教授, 主要从事动物学方面的研究, E-mail:yuanbao365@163.com
*通信作者Corresponding author,E-mail:yanyf01@sina.com
10.11984/j.issn.1000-7083.20150352
Q958.1; Q959.7
A
1000-7083(2016)03-0426-05
猜你喜欢
学与玩(2022年9期)2022-10-31
文苑(2020年12期)2020-04-13
小太阳画报(2019年1期)2019-06-11
小学生必读(低年级版)(2017年5期)2017-08-12
爆笑show(2016年2期)2016-04-11
少儿科学周刊·儿童版(2015年6期)2015-11-24
医学研究杂志(2015年5期)2015-06-10
人生十六七(2015年5期)2015-02-28
爆笑show(2014年7期)2014-09-03
销售与市场·管理版(2009年21期)2009-09-03
