
外源NADP+和NADPH缓解番茄幼苗NaCl胁迫伤害的初步研究
2017-03-30 09:41莘冰茹张健伟刘慧英
西北农业学报 2017年3期
莘冰茹,王 松,周 艳,张健伟,刘慧英
外源NADP+和NADPH缓解番茄幼苗NaCl胁迫伤害的初步研究
莘冰茹1,2,王 松1,2,周 艳1,2,张健伟1,2,刘慧英1,2
(1.石河子大学 农学院园艺系,新疆石河子 832003;2.新疆生产建设兵团特色果蔬栽培生理与种资资源利用重点实验室, 新疆石河子 832003)
采用营养液培法,以加工番茄品种‘里格尔87-5’为材料,通过对NaCl 胁迫下加工番茄幼苗叶片分别喷施5、10、15 μmol·L-1NADP+和NADPH,研究外源NADP+、NADPH对NaCl胁迫下加工番茄幼苗生长及其抗逆生理指标的影响。结果表明:外源喷施NADP+、NADPH均能改善NaCl处理下加工番茄幼苗的生长性状,增加叶片叶绿素含量,提高根系活力和叶片过氧化物酶(POD)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和抗坏血酸过氧化物酶(APX)活性,降低丙二醛(MDA)质量摩尔浓度。通过不同处理综合指标总加权值比较评价得出: NaCl胁迫下喷施5 μmol·L-1NADPH的效果最佳,其次为10 μmol·L-1NADP+。可见,外源喷施NADP+、NADPH通过增加色素含量,可提高抗氧化酶活性,降低膜脂过氧化水平,在一定程度上缓解盐胁迫对植株的伤害,从而增强对盐胁迫的适应性。
盐胁迫;加工番茄;NADP+;NADPH
新疆是中国加工番茄主产区,总产量占全国加工番茄制品产量的90%以上[1]。但新疆丰富的含盐母质、特殊的气候条件和地理格局导致新疆加工番茄现有耕地的土壤盐渍化日益严重,已成为新疆当前发展红色产业的主要障碍[2]。近年来,通过使用外源物质来缓解盐胁迫对作物的伤害已成为有效克服土壤盐渍化的途径之一[3-4]。
还原型辅酶Ⅱ(reduced form of nicotinamide-adenine dinucleotide phosphate,NADPH)是一种极为重要的核苷酸类辅酶,其来源主要有2条:一条是通过位于线粒体的转氢酶催化还原型辅酶Ⅰ(reduced form of nicotinamide-adenine dinucleotid,NADH)和氧化型辅酶Ⅱ(nicotinamide adenine dinucleotide phosphate,NADP+)生成NADPH;另一条是通过NADPH依赖性酶,直接提供氢离子给NADP+,生成NADPH[5]。NADPH作为供氢体在维持细胞中还原型谷胱甘肽(Reduced glutathione,GSH)的含量以及氧化还原平衡和硫氧还蛋白(Thioredoxin,Trx)系统的活性,参与细胞信号转导、抗氧化防御中发挥重要作用[6-8]。许多研究已表明NADPH/NADP+、GSH/ GSSG (Oxidizedglutathione,氧化型谷胱甘肽)和AsA(reduced ascorbic acid,还原型抗坏血酸)/DHA(dehydroascorbate,氧化型抗坏血酸)是3个相互关联的重要氧化还原对,在环境胁迫条件下紧密协调来抵抗胁迫。NADPH氧化还原的同时伴随着细胞壁内部多聚糖阿魏酸残基的铰链,加强了细胞壁对病原物的抵抗能力[9]。通过对拟南芥中1个编码NADPH氧化酶类似物的基因 rhd2进行突变,发现NADPH氧化酶激活质膜Ca2+内流通道,促进细胞的生长,从而调控植物发育[10]。但目前有关NADP+和NADPH如何影响盐胁迫下加工番茄幼苗生长及其缓解盐害、提高盐适应性上的研究尚未见报道。
因此,本试验通过外源喷施不同浓度NADP+和NADPH的方法,研究外源NADP+和NADPH对NaCl 胁迫下加工番茄幼苗的生长、生理指标和耐盐性的影响,以期为利用外源物质减轻盐胁迫伤害提供理论依据。
1 材料与方法
1.1 材料及试验设计
选用加工番茄(Solanumlycopersicum)品种‘里格尔87-5’作为供试材料,试验于石河子大学实验站温室内进行。采用营养液培养的方法。挑选饱满、一致的种子,经温汤浸种、催芽后播于含有混合基质(草炭和蛭石的体积比为2∶1)的72孔穴盘中育苗,在加工番茄4叶1心时期选取生长健壮、长势一致的幼苗移入带泡沫盖板的12 L塑料水桶中,加入10 L去离子水配制的Hoagland营养液(pH=6.2),用空气压缩机保持全天通气。待幼苗6叶1心时期进行处理,以100 mmol·L-1NaCl 作为模拟盐胁迫所用的浓度,直接加入营养液中,NADPH和NADP+2种外源物质以叶片喷施的方式于每日10:00 喷施,其中以NADPNa2作为NADP+供体。共设置8个处理,分别为(1)对照(CK):不加NaCl,叶面喷施蒸馏水;(2)NaCl:NaCl和叶面喷施蒸馏水;(3)T5:NaCl+5 μmol·L-1NADP+;(4)T10: NaCl+10 μmol·L-1NADP+;(5)T15: NaCl+15 μmol·L-1NADP+;(6)N5: NaCl+5 μmol·L-1NADPH;(7)N10: NaCl+10 μmol·L-1NADPH; (8)N15: NaCl+15 μmol·L-1NADPH。采用随机区组设计,每个处理重复3次。于处理后第9天进行取样测定。
1.2 测定指标及方法
随机抽取加工番茄植株,用直尺测量株高,游标卡尺测量茎粗,称量地上部与地下部及整株干、鲜质量,计算根冠比(根冠比 = 地下部干质量/地上部干质量 × 100%)。
叶绿素质量分数采用李和生[11]的丙酮提取法;根系活力的测定采用TTC法[11];丙二醛(MDA)质量摩尔浓度测定参照Draper等[12]的方法;超氧化物歧化酶(SOD)活性的测定参照Rao等[13]的方法;过氧化氢酶(CAT)、过氧化物酶(POD)活性的测定参照Zhang[14]的方法;抗坏血酸过氧化物酶(APX)活性测定参照 Nakano等[15]的方法并加以改进。
1.3 数据处理
采用 Excel 2003 和 SPSS 17.0对数据进行分析,差异显著性用Duncan’s检验法进行多重比较(P<0.05),用OriginPro 7.5进行数据绘图。参照周通等[16]方法计算隶属函数值,即某个性状的隶属函数值等于它的该性状测定值减去该性状最小值与该性状的极差值的比值,可表示为:R(Xi)=(Xi-Xmin)/(Xmax-Xmin),若某一性状与耐盐性为负相关,则利用反隶属函数进行计算,即R(Xi)= 1-(Xi-Xmin)/(Xmax-Xmin)。
2 结果与分析
2.1 外源NADP+和NADPH对NaCl胁迫下番茄幼苗生长的影响
表1显示,NaCl胁迫下,加工番茄幼苗株高、茎粗、整株鲜质量及干质量均显著低于未经 NaCl胁迫处理的正常对照植株,分别下降31.27%、29.11%、35.21%、57.67%;与NaCl胁迫处理相比,叶片喷施5 μmol·L-1NADP+显著提高NaCl胁迫下幼苗的株高和干质量,对茎粗、鲜质量无显著影响。而叶片分别喷施10、15 μmol·L-1NADP+和5、10、15 μmol·L-1NADPH均显著提高NaCl胁迫下幼苗的株高、茎粗、鲜质量及干质量,有效缓解NaCl 造成的生长抑制;在喷施6个不同浓度NADP+、NADPH处理中,以喷施5 μmol·L-1NADPH(N5)效果最好,加工番茄株高、茎粗、整株鲜质量及干质量分别比NaCl胁迫高31.64%、32.14%、43.64%、94.35%,其次为10 μmol·L-1NADPH (N10);3个喷施NADP+处理中以喷施浓度为10 μmol·L-1NADP+处理(T10)效果最好。
同时,NaCl胁迫下幼苗根冠比最大,较对照显著提高;6个喷施不同浓度NADP+、NADPH处理中,除喷施5 μmol·L-1NADP+处理对NaCl胁迫下幼苗的根冠比无显著影响外,其他5个浓度处理均显著降低NaCl胁迫下幼苗的根冠比。说明外源喷施适宜浓度的NADP+(10、15 μmol·L-1)、NADPH(5、10、15 μmol·L-1)能有效增加植株地上部生物量的积累。
2.2 外源NADP+和NADPH对NaCl胁迫下番茄幼苗根系活力的影响
如图1所示,与对照相比,NaCl处理显著地降低幼苗根系活力,而6个不同浓度的外源NADP+、NADPH处理均显著提高NaCl胁迫下的加工番茄幼苗根系活力,其中以N5处理效果最好,N10处理次之,N15和T10处理间无显著差异,但均显著高于T5处理,T10处理的根系活力最低。整体来看,NADPH处理下的植株根系活力高于NADP+处理的。
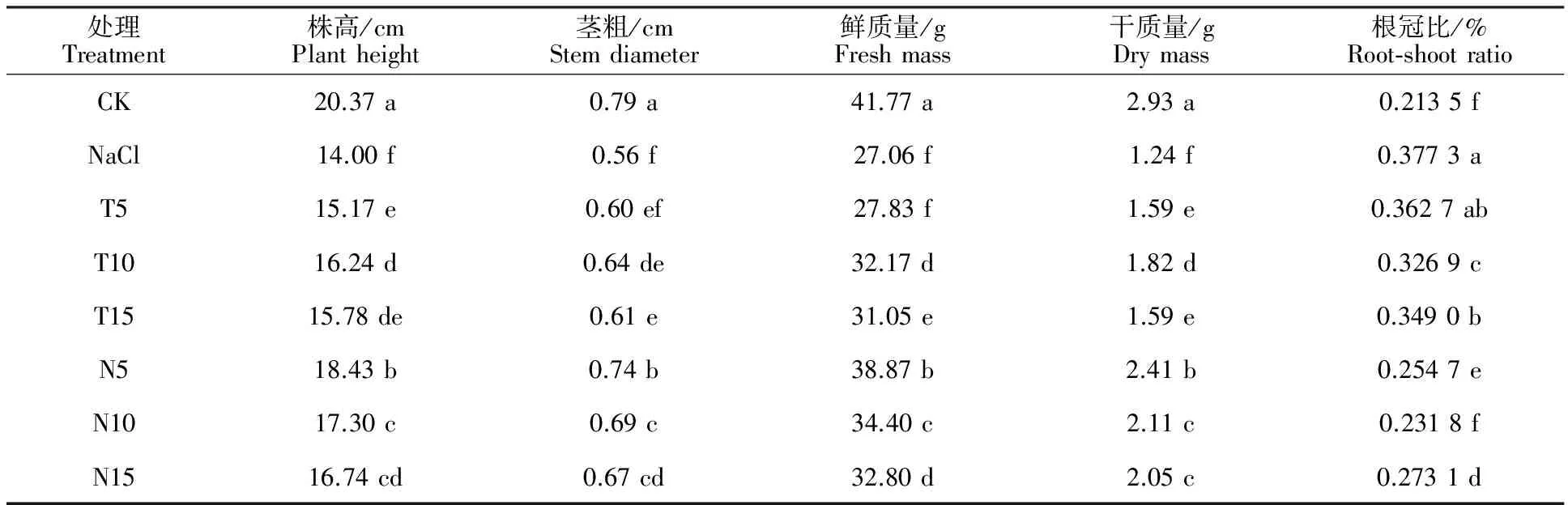
表1 不同浓度NADP+和NADPH对NaCl胁迫下番茄幼苗农艺性状的影响Table 1 Effects of different NADP+ and NADPH concentration on the agronomic traits of processing tomato seedlings under NaCl stress
注:不同字母表示处理间差异显著(P<0.05)。下同。
Note:Different letters indicate significant differences between treatments(P<0.05).The same below.
2.3 不同浓度NADP+和NADPH对NaCl胁迫下番茄幼苗叶绿素含量的影响
如表2所示,NaCl处理下,加工番茄幼苗叶绿素 a、叶绿素 b含量及叶绿素总量显著低于正常对照处理;叶片喷施5、10、15 μmol·L-1NADP+可以使NaCl胁迫下幼苗叶片叶绿素 a、叶绿素 b和叶绿素总含量分别提高1.4~1.8倍、1.7~2.1倍、1.4~1.8倍;外源喷施5、10和15 μmol·L-1NADPH则可以使NaCl胁迫下幼苗叶片叶绿素 a、叶绿素 b和叶绿素总量分别提高1.5~2.1倍、2.0~2.9倍、1.6~2.2倍;在6个喷施不同浓度NADP+、NADPH处理中,以喷施5 μmol·L-1NADPH(N5)效果最佳,其次为喷施10 μmol·L-1NADP+(T10)和10 μmol·L-1NADPH(N10)处理。2.4 不同浓度NADP+和NADPH对NaCl胁迫下番茄幼苗叶片MDA质量摩尔浓度的影响
从图2 可以看出,NaCl处理下加工番茄幼苗叶片的MDA质量摩尔浓度较对照显著提高,且是所有处理中最高的;叶片喷施3个不同浓度的NADP+均显著降低NaCl胁迫下幼苗叶片的MDA质量摩尔浓度,其中T5和T15处理间无显著差异,但均显著高于T10处理;叶片喷施3个不同浓度的NADPH亦均显著降低NaCl胁迫下幼苗叶片的MDA质量摩尔浓度,其中N10和N15处理间无显著差异,但均显著高于N5处理;6个不同浓度的NADP+和NADPH处理中,以T10和N5处理的MDA质量摩尔浓度最低,且两处理间无显著差异。

不同字母表示处理间差异显著(P<0.05)。下同 Different letters indicate significant differences between treatments(P<0.05).The same below

表2 不同浓度NADP+和NADPH对NaCl胁迫下番茄幼苗叶绿素含量的影响Table 2 Effects of NADP+ and NADPH concentrations on chlorophyll content of tomato seedlings under NaCl stress
2.5 不同浓度NADP+和NADPH对NaCl胁迫下番茄幼苗抗氧化酶的影响
图3显示,与对照相比,NaCl胁迫使幼苗叶片SOD活性显著升高。叶片分别喷施6个不同浓度的NADP+和NADPH后均进一步提高NaCl胁迫下幼苗叶片的SOD活性;喷施不同浓度NADPH的3个处理的SOD 活性均显著低于3个喷施不同浓度NADP+处理的,其中T5显著高于T10和T15处理,N10显著高于N15处理,但与N5无显著差异。
NaCl胁迫处理下,加工番茄幼苗POD、CAT和APX活性均较对照显著降低,喷施不同浓度的NADP+和NADPH均使NaCl胁迫下幼苗叶片的3种酶活性显著提高,但不同处理下3种酶活性的提高幅度有差异;在6个不同浓度的NADP+和NADPH处理中,N5处理的POD活性最高,显著高于其他5个处理,N15处理的POD活性最低,显著低于其他5个处理,T5和T10处理间无显著差异,但均显著高于T15和N10处理的;T10和T15处理的CAT活性显著高于T5、N10 和N15处理,N15处理的CAT活性最低,显著低于其他5个处理,N5处理与T10、T15和N10处理间无显著差异;T5和T10处理的APX活性较高,显著高于其他4个处理,而T15和N5处理的APX活性显著高于N10和N15处理。

图2 不同浓度NADP+和NADPH对NaCl胁迫下番茄幼苗叶片MDA质量摩尔浓度的影响Fig.2 Effects of different NADP+ and NADPH concentrations on MDA molality in leaves of tomato seedlings under NaCl stress

图3 不同浓度NADP+和NADPH对NaCl胁迫下番茄幼苗叶片SOD、POD、CAT和APX活性的影响Fig.3 Effects of different NADP+ and NADPH concentrations on activities of SOD,POD,CAT and APX in tomato leaves under NaCl stress
2.6 不同处理番茄幼苗耐盐性差异综合比较
为了综合评价不同处理下加工番茄幼苗耐盐性差异,采用多性状指标的隶属函数值法进行评价。不同处理12个指标加权值和总加权值大小及其排序见表3。其中除根冠比和MDA指标指数对加工番茄耐盐性不利而采用反隶属函数值法计算外,其他指标均对加工番茄耐盐性有利,因此,采用隶属函数值法计算函数值。
从表3可以看出,综合加权值评价处理的抗盐性强弱顺序为N5(NaCl+5 μmol·L-1NADPH)> CK > T10(NaCl+10 μmol·L-1NADP+)> N10(NaCl+10 μmol·L-1NADPH)> T5 (NaCl+5 μmol·L-1NADP+)> T15(NaCl+15 μmol·L-1NADP+)> N15(NaCl+15 μmol·L-1NADPH)> NaCl 。

表3 综合指标的加权隶属函数值Table 3 Subordinate function values of comprehensive indexes
3 讨论与结论
植物在盐胁迫下最常见和最显著的生理过程是生长受到抑制[17-19]。目前,已有大量研究表明盐胁迫会严重影响植物的生长发育,造成生物量积累减少[20]。而选用一些外源物质对植株进行处理,已成为近年来有效提高植物耐盐碱能力的研究热点[21]。本研究中,NaCl胁迫下番茄幼苗株高、茎粗、整株鲜质量及干质量与对照相比下降29%~58%,同时根冠比增加,说明盐分通过抑制和诱导多种酶系统来影响植物的正常生长[22],并且对植物地上部分生长的抑制作用大于根系[23]。而喷施5~15 μmol·L-1NADP+、NADPH显著缓解NaCl的抑制作用,有效促进番茄幼苗生长。这与Chumpolkulwong等[24]报道的通过添加 NADPH能够促进虾青素生物合成的结果相似。盐胁迫既能直接降低根系活力,使水分和矿质营养的吸收受阻,也能通过减少光合产物向根系的分配来影响根的生长,进而影响叶片的光合能力[25]。叶绿素是光合作用的物质基础,其质量分数的多少直接影响植株光合作用的强弱。本试验中,NaCl胁迫下番茄幼苗根系活力降低,叶片叶绿素含量显著低于正常对照,说明叶绿体光能吸收、传递和转换能力下降,影响植物的光合性能,减少光合产物向地下部的运输。这与王颖等[26]在菠菜,赵莹等[27]在玉米上得到结果相似。而外源喷施5~15 μmol·L-1NADP+、NADPH能诱发叶绿素的积累,作为 PSⅡ反应中心的天线色素叶绿素a 和叶绿素b含量的增加促进植株的光合能力[28],最终提高光合产物的积累和生物量的增加。

值得注意的是,通常认为细胞内的氧化还原状态能够调控细胞的多种功能,而维持高的还原库力和还原状态是植物抵抗生物和非生物逆境胁迫,保护植物免受氧化胁迫的必要条件[36]。因此AsA、GSH、NADPH 含量及 AsA/DHA、GSH/GSSG、NADPH/NADP+的比值常被看作是反映胞内氧化还原状态的指标。已有研究证明外源AsA[37]和GSH[38]通过介导上调内源AsA/DHA、GSH/GSSG、NADPH/NADP+的比值以增强对逆境的适应性,而GSSH 和DHA则会加剧逆境的氧化胁迫[39]。NADP+是氧化态的辅酶Ⅱ,但本试验中,喷施NADP+未对NaCl胁迫下番茄幼苗植株产生抑制,却不同程度地缓解了盐胁迫造成的伤害,推测这可能与本试验喷施NADP+的浓度、作用时间以及NaCl胁迫因子和胁迫强度等有关,也可能是植物的一种适应性反应,植物细胞氧化力提高激活植株活性氧的防御抵抗能力和NADP+向NADPH转化合成途径。具体原因还有待进一步研究。
柴媛媛等[40]用隶属函数法对甜高粱萌发期的耐盐性进行综合评价,得出10个品种甜高粱的耐盐顺序,认为隶属函数值法是一种较为理想的评价方法,为今后甜高粱耐盐性的进一步鉴定提供借鉴。本研究通过对番茄12个耐盐生理生化指标进行综合加权值比较,评价出番茄耐盐性强弱顺序依次为NaCl+5 μmol·L-1NADPH > CK > NaCl+10 μmol·L-1NADP+> NaCl+10 μmol·L-1NADPH > NaCl+5 μmol·L-1NADP+> NaCl+15 μmol·L-1NADP+> NaCl+15 μmol·L-1NADPH > NaCl 。可以看出,外源喷施5 μmol·L-1NADPH对加工番茄抗盐性各项指标最有利,与正常对照无显著差异;外源喷施NADP+则是浓度为10 μmol·L-1处理效果较其他两组较好。
综上所述,外源喷施NADP+和NADPH均能通过增加色素含量、提高抗氧化酶活性、降低膜脂过氧化水平而在一定程度上缓解盐胁迫对植株生长的抑制和伤害,从而增强对盐胁迫的适应性,其中以NaCl胁迫下喷施5 μmol·L-1NADPH的效果最佳,其次为10 μmol·L-1NADP+。
Reference:
[1] 庞胜群,王祯丽,张 润,等.新疆加工番茄产业现状及发展前景[J].中国蔬菜,2005(2):39-41.
PANG SH Q,WANG ZH L,ZHANG R,etal.The present situation and development prospect of processing tomato industry in Xinjiang[J].ChinaVegetables,2005(2):39-41(in Chinese ).
[2] 马 龙,喻晓强,樊新民,等.外源硒对盐胁迫下加工番茄幼苗叶片抗氧化系统的调控效应[J].北方园艺,2013(3):4-8.
MA L,YU X Q,FAN X M,etal.Research on physiological property variation on wild fern under different storage conditions[J].NorthernHorticulture,2013(3):4-8(in Chinese with English abstract).
[3] MOHAMMAD J M,HALIMEH H,VAHID N,etal.Exogenous application of penconazole regulates plant growth and antioxidative responses in salt-stressedMenthapulegiumL[J].JournalofPlantInteractions,2014,9(1):791-801.
[4] SHU S,SUN J,GUO S R,etal.Effects of exogenous putrescine on PSⅡ photochemistry and ion distribution of cucumber seedlings under salt stress[J].ActaHorticulturaeSinica,2010,37(7):1065-1072.
[5] FREEMAN H,SHIIMOMURA,COX R D,etal.Nicotinamide nucleotide transhydrogenase: a link between insulin secretion,glucose metabolism and oxidative stress[J].BiochemicalSocietyTransactions,2006,34(5):806-810.
[6] YING W.NAD+/NADH and NADP+/NADPH in cellular functions and cell death: regulation and biological consequences[J].AntioxidRedoxSignal,2008,10(2):179-206.
[7] KIRSCH M,DE G H.NAD(P)H,a directly operating antioxidant?[J].FasebJournal,2001,15(9):1569-1574.
[8] NAKAMURA H.Thioredoxin and its related molecules: update 2005[J].AntioxidRedoxSignal,2005,7(5-6):823-828.
[9] FRY S C,WILLIS S C,PATERSON A E.Intraprotoplasmic and wall-localised formation of arabinoxylan-bound diferulates and larger ferulate coupling-products in maize cell-suspension cultures[J].Planta,2000,211(5):679-692.
[10] FOREMAN J,DEMIDCHIK V,BOTHWELL J H,etal.Reactive oxygen species produced by NADPH oxidase regulate plant cell growth[J].Nature,2003,422:442-446.
[11] 李合生.植物生理生化实验原理与技术[M].北京:高等教育出版社,2001.
LI H SH.Plant Physiological and Biochemical Principles and Techniques[M].Beijing:Higher Education Press,2001(in Chinese).
[12] DRAPER H H,HADLEY M.Malondialdehyde determination as index of lipid peroxidation[J].MethodEnzymol,1990,186:421-431.
[13] RAO K,SRESTY T.Antioxidant parameters in the seedlings of pigeon pea(CajanuscajanL.Mill spaugh) in response to Zn and Ni stresses[J].PlantScience,2000,157:113-128.
[14] ZHANG X Z.The Measurement and Mechanism of Lipid Peroxidation and SOD,POD and CAT Activities in Biological System[C]//Research Methodology of Crop Physiology[M].Beijing:Agriculture Press,1992:208-211.
[15] NAKANO Y,ASADA K.Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts[J].PlantandCellPhysiology,1981,22(5):867-880.
[16] 周 通,王孝纯,王 艳,等.寒地水稻抗氮胁迫能力的筛选研究[J].中国农学通报,2001,27(33):127-136.
ZHOU T,WANG X CH,WANG Y,etal.Screening of different japonica rice germplasm tolerance to nitrogen deficiency in cold region of China[J].ChineseAgriculturalScienceBulletin,2001,27(33):127-136(in Chinese with English abstract).
[17] TAKEMURA T,HANAGATA N,SUGIHARA K,etal.Physiological and biochemical responses to salt stress in the mangrove,Bruguieragymnorrhiza[J].AquaticBotany,2000,68:15-28.
[18] ZHU J K.Salt and drought stress signal transduction in plants[J].AnnualReviewofPlantBiology,2002,53:247-273.
[19] MUNNS R,TESTER M.Mechanisms of salinity tolerance[J].AnnualReviewofPlantBiology,2008,59:651-681.
[20] ZHANG Y,HU X H,SHI Y,etal.Beneficial role of exogenous spermidine on nitrogen metabolism in tomato seedlings exposed to saline-alkaline stress[J].JournaloftheAmericanSocietyforHorticulturalence,2013,38(1):38-49.
[21] 张振兴,孙 锦,郭世荣,等.钙对盐胁迫下西瓜光合特性和果实品质的影响[J].园艺学报,2011,38(10):1929-1938.
ZHANG ZH X,SUN J,GUO SH R,etal.Effects of supplemental calcium on the photosynthetic characteristics and fruit quality of watermelon under salt stress[J].ActaHorticulturaeSinica,2011,38(10):1929-1938(in Chinese with English abstract).
[22] 王东明,贾 媛,崔继哲.盐胁迫对植物的影响及植物盐适应性研究进展[J].中国农学通报,2009,25(4):124-128.
WANG D M,JIA Y,CUI J ZH.Advances in research on effects of salt stress on plant and adaptive mechanism of the plant to salinity[J].ChineseAgriculturalScienceBulletin,2009,25(4):124-128(in Chinese with English abstract).
[23] 朱 义,谭贵娥,何池全,等.盐胁迫对高羊茅(Festucaarundinacea)幼苗生长和离子分布的影响[J].生态学报,2007,27(12): 5447-5454.
ZHU Y,TAN G E,HE CH Q,etal.Effect of salinization on growth and ionhomeostasis in seedlings ofFestucaarundinacea[J].ActaEcologicaSinica,2007,27(12):5447-5454(in Chinese with English abstract).
[24] CHUMPOLKULWONG N,ISHII H,KAKIZONO T.Enzymatic conversion of b-carotene to astaxanthin by cell-extracts of a green algaHaematococcuspluvialis[J].BiotechnologyLetters,1997,19(19):443-446.
[25] 王兴银,张福墁.弱光对日光温室黄瓜光合产物分配的影响[J].中国农业大学学报,2000,5(5):36-41.
WANG X Y,ZHANG F M.Influence of poor light on the distribution pattern of assimilate of cucumber in solar greenhouse[J].JournalofChinaAgriculturalUniversity,2000,5(5):36-41(in Chinese with English abstract).
[26] 王 颖,郭世荣,束 胜,等.外源亚精胺对盐胁迫下菠菜叶绿素合成前体含量的影响[J].西北植物学报,2015,35(10):2026-2034.
WANG Y,GUO SH R,SHU SH,etal.Effects of exogenous spermidine on chlorophyll precursors content of spinach plants under salt stress[J].ActaBotanicaBoreali-occidentaliaSinica,2015,35(10):2026-2034(in Chinese with English abstract).
[27] 赵 莹,杨克军,赵长江,等.外源糖调控玉米光合系统和活性氧代谢缓解盐胁迫[J].中国农业科学,2014,47(20):3962-3972.
ZHAO Y,YANG K J,ZHAO CH J,etal.Alleviation of the adverse effects of salt stress by regulating photosynthetic system and active oxygen metabolism in maize seedlings[J].ScientiaAgriculturaSinica,2014,47(20):3962-3972(in Chinese with English abstract).
[28] WANG L J,JIANG W B,HUANG B J.Promotion of 5-aminolevulinic acid on photosynthesis of melon(Cucumismelo) seedlings under low light and chilling stress conditions[J].PhysiolPlantarum,2004,121(2):258-264.
[29] KANAZAWA S,SANO S,KOSHIBA T,etal.Changes in antioxidative enzymes in cucumber cotyledons during natural senescence: comparison with those during dark-induced senescence[J].PhysiologiaPlantarum,2000,109(2):211-216.
[30] PARIDA A K,DAS A B.Salt tolerance and salinity effects on plants: a review[J].EcotoxicologyandEnvironmentalSafety,2005,60(3): 324-349.
[31] FOYER C H,DESCOURVIERES P,KUNERT K J.Protection against oxygen radicals:An important defence mechanism studied in transgenic plants[J].PlantCellandEnvironmen,1994,17(5):507-523.
[32] 李文兵,王燕凌,李 芳,等.水分胁迫下多枝柽柳体内活性氧与保护酶的关系[J].新疆农业大学学报,2007,30(1):30-34.
LI W B,WANG Y L,LI F,etal.Relationship between sctive oxygen and protective enzymes in tamarix ramosissima under water stress[J].JournalofXinjiangAgriculturalUniversity,2007,30(1):30-34(in Chinese with English abstract).
[33] 马彦霞,张国斌,颉建明,等.外源谷胱甘肽对辣椒自毒作用下辣椒幼苗叶片活性氧清除系统的影响[J].西北植物学报,2009,29(7):1380-1386.
MA Y X,ZHANG G B,XIE J M,etal.Effects of glutathione on active oxygen scavenging system in leaves of pepper seedlings under autotoxicity[J].ActaBotanicaBoreali-occidentaliaSinica,2009,29(7):1380-1386(in Chinese with English abstract).
[34] SREENIVASULU N,GRIMM B,WOBUS U,etal.Differential response of antioxidant compounds to salinity stress in salt-tolerant and salt-sensitive seedlings of foxtail millet(Setariaitalica)[J].PhysiologiaPlantarum,2000,109(4):435-442.
[35] 郭 剑,李彩凤,陈 明,等.碳酸钠胁迫对甜菜幼苗生长及保护酶的影响[J].植物生理学报,2015,51(6):840-846.
GUO J,LI C F,CHEN M,etal.Effects of Na2CO3stress on the growth and antioxidant enzymes ofBetavulgarisseedlings[J].PlantPhysiologyJournal,2015,51(6):840-846(in Chinese with English abstract).
[36] FOYER S H,THEODOULOU F L,DELROT S.The functions of interand intracellular glutathione transport systems in plants[J].TrendsinPlantScience,2001(6):486-492.
[37] 丁继军,潘远智,刘柿良,等.外源 AsA 对土壤重金属镉胁迫下石竹(Dianthuschinensis)幼苗生长的影响[J].农业环境科学学报,2013,32(8):1520-1528.
DING J J,PAN Y ZH,LIU SH L,etal.Effects of exogenous AsA on the growth ofDianthuschinensisseedlings under soil Cd stress[J].JournalofAgro-EnvironmentScience,2013,32(8):1520-1528(in Chinese with English abstract).
[38] 刘会芳,何晓玲,肖春燕,等.外源GSH对NaCl胁迫下番茄幼苗光合特性及碳同化关键酶基因表达的影响[J].应用生态学报,2014,25(9):2637-2644.
LIU H F,HE X L,XIAO CH Y,etal.Effects of exogenous GSH on photosynthetic characteristics and expression of key enzyme genes of CO2assimilation in leaves of tomato seedlings under NaCl stress[J].ChineseJournalofAppliedEcology,2014,25(9):2637-2644(in Chinese with English abstract).
[39] 马 进,郑 钢,裴翠明,等.抗坏血酸-谷胱甘肽循环在紫花苜蓿突变体耐盐性中的作用[J].植物生理学报,2015,51(10): 1749-1756.
MA J,ZHENG G,PEI C M,etal.The function of ascorbate-glutathione cycle in salt tolerance of alfalfa mutant[J].PlantPhysiologyJournal,2015,51(10):1749-1756(in Chinese with English abstract).
[40] 柴媛媛,史团省,谷卫彬.种子萌发期甜高粱对盐胁迫的响应及其耐盐性综合评价分析[J].种子,2008,27(2):43-47.
CHAI Y Y,SHI T SH,GU W B.Response to salt stresses and salt resistance evaluation of sweet sorghum during seed germination stage[J].Seed,2008,27(2):43-47(in Chinese with English abstract).
(责任编辑:潘学燕 Responsible editor:PAN Xueyan)
Exogenous NADP+and NADPH Alleviate the Damage of NaCl Stress on Tomato Seedlings
XIN Bingru1,2,WANG Song1,2,ZHOU Yan1,2,ZHANG Jianwei1,2and LIU Huiying1,2
(1.Department of Horticulture,College of Agronomy,Shihezi University,Shihezi Xinjiang 832000,China; 2.Key Laboratory of Special Fruits and Vegetables Cultivation Physiology and Germplasm Resources Utilization of Xinjiang Production and Construction Corps,Shihezi Xinjiang 832003,China)
The objectives of this hydroponic experiment were to determine the effects of exogenous NADP+and NADPH on the growth of processing tomato(Solanumlycopersicumcv.Ligeer 87-5) seedlings under NaCl stress.The tomato leaves were sprayed with either NADP+or NADPH at concentrations of 5,10,and 15 μmol·L-1.The results showed that exogenous NADP+and NADPH reduced malondialdehyde (MDA) mass molality and increased the growth of tomato seedlings under NaCl stress.Exogenous NADP+and NADPH increased the activities of superoxide dismutase (SOD),peroxidase (POD),catalase (CAT) and ascorbate peroxidase (APX).Exogenous NADP+and NADPH also increased leaf chlorophyll content and root activities.Comprehensive evaluation of the physiological indexes by subordinate function values analysis indicated that spraying 5 μmol·L-1NADPH in the NaCl treatment had the best treatment effect,followed by 10 μmol·L-1NADP+.These results indicated that exogenous NADP+and NADPH can alleviate the damage of NaCl stress on tomato seedlings by reducing membrane lipid peroxidation and increasing leaf chlorophyll mass fraction and antioxidant enzyme activities.
NaCl stress; Processing tomato; NADP+; NADPH
XIN Bingru,female,master,Research area: horticulture and soilless cultivation.E-mail:595170733@qq.com
LIU Huiying,female,professor.Research area: horticulture vegetable physiology and biochemistry.E-mail:hyliuok@aliyun.com
日期:2017-03-03
网络出版地址:http://kns.cnki.net/kcms/detail/61.1220.S.20170303.0834.056.html
2016-02-06
2016-04-06
国家自然科学基金(31360478,31160391);国家星火重点项目(2015GA891008);兵团国际合作项目(2014BC002)。
莘冰茹,女,硕士, 从事设施园艺与无土栽培研究。E-mail:595170733@qq.com
刘慧英,女,教授,从事蔬菜生理生化及设施园艺研究。E-mail:hyliuok@aliyun.com
S641.2
A
1004-1389(2017)03-0420-09
Received 2016-02-06 Returned 2016-04-06
Foundation item The National Natural Science Foundation of China(No.31160391,No.31360478); the Key Project of China Spark(No.2015GA891008);the International Cooperation Project of Xinjiang Production and Construction Corps (No.2014BC002).
猜你喜欢
舰船科学技术(2022年11期)2022-07-15
江苏农业学报(2022年6期)2022-02-06
阅读(科学探秘)(2020年8期)2020-11-06
河南农业科学(2020年2期)2020-03-11
西藏农业科技(2019年3期)2019-11-04
中国果业信息(2019年1期)2019-01-05
现代园艺(2018年3期)2018-02-10
生物学教学(2017年9期)2017-08-20
上海农业学报(2017年3期)2017-04-10
食品工业科技(2014年6期)2014-05-10
