
3种菊科入侵植物白花鬼针草、胜红蓟和假臭草的种子生物学特性比较研究
2017-04-14 09:23钟军弟周宏彬刘锴栋袁长春李先琨
杂草学报 2016年2期
钟军弟 周宏彬 刘锴栋 袁长春 李先琨 刘晚苟
摘要:为探讨菊科入侵植物种子的繁殖特性,从种子的形态特征及种子萌发率等指标入手,对胜红蓟(Ageratumconyzoide)、白花鬼针草(Bidensalba)及假臭草(Eupatoriumcatarium)3种菊科入侵植物的种子特性进行比较研究。结果表明,白花鬼针草种子的冠毛芒最短及最少,沉降速度最大,种子的风媒传播能力最弱,但种子全身具长倒钩刺,主要通过人畜和货物等实现远距离散播。假臭草种子冠毛芒较胜红蓟的长和多,种子沉降速度较胜红蓟的小,其风媒传播能力较胜红蓟的强。白花鬼针草种子千粒质量、发芽指数、胚根、胚芽和胚轴均最大或最长,发芽率和发芽势也较高,其种子质量和种子活力为三者中最高。假臭草的千粒质量、胚根、胚芽和胚轴与胜红蓟无显著差异,但发芽势及发芽指数均显著高于胜红蓟,说明假臭草种子活力较胜红蓟的高。假臭草发芽率为三者中最小,这可能与其种子含水率(19%)高有关,高含水率隐含着种子在潮湿环境下易发生霉变。胜红蓟种子除了发芽率较高外,千粒质量、发芽势、发芽指数、胚根、胚芽和胚轴等指标均较小或大,反映其种子质量和活力较小。
关键词:胜红蓟;假臭草;白花鬼针草;种子生物学特性;种子活力
中图分类号:Q94文献标志码:A文章编号:1003-935X(2016)02-0007-05
菊科(Asteraceae)不仅是我国入侵种最多的科,也是危害性最为严重的科之一[1]。菊科中的胜红蓟(Ageratumconyzoides)、白花鬼针草(Bidensalba)、假臭草(Eupatoriumcatarium)等入侵种对我国的生物多样性、生态系统安全和区域经济发展等已经造成不同程度的危害,这不得不引起我国对菊科入侵植物的更多关注和研究。
胜红蓟别称藿香蓟,菊科藿香蓟属(Ageratum)植物,一年生草本植物,原产中南美洲、墨西哥一带,现广泛分布于我国长江流域及以南地区[1],特别是广东、广西、云南、香港、福建及海南岛等地生长最多。胜红蓟为区域性恶性杂草[2],并具有较强的入侵性,在入侵地常形成优势种群,抑制其他植物的生长,严重威胁当地生物多样性及生态系统。
白花鬼针草,为菊科鬼针草属(Bidens)常见的一年生农业杂草,广泛分布于热带和亚热带区域。在我国,白花鬼针草广泛分布广东省,尤其在粤西地区如湛江、茂名、阳江等地入侵最为严重[3]。鬼针草属植物具有强烈化感作用,常形成大面积单优种群[4],造成当地生物多样性减少,严重威胁着本土植物的生存。
假臭草别称猫腥菊,属菊科泽兰属(Eupatorium)一年生草本植物,原产南美,主要分布于阿根廷、巴西及南美洲其他一些国家,现散布于东半球热带地区。在我国,假臭草主要发生于华南地区,现广泛分布于广东、福建、澳门、香港、台湾、海南等地[5]。假臭草对土壤养分、水分等吸收力强,对土壤的破坏力极大,对农田、人工草地、果园、人工种植林等植被造成较严重的危害,不但在经济上造成损失,还对当地生态系统生物多样性构成巨大的潜在威胁。
因此,这些外来植物的入侵已成为一个越来越严重的问题,它不仅严重威胁着当地的生物多样性,还对生态系统的结构和功能产生了影响。确定这些外来植物的入侵机制,是有效控制其继续入侵的前提。种子特性对植物种群更新和分布区的扩张具有重大的影响[6]。研究表明,许多外来入侵植物都具有较强的有性繁殖能力,其有性繁殖能力的大小与入侵能力呈正相关,并在其成功入侵过程中起关键性作用[7]。目前,国内外学者已经对一些菊科入侵种种子萌发等特性进行了研究[8-10],但比较几种入侵植物种子特性差异以探讨外来植物成功入侵因素方面的研究则较少报道。因此,本研究以外来入侵植物胜红蓟、白花鬼针草、假臭草为研究对象,通过对其种子形态、种子结构特征、种子萌发等特性的比较,了解影响入侵植物的入侵能力差异的种子特性情况,以探讨外来植物成功入侵的主要原因,为有的放矢地采取有效措施对其种群进行防除和控制提供理论依据。
1材料与方法
1.1材料
试验的胜红蓟、白花鬼针草和假臭草种子于2014年12月分别随机采自于广东省湛江地区的全光照下的健康植株。
1.2方法
1.2.13种菊科入侵植物种子外部形态观察
随机选取成熟、饱满、干燥种子各50粒,利用OlympusSZX7体视显微镜观察种子形状、均匀度及种子冠毛情况等,并拍照,利用Image-ProPlus图像分析软件测量种身长、宽、高和冠毛芒长、芒刺长,利用过根系分析系统WINRHIZOPro2009b测得种身体积。
1.2.23种菊科入侵植物种子千粒质量及比重的测定
将采集的种子自然风干后,每种植物随机选取成熟、饱满、干燥的种子各100粒,设置5个重复,用电子天平测量其质量,然后将其平均值乘以10得到千粒质量。最后计算种子比重,种子比重(g/cm3)=种子千粒质量(g)/种子体积(cm3)。
1.2.33种菊科入侵植物种子沉降速度的测定
种子沉降速度的测定方法见文献[11]。沉降速度的计算公式为沉降速度v=h/t,h为种子释放高度(cm),t为沉降时间(s)。
1.2.43种菊科入侵植物种子含水率的测定
每种植物种子随机选取成熟、饱满的种子各100粒,设置5个重复,用电子天平称质量,记录其质量(m1)后,置一干净烧杯中,在75℃烘箱干燥至恒质量,记录其质量(m2),种子含水率=(m1-m2)/m1×100%。
1.2.53种菊科入侵植物种子发芽率、发芽势及发芽指数的测定
试验采用纸皿法[12],即每种植物挑选成熟饱满的种子各50粒,将种子均匀放入底部垫有2层滤纸、直径为12cm并且在121℃高温下灭菌20min的培养皿中进行试验,加入适量蒸餾水后,置于人工气候箱(RXZ型)中培养。培养条件设置为相对湿度70%,温度25℃,光照度1级,光照时间为10h/d。种子萌发统一以胚根露出种皮1mm为准,每天统计发芽的种子数,并且隔天换1次干净的水,直到连续2d无种子萌发为止,然后分别计算发芽率、发芽势和发芽指数。每处理3次重复。
发芽率=(发芽种子数/试验种子数)×100%;
发芽势=前3天发芽种子数/供试种子数×100%。
根据记录的前7d种子发芽情况,代入公式:
发芽指数(GI)=∑(Gt/Dt),式中:Gt指在t时间内的发芽数,Dt指发芽天数。
1.2.63种菊科入侵植物种子萌发后胚芽、胚轴及胚根的测定
每种植物选取已发芽的种子各3颗,借助OlympusSZX7体视显微镜测量种子的体长、胚芽、胚轴、胚根的长度,为3个重复,其中每颗种子的芒长测量4次,芒刺长测量5次,再分别计算其平均值。
1.3统计分析
采用Excel2003和SPSS19.0软件进行数据统计及单因素方差分析(One-wayANOVA),并运用LSD多重比较法对各处理间的差异性进行显著性检验。
2结果与分析
2.13种菊科入侵植物种子形态特征和物理特性的比较
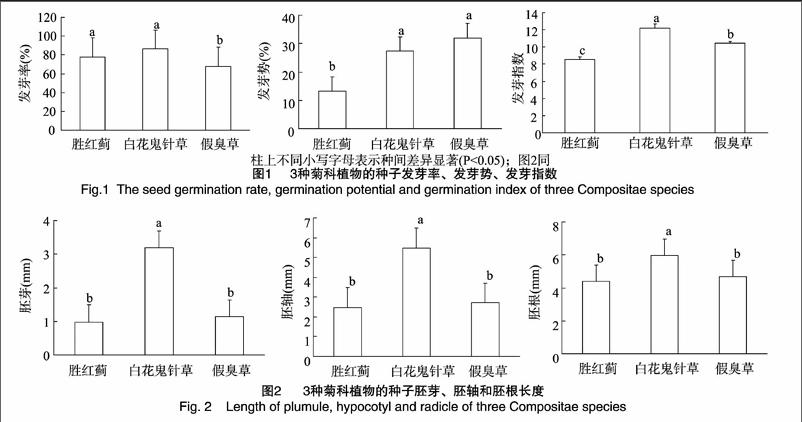
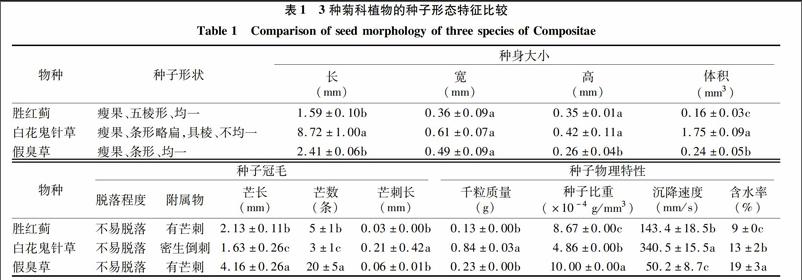
由表1可知,胜红蓟、白花鬼针草和假臭草的果实均为瘦果,5棱形或条形,均具芒及芒刺。白花鬼针草的种子较长[(8.72±1.00)mm],体积[(1.75±0.09)mm3]较大,种子比重[(4.86×10-4±0.00×10-4)g/mm3]较小,种子冠毛芒长度[(1.63±026)mm]较短且数量[(3±1)条]较少,种子沉降速度[(340.5±15.5)mm/s]较大,说明白花鬼针草种子风媒传播能力较弱。白花鬼针草种子全身具密且较长倒钩刺[(0.21±0.42)mm],说明其种子易通过附着人畜和货物等实现远距离散播。假臭草的种子小,体积为[(0.24±0.05)mm3],种子比重虽较大,但其具较长[(4.16±0.26)mm]且数量[(20±5)条]较多的冠毛芒,种子沉降速度[(50.2.4±8.7)mm/s]较小,说明假臭草种子风媒传播能力较强,种子易通过风媒长距离传播。胜红蓟种子比重[(8.67×10-4±0.00×10-4)g/mm3]虽较小,但种子的冠毛芒显著短[(2.13±0.11)mm]且少[(5±1)条]于假臭草,种子沉降速度[(143.4±18.5)mm/s]较假臭草的大,说明胜红蓟种子风媒传播能力远弱于假臭草种子。由此可见,种子冠毛长度和数量等冠毛性状极大地影响着菊科外来入侵种瘦果的沉降速度,影响着种子的传播方式及范围。
白花鬼针草种子千粒质量[(0.08±0.00)g]远大于胜红蓟和假臭草种子,而含水率适中,为(13±2)%,并没有超出国标安全含水率标准(≤15%),说明白花鬼针草种子质量较高。假臭草的种子千粒质量较小,为(0.02±0.00)g,含水率却高达(19±3)%,远超出国标安全含水率,隐含着假臭草种子易发生霉变。胜红蓟种子的千粒质量和含水率均最小,分别为(0.01±0.00)g和(9±0)%。
2.23种菊科入侵植物种子萌发参数的比较
由图1可知,白花鬼针草的发芽率(87±6)%、发芽势(27±3)%和发芽指数(12±1)均较高,说明白花鬼针草的种子活力较高。另外,假臭草的种子发芽势和发芽指数较胜红蓟的高,说明假臭草种子活力较胜红蓟的高。假臭草种子发芽率为(68±3)%,超过50%,但较胜红蓟[(78±3)%]的略小。
2.33种菊科植物种子胚芽、胚轴及胚根长度的比较
由图2可知,发芽后测得的种子胚芽、胚轴与胚根中,白花鬼针草的胚芽、胚轴和胚根均显著长于胜红蓟和假臭草的;胜红蓟的胚芽、胚轴和胚根最短,但与假臭草的无显著差异。说明白花鬼针草种子萌发成幼苗初期的生活能力较假臭草和胜红蓟的强。
3讨论
种子的形态特征不仅是其稳定遗传信息的表达,还与其生长特性有很大的关联。对于适应风力传播的种子而言,沉降速度是衡量植物种子传播能力的重要参数[13-14]。种子沉降速度越大,种子在空气中的漂浮时间越长,种子扩散的距离就越远[15]。种子沉降速度受种子饱满度、成熟度及水分、冠毛面积及张开角度、雷诺数等自身因素影响[14]。但其并不是受简单一因子的影响,而是受众多因子的综合作用的影响[16],其中某些因子可能起着决定性作用。有研究表明,沉降速度与种子上的冠毛数量和长度均呈直接的负相关关系[13-14]。本研究的白花鬼针草种子相对较大,种子比重虽较小,但其冠毛芒数量少且短,种子的沉降速度较大,种子在空中漂浮时间较短,因此,种子风力传播能力相对较弱,但因其全身具有倒钩刺,钩刺较长,所以易于通过附着于人畜和货物等实现远距离散播。相反,假臭草和胜红蓟的风媒传播能力相对较强,原因是假臭草和胜红蓟种子较小,冠毛芒较长、芒数量较多,种子沉降速度较小,种子在空中漂浮时间较长,因而,二者主要通过风媒进行传播,二者较容易优先入侵空生态位资源。假臭草种子的比重虽比胜红蓟的大,但其种子冠毛芒和芒数量远比胜红蓟的长或多,种子沉降速度远比胜红蓟的小,因此,种子风媒传播能力及传播范围较胜红蓟的大。可见,假臭草种群分布较胜红蓟广泛,入侵较胜红蓟为严重。这与国家环境保护总局和中国科学院联合公布的外来入侵种情况相一致的,胜红蓟并没有列入我国3批外来入侵种名单中,而假臭草则列入第3批18种外来入侵种名单中的第7位[17-19]。以上分析可见,在种子重量相对较轻的菊科植物中,种子的冠毛数量和长度等冠毛性状对种子的沉降速度起决定性作用,原因可能是较长和多的种子冠毛增加了種子与空气接触的表面积,从而增加空气对种子的阻力造成的。
种子生活力的表征指标主要有发芽率、发芽势、发芽指数等指标。一般认为,种子发芽率、发芽势和发芽指数高,种子活力也较高[20]。种子胚芽、胚轴和胚根在一定程度上反映幼苗的生活能力,也与种子活力相关[21]。千粒质量也是反映种子品质的重要指标。千粒质量越重,种粒愈大,营养物质较多,种子发芽率高,幼苗生长质量好。植物种子含水率高,种子的呼吸强度增加,种子的温度升高、种子表面易结露甚至发霉[21]。本研究结果表明,在3种入侵植物中,白花鬼针草种子的千粒质量较大,发芽率、发芽势和发芽指数均较高,种子胚芽、胚轴和胚根也均较长,而种子含水率并没有超出国标安全含水率标准,说明白花鬼针草各项种子特性指标良好,种子质量、种子活力及初期幼苗生活能力较高。白花鬼针草为湛江地区的适生种群,为分布范围较广、危害较大的外来入侵杂草之一,这与Linda等的报道[22-23]是相一致的,也与自然界到处可见白花鬼针草单优种群的现象相符合。假臭草种子的千粒质量、发芽势和发芽指数较胜红蓟种子的大,反映出假臭草种子质量和种子活力较胜红蓟的高。假臭草种子发芽率略小于胜红蓟种子发芽率,可能是由高含水率的假臭草种子在萌发培育过程中易发生霉变造成的。假臭草和胜红蓟种子胚芽、胚轴和胚根均较小或较短,说明假臭草和胜红蓟的种子萌发成幼苗初期的生活能力较差,暗示假臭草和胜红蓟的种子质量和活力较白花鬼针草的低,这可能与其种子自身的遗传特性有关。
综上所述,白花鬼针草种子的质量及活力和幼苗初期的生活能力较高,且适宜附着于人畜和货物等实现远距离散播。假臭草和胜红蓟的种子质量和活力及幼苗初期的生活能力相对较差,但其种子发芽率均较高,超过50%,这对种群空间资源的迅速占据是十分有效的。研究可见,应根据各入侵种种子自身特性对其进行布防,如应在各入侵种种子未成熟之前将其植株机械砍掉,阻止植株营养生长向有性生长(种子的发生及成熟)的生活史阶段过渡。另外,还应对通过附着方式进行远距离传播的种子(如白花鬼针草种子)进行严格地域关口检疫把关,防止其进一步远距离传播;多植树造林,减缓风速,减缓通过风媒传播种子(如假臭草和胜红蓟种子)的远距离传播。
参考文献:
[1]徐海根,强胜.中国外来入侵物种编目[M].北京:中国环境科学出版社,2004.
[2]强胜,曹学章.中国异域杂草的考察与分析[J].植物资源与环境学报,2000,9(4):34-38.
[3]陈志云,梁水凤,李东文,等.假臭草等12种植物对白花鬼针草幼苗的化感作用[J].热带亚热带植物学报,2011,19(5):454-462.
[4]杜凤移,张苗苗,马丹炜,等.三叶鬼针草化感作用的初步研究[J].中国植保导报,2007,27(9):8-11.
[5]吴海荣,胡学难,钟国强,等.外来杂草假臭草的特征特性[J].杂草科学,2010(5):174-175.
[6]刘路芳,马绍宾.滇大蓟种子特性和影响萌发因素研究[J].种子,2005,24(12):57-59.
[7]徐汝梅,叶万辉.生物入侵——理论实践[M].北京:科学出版社,2003.
[8]阚丽艳,谢贵水,安锋,等.海南省入侵植物假臭草种子萌芽分析及其防治对策[J].广西农业科学,2008,39(1):46-50.
[9]ChauhanBS,JohnsonDE.GerminationecologyoftwotroublesomeAsteraceaespeciesofrainfedrice:siamweed(Chromolaenaodorata)andcoatbuttons(Tridaxprocumbens)[J].WeedScience,2008,56:567-573.
[10]洪岚,沈浩,杨期和,等.外来入侵植物三叶鬼针草种子萌发与贮藏特性研究[J].武汉植物学研究,2004,22(5):433-437.
[11]潘燕,王帅,王崇云,等.云南松与云南油杉种子风力传播特征比较[J].植物分类与资源学报,2014,36(3):403-410.
[12]曾任森.化感作用研究中的生物测定方法综述[J].应用生态学报,1999,10(1):123-126
[13]AndersonMC.Ananalysisofvariabilityinseedsettlingvelocitiesofseveralwind-dispersedAsteraceae[J].AmericanJournalofBotany,1992,79:1087-1091
[14]AndersonMC.Diasporemorphologyandseeddispersalinseveralwind-dispersedAsteraceae[J].AmericanJournalofBotany,1993,80:487-492
[15]SchulzB,JoachimD,GerhardG.Apparatusformeasuringthefallvelocityofanemochorousdiasporeswithresultsfromtwoplantcommunities[J].Oecologia,1991,86:454-456
[16]郝建华.部分菊科入侵种的有性繁殖特征与入侵性的关系[D].南京:南京农业大学,2008.
[17]国家环保总局,中国科学院.中国第一批外來入侵物种名单[J].中华人民共和国国务院公报,2003(23):41-46.
[18]国家环保总局,中国科学院.中国第二批外来入侵物种名单[J].中华人民共和国国务院公报,2010.
[19]国家环保总局,中国科学院.中国第三批外来入侵物种名单[J].中华人民共和国国务院公报,2014.
[20]许慧男,王文杰,于兴洋,等.菊科几种入侵和非入侵植物种子需光发芽特性差异[J].生态学报,2010,30(13):3433-3440.
[21]韩利红,刘潮,郑玉龙.紫茎泽兰与3种同属本地植物种子特性比较[J].种子,2010,29(2):73-76.
[22]LindaCD.Bidensalba[EB/OL].(2009-12-16)[2016-02-01].http://www.floridata.com/ref/B/bide_alb.cfm.
[23]田兴山,岳茂峰,冯莉,等.外来入侵杂草白花鬼针草的特征特性[J].江苏农业科学,2010(5):174-175.
