
山地木麻黄种源在海南临高的遗传变异及选择
2017-08-07 10:18仲崇禄姜清彬KhongsakPinyopusarerk
林业科学研究 2017年4期
韩 强,仲崇禄*,张 勇,姜清彬,陈 羽, 陈 珍,Khongsak Pinyopusarerk
(1. 中国林业科学研究院热带林业研究所,广东 广州 510520;2. 澳大利亚联邦科学与工业研究组织林木种子中心, ACT 2601,堪培拉,澳大利亚)
山地木麻黄种源在海南临高的遗传变异及选择
韩 强1,仲崇禄1*,张 勇1,姜清彬1,陈 羽1, 陈 珍1,Khongsak Pinyopusarerk2
(1. 中国林业科学研究院热带林业研究所,广东 广州 510520;2. 澳大利亚联邦科学与工业研究组织林木种子中心, ACT 2601,堪培拉,澳大利亚)
[目的]研究山地木麻黄种源间抗风性、生长及形质性状的遗传变异规律,为山地木麻黄的良种选育和种质资源的合理开发利用提供科学依据。[方法]以27个山地木麻黄种源为试验材料,于造林后2、5、7 a时测定山地木麻黄种源的树高、胸径、单株材积和保存率等数量性状,并于造林后7 a时调查主干分叉习性(AP)、主干通直度(SFS)、侧枝密度(DPB)、侧枝直径(TPB)、绿色小枝长度(LDB)、侧枝分枝角(APB)、侧枝长度(LPB)等形质性状以及抗风性(RES),通过方差分析、相关性分析及遗传参数估算揭示其遗传变异规律。应用坐标综合评定法对山地木麻黄种源进行综合评定。[结果]表明:造林后2、5、7 a时,27个山地木麻黄种源间保存率和抗风性差异显著(P<0.05);对造林后7 a时保存率较高的18个种源进一步分析显示,上述3个年份各种源间树高、胸径和单株材积等生长性状均存在极显著差异 (P<0.01);7 a时,TPB、APB、LDB、AP和SFS等形质性状在种源间亦存在显著或极显著差异;生长性状的种源遗传力明显高于形质性状,二者分别受中度或中度偏下和低度遗传控制;随着林龄的增长,树高的遗传变异系数变化不大,而胸径和单株材积的遗传变异系数呈先增加后降低的趋势,树高、胸径、单株材积的遗传变异系数分别为11.89%~12.30%、11.67%~13.67%、30.20%~38.11%;7 a时,形质性状的遗传变异系数为3.84×10-5%~5.56%。性状间相关分析表明:树高作为山地木麻黄早期选择性状较适宜。[结论]依据坐标综合评定法,筛选出17877、19489和19490等3个优良种源,可在生产上大面积推广。
山地木麻黄;种源选择;遗传力;遗传变异
木麻黄(Casuarinaspp.)具有速生、防风、固沙、抗逆及耐瘠薄等优良特性,是重要的防护林、用材林和多用途林树种[1-2]。我国引种木麻黄有110多年历史,最早于1897年引入台湾[3],20世纪80年代中期,借助国际合作项目,我国开始系统研究木麻黄种质资源的引种和选育[4]。目前,我国引进的木麻黄有20多种,人工种植面积达30多万公顷,但主要为短枝木麻黄(C.equisetifoliaL. ) 、细枝木麻黄(C.cunninghamianaMiq.)和粗枝木麻黄(C.glaucaSieber. ex Spr.)。由于造林树种单一,种质资源匮乏,且长期受自然灾害及病虫害的影响,严重制约了木麻黄人工林的可持续经营[5]。开展山地木麻黄(C.junghuhnianaMiq.)种质资源引种与测试,可丰富木麻黄种质资源,为木麻黄新品种创制和选育等提供基础材料,从而有利于提高沿海防护林的稳定性。
山地木麻黄原产印度尼西亚,具有固氮、速生、抗旱及耐水湿等特性,天然分布于山地及沿海地区,已被广泛引种到世界热带及亚热带地区。国外对山地木麻黄的研究主要集中在种源间生长表现和适应性比较以及优良种源选择[6-8],对其种源遗传变异规律及遗传参数估算研究较少[9]。我国引种山地木麻黄始于20世纪80年代,相继在沿海各地开展小规模引种试验。1996年以来,中国林科院热带林业研究所在广东湛江和福建漳州开展了国际山地木麻黄种源试验,对其遗传参数进行估算,并筛选出适合华南地区生长的优良种源[10-11];福建省林科院在福建漳州开展了滨海沙地山地木麻黄种源试验[12-13];但海南省尚无系统的山地木麻黄种源试验报道,特别是针对大量原产地种质材料的测试。本文以从澳大利亚引进的27个山地木麻黄种源为材料开展种源试验,分析其生长、形质等性状方面的变异状况,揭示种源间性状的遗传变异规律,同时应用坐标综合评定法对各种源间生长和形质性状进行综合评价,以期为山地木麻黄的良种选育和种质资源的合理利用提供参考。
1 试验地概况
试验地位于海南省西北部临高县临城镇林木良种场(19°91′N, 109°69′E),平均海拔5 m,属热带季风气候,高温多雨,光照充足。年均气温23.5℃,1月份平均气温为16.9℃,7月份平均气温为28.3℃。年平均雨日为139.5 d,年均降水量1 417.8 mm。土壤为砖红壤。试验地前茬为桉树。
2 材料与方法
2.1 试验材料与试验设计
试验所用山地木麻黄27个种源种子均由澳大利亚林木种子中心提供(表1)。2008年3月开始育苗,9月造林,采用机耕全垦方式整地,株行距2 m×2 m,造林时每株施150 g复合肥作为基肥。
试验采用随机区组设计,以种源为处理,8次重复,每个种源采用单行6株小区。试验地四周用3行木麻黄无性系A8作为保护行。至2013年调查时,试验林未受任何人为和自然灾害的破坏,总体保存率达80%以上。由于2014年经历2次台风袭击(即威尔逊台风,14级,2014年7月18号登陆;海鸥台风,13级,2014年9月16号登陆),部分种源保存率较低。

表1 27个山地木麻黄种源信息
注:Ind.是印度尼西亚,Ken.是肯尼亚,Tanz.坦桑尼亚。
Note: Ind. is Indonesia, Ken. Is Kenyan. Tanz. is Tanzania.
2.2 试验观测
造林后2、5、7 a时,对所有种源进行每木调查,包括树高(H, m)、胸径(DBH, cm)和保存率(SUR, %)。抗风性观测在台风危害后进行。造林7 a时,对主干通直度(SFS)、主干分叉习性(AP)、侧枝直径(TPB)、侧枝密度(DPB)、侧枝长度(LPB)、侧枝分枝角(APB)、绿色小枝长度(LDB)、抗风性(RES)进行观测分级[14-16]
2.3 数据统计分析
山地木麻黄各性状以单株测定数据进行统计分析,单株材积 (V, m3·株)计算公式为[17]:
V=3.141 592 6×DBH2×H/120 000
在方差分析前,对保存率进行反正弦转换,形质性状数据进行平方根转换。利用R语言结合软件ASReml 3.0和SPSS 22.0进行方差分析与多重比较、相关分析及遗传参数估算[18]。
观测值线性模型为:
yijk=μ+Ri+Pj+RPij+ηijk
式中:yijk为观测值;μ为总体平均值;Ri为第i个区组的固定效应;Pj为第j个种源的随机效应;RPij为第i个区组与第j个种源互作的随机效应;ηijk为个体机误。

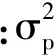
遗传变异系数(GCVA, %)为:

性状间的遗传相关系数(rG)和表型相关系数(rP)为:

种源选择遗传增益估算:
基于各性状的观测值,采用坐标综合评定法[21]对各种源进行综合评价,筛选优良种源。
式中:Pi为综合值, Xij为第i个种源第j个性状的平均观测值,Xjmax为第j个性状的最大平均观测值,n 为性状个数。
3 结果与分析
3.1 山地木麻黄保存率和抗风性
表2表明:在测定的3个年份,山地木麻黄各种源间保存率均差异显著(P<0.05);造林后2、5、7a,其保存率分别为19.67%~99.41%、0.59%~93.93%、0.28%~76.67%,其中,造林后2、5a时,山地木麻黄的整体平均保存率在74%以上,7a时因遭受超强台风袭击,整体保存率降至40%。造林后2、5a时,种源18948、19489和17878保存率最高,均达90%以上;造林后7a时,种源18844、18846、18849和19489保存率最高,均在70%以上。
表2还表明:造林后7a时,山地木麻黄种源抗风性差异显著,种源18952的抗风性最差,种源18849、18845、18954、18846、18852、19239和18853的抗风性相对较好。
由于超强台风的影响,部分种源保存率较低,树干倒伏、折断较严重,小区数据缺失,为保证分析结果的可靠性,基于上述保存率及抗风性分析,剔除7a时缺失5个小区以上的9个种源(18952、18948、18951、18949、18847、18848、18851、19491和18953),仅对保存有5个小区以上的18个种源的生长和形质性状进行统计分析。
3.2 山地木麻黄生长性状与形质性状的变化
从表3可看出:造林后2、5、7a时,18个种源的树高、胸径、单株材积均存在极显著差异 (P<0.01),说明生长性状在种源水平上具有很大的遗传改良潜力;各种源的树高均值分别为3.48~6.12、4.98~8.26、7.50~11.67m,胸径均值分别为2.66~4.75、4.84~7.73、5.96~9.55cm,单株材积均值分别为1.02×10-3~4.46×10-3、4.43×10-3~16.14×10-3、10.20×10-3~34.88×10-3m3,其中,种源19490和17877表现最好,种源18846和18845表现最差。
表4表明:造林后7a时,山地木麻黄侧枝直径(TPB)、侧枝分枝角(APB)、绿色小枝长度(LDB)及主干通直度(SFS)在种源间差异极显著;主干分叉习性(AP)在种源间差异显著;而抗风性(RES)、侧枝密度(DPB)及侧枝长度(LPB)在种源间差异不显著。
3.3 山地木麻黄种源遗传参数估算和相关分析
由表5可知:树高、胸径、单株材积等生长性状的种源遗传力随着林龄的增长,呈先增长后降低的趋势,其种源遗传力分别为13.80%~16.00%、8.64%~11.90%、8.42%~14.30%。造林后7a时,各形质性状间种源遗传力差异较大,TPB、APB、LDB、SFS的种源遗传力均>5%,RES、DPB、LPB、AP的种源遗传力均<1%。树高的遗传变异系数随着林龄的增加变化不大,其遗传变异系数为12%左右;胸径和单株材积的遗传变异系数随林龄的增加呈先增加后降低的趋势,其遗传变异系数分别为11.67%~13.67%、30.20%~38.11%。造林后7a时,除DPB的遗传变异系数接近于0,其他形质性状的遗传变异系数为0.34%~5.56%。采用20%的入选率,造林后2、5、7 a时,树高、胸径、单株材积的遗传增益分别为2.59%~3.08%、1.95%~2.69%、4.86%~9.08%。造林后7 a时,形质性状的遗传增益为3.29×10-8%~2.45%,明显低于生长性状,选择生长性状进行遗传改良效果较好。

表2 27个山地木麻黄种源的保存率和抗风性
注:*P<0.05的显著水平,**P<0.01的极显著水平,ns为不显著,小写字母表示在0.05水平上显著,下同。
Note:*P<0.05 level of significance, **P<0.01 level of significance, ns-no significant, lowercase letter indicate significant at 0.05 level, The same below.

表3 造林后2、5、7 a时山地木麻黄18个种源树高、胸径及单株材积的均值及其多重比较结果

表4 造林后7 a时18个种源山地木麻黄形质性状的得分均值及其多重比较结果

表5 山地木麻黄各性状的方差分量(种源、互作、表型和误差)、种源遗传力、遗传变异系数及遗传增益
注:表中a 表示方差分量×10-3, b 表示方差分量×10-6;H2、DBH2、V2、H5、DBH5、V5、H7、DBH7、V7分别表示造林后2、5、7 a时的树高、胸径和单株材积;RES、DPB、TPB、APB、LPB、LDB、AP、SFS为造林后7 a时测定;下同
Note: a represents variances×10-3, brepresents variances×10-6.H2,DBH2,V2,H5,DBH5,V5,H7,DBH7,V7 represents the height, diameter at breast height and volume at 2, 5 and 7 years after planting, respectively.RES,DPB,TPB,APB,LPB,LDB,AP,SFSis measured at 7 years after planting. The same below.
由表6可看出:树高、胸径、单株材积3个生长性状间的表型和遗传均呈极显著相关,相关系数均>0.68。TPB与3个生长性状间的遗传和表型均呈显著或极显著负相关,而APB与3个生长性状间的遗传和表型均呈显著或极显著正相关;APB除与TPB间的表型和遗传呈极显著负相关外,与其他形质性状均相关不显著;树高与SFS间的表型和遗传呈极显著正相关,这对选择生长快和干形好的种源极为有利。不同林龄间,树高和胸径的遗传相关系数均在0.78以上,达极显著相关,说明对生长性状进行早期选择是可行的。

表6 山地木麻黄各性状的遗传相关系数(上三角),表型相关系数(下三角)
3.4 综合评价与优良种源的选择
选择种源间差异显著的生长及形质性状,应用坐标综合评定法对18个种源进行综合评定(表7)发现,最好的3个种源为17877、19489和19490,其评估得分分别为0.074 5,0.095 4和0.212 2;种源19242最差,其评估得分值为1.277 0。
4 讨论
目前,在山地木麻黄的遗传改良过程中,种源选择是遗传改良的常规方法,探究种源的遗传变异规律,可为杂交育种亲本选配提供依据。泰国和印度的山地木麻黄种源试验结果表明,不同种源在生长表现、木材密度和含水量、树皮厚度、干形以及分枝特性等方面存在显著差异[7-8,22]。我国相继开展了多地种源试验,仲崇禄等[10]在福建漳州和广东电白的研究表明,4年生时,树高、胸径和单株材积在种源间存在显著或极显著差异。本研究结果显示,造林后7 a时,山地木麻黄的生长性状(树高、胸径、单株材积)和部分形质性状(侧枝直径、侧枝分枝角、绿色小枝长度、主干通直度及主干分叉习性)在种源水平上亦存在显著或极显著差异,与上述结论相似,说明开展山地木麻黄种源选择具有较大潜力。
山地木麻黄生长性状的各遗传参数在生长过程中呈现出较强的规律性。树高、胸径和单株材积的种源遗传力随林龄的增长均呈先增加后降低的趋势,造林后7 a时种源遗传力下降,很可能是由于2014年超强台风的袭击导致环境方差分量增大造成的。仲崇禄等[10]开展山地木麻黄多地点种源试验,其4年生时生长性状的种源遗传力比本研究造林后2、5、7 a时都低,可能是由于本研究基于单一地点试验的缘故[23];就单一地点而言,本研究显示海南临高山地木麻黄生长性状的种源遗传力比广东电白的低[11],说明地点对山地木麻黄种源试验的影响较大,因此,开展多地点种源试验,有利于提高遗传参数估算的可靠性。Robinson[24]对遗传力的程度进行了分类,受高度、中度和低度遗传控制时,其遗传力分别为30%~ 60%、10 %~30%、5%~10%。山地木麻黄生长性状种源遗传力随林龄的波动不大,其变化范围为8.42 %~16.00%,说明其受中度或中度偏下的遗传控制。造林后7 a时,在观测的多个性状中,生长性状的遗传变异系数明显大于形质性状,而生长性状中树高的遗传变异系数高于胸径,说明树高在种源间变异更显著,而且其与大部分形质性状显著相关,因此,树高是山地木麻黄种源选择最有效的性状。
林木生长周期较长,良种选育难度较大,为缩短育种周期,早期选择作为筛选优良种源的有效途径颇受重视。国内研究认为,木麻黄速生用材林的轮伐期为7~15 a[25];泰国研究认为,用于速生用材林和薪炭林的山地木麻黄轮伐期为5 a[26]。本文选用造林后2、5、7a 3个林龄的生长性状进行早晚相关分析发现,山地木麻黄的树高和胸径早-晚相关十分紧密,且主要受遗传因素控制。比较树高和胸径的种源遗传力及早-晚相关系数,发现树高的种源遗传力和早-晚相关系数比胸径相对应的值大,因此,用树高作为山地木麻黄早期选择性状比较适宜。
基于山地木麻黄的生长及形质性状进行遗传参数估算,本研究初步筛选出一批优良种源,为山地木麻黄优良单株的选择奠定了基础。此外,还应对山地木麻黄的木材材性进行研究,开展生长和材性性状的遗传相关分析,将有助于速生优质山地木麻黄新品种选育,亦为将来木麻黄种内和种间杂交育种提供亲本材料。
5 结论
(1)造林后2、5、7 a时,参试的27个山地木麻黄种源间保存率和抗风性均差异显著(P<0.05);而造林后7 a时保存率较高的18个种源间树高、胸径和单株材积等生长性状均存在极显著差异(P<0.01);7 a时,TPB、APB、LDB、AP和SFS等形质性状在种源间亦存在显著或极显著差异。
(2)山地木麻黄的树高、胸径及单株材积等生长性状受中等或中等偏下的遗传控制,而其形质性状受低程度遗传控制。
(3)各性状的遗传变异系数及性状间相关性分析表明,树高适宜作为山地木麻黄早期选择性状。
(4)通过各性状综合评定,筛选出17877、19489和19490等3个优良种源。
[1] Midgley S J, Turnbul L, Johnston R D. Casuarina Ecology [M]. Canberra: CSIRO, Management and Utilization, 1983.
[2] Sougoufara B, Diem H G, Dommergues Y R. Response of field-grownCasuarinaequisetifoliato inoculation with Frankia strain ORS 021001 entrapped in alginate beads [J]. Plant and Soil, 1989, 118(1):133-137.
[3] 杨政川,张添荣,陈财辉,等.木贼木麻黄在台湾之种源试验Ⅰ.种子重与苗木生长[J].林业试验研究报告,1995,10(2):2-7.
[4] 仲崇禄,白嘉雨,张 勇.我国木麻黄种质资源引种与保存[J].林业科学研究,2005,18(3):345-350.
[5] 张 勇.三种木麻黄的遗传改良研究[D].北京:中国林业科学研究院,2013.
[6] Mwihomeke S T, Mugasha A G, Chamshama S A O,etal. Early performance of provenances/land races at Lushoto, Tanzania [J]. Southern African Forestry Journal, 2002, 194(1):7-14.
[7] Luechanimitchit P. Early results of a progeny trial ofCasuarinajunghuhnianainThailand[J]. NFT News, 2006, 9(1):1-2.
[8] Nicodemus A, Varghese M, Nagarajan B. Selection ofCasuarinajunghuhnianaMiq. provenances for multiple end uses in India[C]// Tewari V P, Srivastava R L. ‘ Multipurpose trees in the tropics: management and improvement strategies ’ . Proceedings of the IUFRO international conference, 22-25 November 2004, Jodhpur, India,2005:650-657.
[9] Varghese M, Nicodemus A,Ravi N. Genetics and ecological variation in wood properties ofCasuarinaspecies[C]// Gurumurthi K, Nicodemus A,Siddappa.CasuarinaImprovement and Utilization. IFGTB, Institute of Forest Genetics and Tree Breeding, Coimbatore, India,2001:29-38.
[10] 仲崇禄,施纯淦,王维辉,等.华南地区山地木麻黄种源试验与筛选[J].林业科学,2002,38(6):58-65.
[11] 林什全,仲崇禄,白嘉雨.广东省电白县5年生山地木麻黄种源试验及评选[J].林业科学研究,2003,16(4):506-510.
[12] 谭芳林.福建滨海沙地山地木麻黄种源试验与选择研究[J].防护林科技,2009(2):1-2.
[13] 施成坤.山地木麻黄种源选择研究[J] 安徽农学通报,2009,15(3):141-142.
[14] Pinyopusarerk K, Kalinganire A, Williams E R,etal. Evaluation of international provenance trials ofCasuarinaequisetifolia.[M]. Canberra: CSIRO, Technical Reports, 2004.
[15] 仲崇禄,白嘉雨.山地木麻黄家系遗传参数估算与家系选择[J].林业科学研究,1998,11(4):361-369.
[16] 许秀玉,王明怀,仲崇禄,等.不同树种木材性质及其抗台风性能[J].浙江农林大学学报,2014,31(5):751-757.
[17] 仲崇禄.木麻黄遗传变异规律的研究[D].北京:中国林业科学研究院,2000.
[18] 林元震.R与ASReml-R统计分析教程[M]. 北京:中国林业出版社,2014.
[19] Gapare W J, Musokonyi C, Musokonyi C. Provenance performance and genetic parameter estimates forPinuscaribaeavar.hondurensisplanted at three sites in Zimbabwe [J]. Forest Genetics, 2002, 9(1):183-189.
[20] Xi yang Zhao, Cheng Wang, Shuchun Li,etal. Genetic variation and selection of introduced provenances of Siberian Pine (Pinussibirica) in frigid regions of the Greater Xing'an Range, Northeast China [J]. Journal of Forestry Research, 2014, 25(3):549-556.
[21] 刘 宇,郭建斌,邓秀秀,等.秦岭火地塘林区3种土地利用类型的土壤潜在水源涵养功能评价[J].北京林业大学学报,2016,38(3):73-80.
[22] Jayaraj R S C.Casuarinajunghuhniana(Casuarinaceae) in India [J]. Australian Journal of Botany, 2010, 58(2): 149-156.
[23] White T L, Adams W T, Neale D B.森林遗传学[M].崔建国,李火根,译. 北京:科学出版社,2013:101-130.
[24] Robinson H F, Comstock R E, Harvey P H. Estimates of Heritability and the Degree of Dominance in Corn [J]. Agronomy Journal, 1949, 41(8):353-359.
[25] 徐燕千,劳家骐.木麻黄栽培[M].北京:中国林业出版社,1984.
[26] Orwa C, A Mutua, Kindt R,etal. 2009 Agroforestree Database: a tree reference and selection guide version 4.0 (http://www.worldagroforestry.org/sites/treedbs/treedatabases.asp)
(责任编辑:徐玉秀)
Genetic Variation and Selection ofCasuarinajunghuhnianaProvenances at Lingao, Hainan
HANQiang1,ZHONGChong-lu1,ZHANGYong1,JIANGQing-bin1,CHENYu1,CHENZhen1,KhongsakPinyopusarerk2
(1. Research Institute of Tropical Forestry, Chinese Academy of Forestry. Guangzhou 510520, Guangdong, China. 2. CSIRO Australian Tree Seed Centre, GPO Box 1600, ATC 2601, Canberra, Australia)
[Objective]In order to screen out superior provenances and individual trees for further cross-breeding, the inter-provenance genetic variations in terms of wind-resistance, growth and morphological traits were discussed in this context due to a trial ofCasuarinajunghuhnianaincluding 27 provenances at Lingao, Hainan, China. [Method] The height, DBH, volume and survival were measured at the 2nd, 5th and 7th years after planting, and the qualitative traits including AP, SFS, DPB, TPB, LDB, APB, LPB and RES were investigated at the 7th year after planting. The genetic variations of these traits were studied by variance analysis, genotypic and phenotypic correlations, and genetic parameters. The provenances ofC.junghuhnianawere also assessed by comprehensive coordinate method. [Result] Significant differences (P<0.05) were detected among 27 provenances in survival and RES at the 2nd, 5th and 7th years after planting. For a further analysis, the top 18 provenances in higher survivals at age of 7 indicated signi?cant differences (P<0.01) in tree height, DBH, volumes among provenances at the 2nd, 5th and 7th years after planting. It was also found that there were significant differences in TPB, APB, LDB, SFS (P<0.01) and AP (P<0.05) among provenances 7 years after planting. The heritability was moderate for growth traits and low for qualitative traits, suggesting the genetic controls were moderate on growth and weak on qualitative traits. Trends of coefficient of genetic variation for height were nearly stable, while the coefficients of genetic variations in DBH and volume tended to decreasing at initial and then increasing with age. The coefficients of genetic variation of height, DBH and volume were 11.89%-12.30%, 11.67%-13.67% and 30.20%-38.11%, respectively, and those for qualitative traits 7 years after planting ranged from 3.84×10-5% to 5.56%. The high age-age and trait-trait phenotypic and genetic correlations for height and DBH indicated that early selection and multiple traits selection were feasible. Height was superior to other traits in the early selection forC.junghuhnianadue to its genetic stability. [Conclusion] By analyzing nine traits at the 7th year after planting by comprehensive coordinate method, three optimal provenances were selected, which are worthy to be developed for producing and cross-breeding.
Casuarinajunghuhniana; provenance selection; heritability; genetic variations
10.13275/j.cnki.lykxyj.2017.04.009
2016-11-18
广东省林业科技创新项目(2014KJCX017);国家自然科学基金项目(31470634);“十二·五”科技支撑项目(2012BAD01B0603)
韩强(1988—),男,在读博士研究生,主要从事木麻黄遗传改良研究.E-mail:qizhouhanqiang@sina.com
* 通讯作者:仲崇禄,研究员,主要研究方向为林木遗传改良.E-mail:zcl@ritf.ac.cn
S792.93
A
1001-1498(2017)04-0595-09
猜你喜欢
世界热带农业信息(2022年11期)2022-10-18
现代园艺(2021年8期)2021-05-13
吉林林业科技(2020年6期)2020-12-16
中学生博览(2020年23期)2020-12-10
文苑·经典美文(2019年9期)2019-09-09
绿色科技(2019年13期)2019-08-31
天然产物研究与开发(2018年3期)2018-05-07
闽南风(2017年12期)2018-01-04
数学教学通讯·小学版(2016年4期)2016-05-27
中国篆刻·书画教育(2016年2期)2016-03-07
