
干旱胁迫下接种ERM真菌对桃叶杜鹃幼苗光合特性的影响
2017-12-01 10:11,,,,
种子 2017年6期
, , , ,
(1.贵州大学林学院, 贵阳 550025; 2.贵州省生物研究所, 贵阳 550009)
干旱胁迫下接种ERM真菌对桃叶杜鹃幼苗光合特性的影响
龙海燕1,欧静1,谌端玉1,陈荣建1,张玉武2
(1.贵州大学林学院, 贵阳 550025; 2.贵州省生物研究所, 贵阳 550009)
以1年生桃叶杜鹃幼苗为材料,研究干旱胁迫下接种ERM真菌对桃叶杜鹃幼苗光合特性的影响。结果表明:接种ERM真菌显著提高了桃叶杜鹃幼苗的叶绿素含量、净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)以及水分利用效率(WUE),同时降低了叶片胞间CO2浓度(Ci),对照不接菌处理差异显著;在持续干旱条件下,桃叶杜鹃菌根苗通过促进叶片叶绿素的合成与积累,提高水分的利用效率,以减缓幼苗各光合参数的下降速度,使幼苗保持较高的光合能力;复水后,菌根苗能较快恢复其光合作用,恢复能力显著高于对照苗,且TY 35和TY 29基本能恢复至旱前水平。由此可见,接种ERM真菌能够显著提高桃叶杜鹃幼苗的光合能力以及水分利用效率,从而增强桃叶杜鹃的抗旱能力,且以TY 35和TY 29的表现最好。
杜鹃花类菌根; 桃叶杜鹃; 叶绿素含量; 气体交换参数; 干旱胁迫
杜鹃花类菌根(Ericoid mycorrhizal,简称ERM)又称拟欧石楠类菌根,是大多数杜鹃花科植物根系与一些土壤真菌形成的具有典型特征的共生体[1]。已有研究表明,ERM对杜鹃花类植物的营养吸收以及增强对逆境因子的抗性等方面具有重要作用[2],是杜鹃花科植物在恶劣的生境条件下广泛分布的关键原因。国内对于ERM的研究起步较晚,研究主要集中在杜鹃花类菌根的结构[3]、真菌的分离和鉴定[4-5]、菌根真菌纯培养以及接种效应[6]等方面,关于ERM提高宿主植物抗旱性方面的研究鲜见报道。
桃叶杜鹃(RhododendronannaeFranch.)是高山常绿杜鹃代表种,仅分布于贵州和云南的高山地区,花色艳丽,是一种观赏价值较高的晚花类杜鹃[7],具有极高的园林应用前景,但其受遗传因素的制约,喜冷凉、湿润环境,根系为浅根系,不能伸入土层深处,抗旱性弱,制约了桃叶杜鹃引种栽培与园林应用。欧静等研究发现,接种ERM真菌的桃叶杜鹃其幼苗发芽率及移栽成活率均显著提高[8];接种后,桃叶杜鹃幼苗的苗高、地径和生物量等生长指标也明显提高[9]。ERM真菌对桃叶杜鹃具有较好的促生效应,但关于ERM提高桃叶杜鹃抗旱性的研究未见报道。
本试验以1年生桃叶杜鹃苗为试验材料,通过测定干旱胁迫下接种不同ERM真菌桃叶杜鹃幼苗的叶绿素含量、气体交换参数以及水分利用效率等指标,分析接种ERM真菌对桃叶杜鹃幼苗光合特性的影响,以期为桃叶杜鹃的引种栽培及园林应用提供参考。
1 材料与方法
1.1 试验材料
1.1.1 试验地
试验在贵州大学南校区林学院苗圃内进行。
1.1.2 供试菌株
4个试验菌株分离自野生桃叶杜鹃根系中,编码分别是TY 02(AJ 279484:Ascomycetesp.),TY 21(AB 369417:Cryptosporiopsisericae),TY 29(AB 158314:Trametesochracea),TY 35(AB 378554:Pochoniabulbill-osa)(括号中前者为国际GenBank中的登录号,后者为与试验菌株亲缘关系最近种)[9],保存于4 ℃冰箱。采用PDA培养基对试验菌株进行平板培养,接种后放入25 ℃暗箱内培养15 d左右,待基质长满菌丝后备用。
1.1.3 试验苗木及基质
试验苗为通过播种繁殖培养的1年生桃叶杜鹃苗。桃叶杜鹃种子、土壤均采自贵州省毕节市百里杜鹃风景区,试验基质是由腐殖土和石英砂按3∶1比例混合。土壤理化性质:pH=4.75、有机质44.10 g/kg、全氮1.54 g/kg、全磷0.163 g/kg、碱解氮268.52 mg/kg、有效磷7.00 mg/kg、速效钾205.96 mg/kg。
1.2 试验方法
1.2.1 接种处理
试验设5个处理,分别是接种菌株TY 02、TY 21、TY 29、TY 35和不接种作为对照ck。在121 ℃的高压灭菌锅中将基质土壤连续灭菌2 h。塑料盆和饱满的桃叶杜鹃种子在0.5% KMnO4溶液中浸泡10 min进行消毒,用蒸馏水冲洗干净。接种前在规格为高210 mm×直径160 mm的塑料盆内装入2/3的灭菌土,接种时将培养好的菌株平铺于灭菌土上,每盆接种1个培养皿的菌株,之后再铺上1层灭菌土,播入灭菌种子。由于桃叶杜鹃种子极小,播种方法采用撒播,100粒/盆,每处理15盆,轻轻盖上剪碎并灭菌后的马尾松针叶,处理结束后放入人工气候培养箱(白天、夜间都为25 ℃,湿度为90%、光照12 h)培养。
1.2.2 持续干旱及复水处理
待幼苗出土生长90 d后,将其移出人工气候培养箱,移栽到贵州大学林学院苗圃大棚内进行菌根苗培育。于2015年8月选取各处理生长一致的1年生桃叶杜鹃幼苗,每处理10盆,每盆1株,各处理重复3次,共150盆,统一浇水管理30 d后,使每盆的土壤含水量均达到饱和状态,于2015年9月开展持续干旱胁迫,置于温室大棚内模拟自然干旱,使水分不断消耗。分别在持续干旱当天(0)、第5天、第10天和第15天对相关指标进行测定,同时在干旱胁迫的第15天对幼苗进行复水,并在复水5 d后进行取样调查,测量相关光合指标。
1.2.3 指标测定
1) 叶绿素含量测定。参照邹琦[10]的方法进行测定,每处理重复3次。
2) 光合特征参数的测定。使用Li-6400(Li-Cor.,美国)便携式光合测定仪对桃叶杜鹃菌根苗叶片的净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、气孔导度(Gs)等光合参数进行测量,测定时叶室配备LED红蓝光源, 光量子通量密度(PPFD)为1 000μmol/(m·s),CO2浓度为400μmol/mol,温度为27 ℃。测定1次,每片叶子待系统稳定后记录3个瞬时值。测定时选取各处理幼苗受光方向相同、无病虫害、叶面积相近且生长一致的第4片叶,每处理重复测量5株,每株固定3片。利用公式 WUE=Pn/Tr计算叶片水分利用效率。
1.3 数据处理
试验数据用Excel 2010和SPSS 20.0统计软件进行统计分析;采用单因素方差分析法和Duncan多重比较法进行方差检验。
2 结果与分析
2.1 持续干旱及复水下接种不同ERM真菌对桃叶杜鹃幼苗叶绿素含量的影响
持续干旱及复水下接种不同ERM真菌对桃叶杜鹃幼苗叶绿素含量的影响如图1~图3所示,在相同干旱时间段内,接种ERM真菌极显著提高了桃叶杜鹃幼苗的叶绿素a含量、叶绿素b含量和叶绿素总含量(plt;0.01)。
如图1、图2所示,各处理幼苗的叶绿素a、叶绿素b含量随着干旱胁迫时间的延长呈现出先增后减的趋势,复水后显著上升。在持续干旱的当天,各接菌处理苗的叶绿素a、叶绿素b含量均高于ck,且差异极显著(plt;0.01)。在持续干旱第5天时各处理幼苗的叶绿素a、叶绿素b含量均有所增加,TY 29、TY 35菌根苗的增幅最大,而ck的增幅最小,其中TY 35的叶绿素a含量达到最高水平,较ck相比提高了75.0%,其次是TY 29、TY 02和TY 21,分别提高了61.1%、34.7%、24.6%,与ck间差异极显著(plt;0.01)。随着干旱时间的延长各处理幼苗的叶绿素a、叶绿素b含量则逐渐减少,在干旱15 d时均达到最低值,与干旱当天相比,TY 35、TY 29、TY 02和TY 21的叶绿素a含量分别降低了1.3%、3.3%、20.9%和23.9%,ck的降幅最大,达到了47.4%;菌根苗的叶绿素b含量变化幅度小于叶绿素a,TY 35、TY 29、TY 02和TY 21的叶绿素b含量分别降低了1.3%、0.4%、17.8和17.6%,ck的降幅最大,达到56.3%。复水后,各处理苗的叶绿素a、叶绿素b含量开始增加,且ck的增幅最小,TY 35和TY 29的增幅较大,超过干旱前水平。
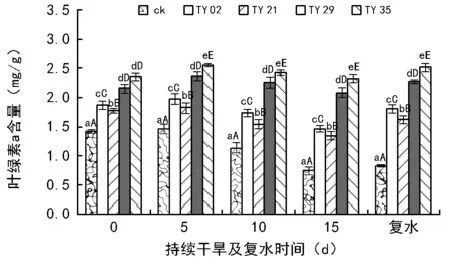
注:ck为未接种。柱状图上端不同小写字母表示差异达到显著水平(plt;0.05);不同大写字母表示差异达到极显著水平(plt;0.01)。
各处理苗的叶绿素a、叶绿素b含量呈现出先增后减的趋势,说明了适度的干旱胁迫能够促进桃叶杜鹃叶绿素的合成与积累,从而促进植物的光合作用以抵御干旱带来的伤害,但随着胁迫时间的延长,植物的叶片处于缺水状态,使叶绿素的合成受到影响,同时加速了叶绿素的分解,叶绿素含量降低。
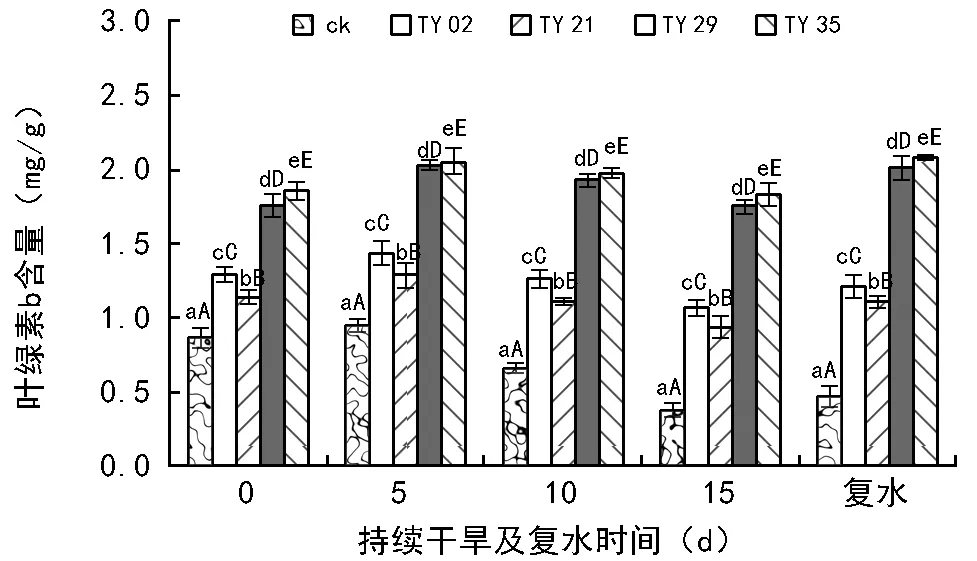
图2 持续干旱及复水后接种不同ERM菌株对桃叶杜鹃叶片叶绿素b含量的影响
桃叶杜鹃各处理苗在持续干旱条件下,叶绿素总含量的变化如图3所示,在相同干旱时间段内,菌根苗的叶绿素总含量均显著高于对照。说明接种能够显著提高桃叶杜鹃幼苗的叶绿素含量,从而促进幼苗的光合作用。随着干旱时间的延长,各处理苗的叶绿素总含量都不同程度的下降,ck的降幅最为明显,说明其对不利的干旱环境最为敏感。

图3 持续干旱及复水后接种不同ERM菌株对桃叶杜鹃叶片叶绿素总含量的影响
如图4所示,同一干旱时间条件下,接种苗的叶绿素a/b均低于对照,这可能是由于接种后桃叶杜鹃幼苗的叶绿素b增加大于叶绿素a所致。随着干旱时间的延长,菌根苗与对照苗的叶绿素a/b均呈先减后增的趋势,且ck的变化幅度最大,TY 29和TY 35的变化不明显;复水后,除TY 02和TY 21的叶绿素a/b增加外,其他处理苗的叶绿素a/b均减小,且ck的降幅最大。
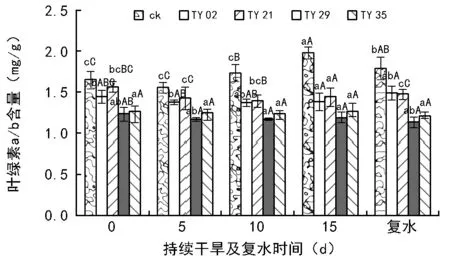
图4 持续干旱及复水后接种不同ERM菌株对桃叶杜鹃叶片叶绿素a/b含量的影响
2.2 持续干旱及复水下接种不同ERM真菌对桃叶杜鹃幼苗气体交换参数的影响
图5~图8分别反映了桃叶杜鹃幼苗净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、气孔导度(Gs)在持续干旱及复水条件下接种ERM真菌的变化趋势。如图5所示,接种ERM真菌极显著提高了桃叶杜鹃幼苗的净光合速率(Pn)(plt;0.01),随着干旱程度的增加,各处理苗的Pn均呈下降趋势,在干旱胁迫的第5天各菌根苗的下降幅度均较小,下降幅度在2.5%~11.5%之间,ck的降幅最大,达到19%。在干旱胁迫的第10天和第15天各处理苗的下降幅度逐渐增大,与干旱当天相比,第15天时ck的降幅达到90.5%,其次是TY 21和TY 02降幅分别为42.3%、31%,TY 29和TY 35降幅较小分别为20.1%和13.6%。在同一胁迫时间段,接菌苗的净光合速率均高于对照,且差异极显著(plt;0.01),随着干旱胁迫程度的增加,菌根苗的下降幅度均明显小于对照。
干旱条件下,植物以减少叶片气孔开度,降低蒸腾来抵御不利环境,而气孔导度减小将影响CO2的吸收,引起CO2供应不足,光合速率下降。如图6、图7所示,幼苗蒸腾速率(Tr)和气孔导度(Gs)的变化趋势与净光合速率(Pn)相似,都随着干旱胁迫程度的增加而降低,在同一胁迫时间段下,菌根苗的Tr和Gs均极显著高于对照(plt;0.01),且下降幅度均小于对照。由图8可见,持续干旱处理下各处理苗的胞间CO2浓度(Ci)随土壤水分含量的降低总体呈现出先减后增的趋势,其中ck、TY 02、TY 21在胁迫5 d时其Ci均下降,在胁迫10,15 d时升高;TY 29、TY 35则是在胁迫第5天和第10天呈下降趋势,在胁迫第15天上升。
复水5 d后,除ck外,各接菌苗的气体交换参数均得到不同程度的恢复,且TY 29和TY 35基本上能恢复到干旱前期的水平。与旱前相比,各接菌苗的Pn降幅不大,下降幅度在0.8%~1.6%之间,而ck则下降了90%。干旱胁迫显著降低了Tr和Gs,但复水后菌根苗的Tr和Gs均能得到较快恢复。复水后,TY 29和TY 35的Ci均能恢复到复水之前,ck、TY 02和TY 21较旱前有所升高,其中ck增幅最大达到20%。
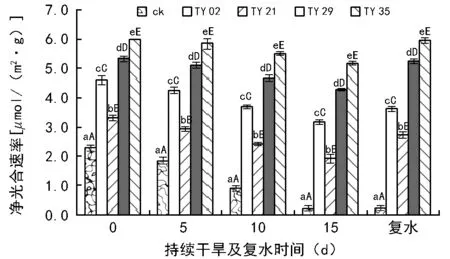
图5 持续干旱及复水后不同ERM菌株处理对桃叶杜鹃叶片净光合速率的影响

图6 持续干旱及复水后不同ERM菌株处理对桃叶杜鹃叶片蒸腾速率的影响

图7 持续干旱及复水后不同ERM菌株处理对桃叶杜鹃叶片气孔导度的影响
2.3 持续干旱及复水下接种不同ERM真菌对桃叶杜鹃幼苗叶片水分利用效率的影响
植物的水分利用效率(WUE)为Pn与Tr的比值,它在一定程度上可以反映植物的耗水性和对干旱的适应性。如图9所示,但在同一干旱处理时间下,桃叶杜鹃菌根苗的水分利用效率(WUE)均极显著高于对照;干旱第5天,除ck和TY 21外,其他菌株处理苗的WUE略高于干旱当天;在干旱第10天和第15天,ck的WUE开始急剧下降,菌根处理苗也呈下降趋势,但降幅均小于对照;持续干旱处理过程中,接种TY 29和TY 35的变化幅度较平缓,且在干旱第10天其WUE略大于干旱当天,即这2个菌株处理苗在干旱胁迫条件下能保持较高的水分利用效率。复水后,各处理苗的WUE均得到恢复,回升幅度为cklt;TY 21lt;TY 02lt;TY 29≈TY 35,其中,TY 29和TY 35基本能恢复到干旱胁迫前期的水平。
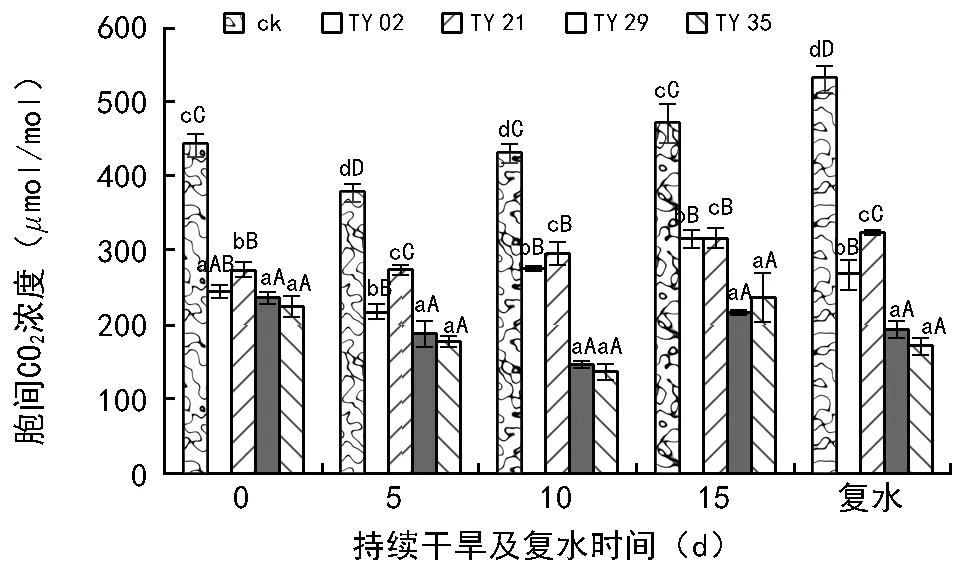
图8 持续干旱及复水后不同ERM菌株处理对桃叶杜鹃叶片胞间二氧化碳浓度的影响

图9 持续干旱及复水后不同ERM菌株处理对桃叶杜鹃叶片水分利用效率的影响
3 讨论与结论
叶绿素是植物光合色素中最重要的色素之一,其含量的高低标志着植物光合能力的强弱,叶绿素含量越高,说明苗木对光的利用率越高,则植株的光合作用越强。本研究结果表明,持续干旱条件下接种ERM真菌能够显著提高桃叶杜鹃幼苗的叶绿素a、叶绿素b含量和叶绿素总含量,相同干旱时间段内,各菌根苗的叶绿素含量均高于对照,这与张延旭[11]、郭辉娟[12]、高悦[13]等的研究结果一致。干旱胁迫会引起植物体内活性氧的积累,导致膜脂过氧化作用增强,叶绿素的合成受到破坏,分解速度加快,导致叶片中叶绿素含量下降及叶绿素a/b比值降低[14]。本试验中,随着干旱时间的延长,植物叶片的叶绿素a、叶绿素b含量和叶绿素总含量均不同程度下降,但各处理菌根苗叶绿素的降幅均明显小于对照,且TY 29和TY 35菌株在胁迫下均能保持相对较高的叶绿素含量,说明了接种ERM菌株能够增强植株对水分的吸收与利用,促进叶片叶绿素合成,降低光合机构的破坏程度,从而有利于叶片光合作用的进行。叶绿素a/b可以反映捕光色素复合体Ⅱ(LHCⅡ)在所有含叶绿素的结构中所占的比重,其值升高表明LHCⅡ含量的减少[15]。本研究结果显示,各处理苗的叶绿素a/b值均呈先减后增的趋势,菌根苗的叶绿素a/b值变化趋于稳定,ck的变化幅度较大,说明了超过一定程度的干旱,桃叶杜鹃幼苗通过升高叶绿素a/b值来减少叶片对光能的捕获,以降低光合机构遭受光氧化的破坏,这是其适应干旱胁迫的一种光保护调节机制。
本试验结果表明,干旱处理降低了桃叶杜鹃幼苗的净光合速率、蒸腾速率、气孔导度,但接种不同ERM菌株能不同程度地提高幼苗的净光合速率等光合指标,且在干旱条件下减缓其下降速度,这与张珊珊等[16]对蓝果树,田帅等[17]对刺槐的研究结果相似。干旱胁迫导致植物净光合速率下降的2个主要因素为气孔限制和非气孔限制。Farquhar等认为,在干旱胁迫下当Pn、Gs和Ci变化规律相同时,Pn的变化主要是由Gs引起的,反之如果Pn、Gs和Ci的变化不一致,则Pn的变化是由叶肉细胞活性决定的即非气孔限制[18]。试验结果表明,在干旱前期,桃叶杜鹃幼苗Gs和Ci的变化趋势与Pn相似,都随着干旱胁迫程度的增加而降低,而随着持续时间的增加,幼苗的Ci呈先减后增的趋势,说明在干旱初期(持续干旱第5天),桃叶杜鹃幼苗光合速率的下降主要是由气孔因素主导,到了干旱中期(持续干旱第10天),光合速率的下降以非气孔因素作为主导。复水后各菌根苗的气体交换参数均得到恢复,ck恢复较慢,TY 35和TY 29基本上能恢复到干旱前期的水平,说明了干旱胁迫没有对菌根苗造成不可逆转的伤害,接种ERM真菌能不同程度地提高了桃叶杜鹃幼苗对干旱的适应性以及旱后恢复能力。
水分利用效率(WUE)是植株对本身蒸腾消耗水量的利用能力,Heitholt等认为,对植物进行适当的干旱胁迫能够提高植株的WUE[19]。本试验中,在持续干旱的第5天,除ck和TY 21外,其他菌株处理苗的水分利用效率略高于干旱当天,表明适当的干旱胁迫能够提高植物的WUE,随着干旱胁迫加剧,植物水分利用效率均有所下降,且在同一干旱处理时间下,桃叶杜鹃菌根苗的水分利用效率(WUE)均极显著高于对照,说明接种ERM真菌能够有效地提高桃叶杜鹃对水分的吸收与利用,改善植物水分代谢状况,以增强植物抗旱性。
综上所述,接种ERM真菌显著提高了桃叶杜鹃幼苗的叶绿素含量、气体交换参数以及水分利用效率,同时降低叶片胞间CO2浓度,以增强桃叶杜鹃的光合作用。在干旱胁迫下,ERM菌株通过减缓各光合指标的下降速度,提高植株的水分利用效率,来缓解干旱胁迫对桃叶杜鹃幼苗所造成的伤害,增强桃叶杜鹃抵御干旱的能力,且以TY 35和TY 29的表现最好。本试验仅对干旱胁迫下桃叶杜鹃幼苗的叶绿素含量和气体参数等光合指标进行了研究,而叶绿体作为植物光合作用的主要细胞器,干旱条件下接种ERM真菌是如何通过影响植物叶绿体等细胞结构进行光合作用,从而引起植物光合参数的变化,还有待更加深入的研究。
[1]Read D J.The structure and function of the ericoid mycorrhizal root[J].Annals of Botany,1996,77:365-374.
[2]CAIRNEY J W G,MEHARG A A.Ericoid mycorrhiza:A partnership that exploits harsh edaphic conditions[J].European Journal of Soil Science,2003,54:735-740.
[3]刘仁阳,欧静.雷山杜鹃菌根的显微结构与菌根真菌的侵染率[J].贵州农业科学,2014(9):109-111.
[4]刘振华,姚娜,杨凯,等.树枫杜鹃菌根真菌分离与鉴定[J].林业科学研究,2012,25(6):795-797.
[5]李丽丽,杨洪一.杜鹃花类菌根真菌研究进展[J].北方园艺,2014(11):173-176.
[6]陈真,杨兵,张春英,等.锦绣杜鹃菌根真菌rDNA ITS序列分析及接种效应研究[J].菌物学报,2011,30(5):729-737.
[7]陈训,巫华美.中国贵州杜鹃花[M].贵阳:贵州科技出版社,2003:21.
[8]欧静,刘仁阳,陈训,等.菌根真菌对桃叶杜鹃种子萌发及幼苗移植的影响[J].南方农业学报,2013,44(2):293-298.
[9]欧静,韦小丽,何跃军,等.接种ERM真菌对桃叶杜鹃幼苗的促生效应及生理生化影响[J].林业科学,2013,49(7):48-56.
[10]邹琦.植物生理学实验指导[M].北京:中国农业出社,2000:72-75.
[11]张延旭,毕银丽,裘浪,等.接种丛枝菌根对玉米生长与抗旱性的影响[J].干旱地区农业研究,2015,33(2):91-94.
[12]郭辉娟,贺学礼.水分胁迫下AM真菌对沙打旺生长和抗旱性的影响[J].生态学报,2010,30(21):5 933-5 940.
[13]高悦,吴小芹.6种外生菌根菌对3种松苗叶绿素含量及叶绿素荧光参数的影响[J].南京林业大学学报(自然科学版),2010,34(6):9-12.
[14]张明生,谈锋.水分胁迫下甘薯叶绿素a/b比值的变化及其与抗旱性的关系[J].种子,2011,20(4):23-25.
[15]ANDERSON J M,ARO E M.Grana stacking and protection of photosystemⅡin thylakoid membranes of higher plant leaves under sustained high irradiance:a hypothesis[J].Photosynthesis Research,1994(41):315-326.
[16]张珊珊,康洪梅,杨文忠,等.干旱胁迫下AMF对云南蓝果树幼苗生长和光合特征的影响[J].生态学报,2016,36(21):1-13.
[17]田帅,刘振坤,唐明.不同水分条件下丛枝菌根真菌对刺槐生长和光合特性的影响[J].西北林学院学报,2013,28(4):111-115,208.
[18]Farquhar G D,Sharkey T D.Stomatal conductance and photosynthesis[J].Annual review of Plant Physiology,1982,33(3):317-345.
[19]Heitholt J J.Water use efficiency and dry matter distribution in nitrogen-and water-stressed winter wheat[J].Agron.J.,1989(81):464-469.
The Effects of Inoculation with ERM Fungi Under Drought Stress onPhotosynthetic Characteristics ofRhododendronannaeFranch. Seedlings
LONGHaiyan1,OUJing1,CHENDuanyu1,CHENRongjian1,ZHANGYuwu2
(1.College of Forest,Guizhou University,Guiyang 550025,China;2.Guizhou Institute of Biology,Guiyang 550009,China)
The seedlings of 1 a Rhododendron were used as materials,to study the effects of ERM fungi on the photosynthetic characteristics of Rhododendron fortunei seedlings under drought stress.The results showed that the chlorophyll content (Chl),net photosynthetic rate (Pn),transpiration rate (Tr),stomatal conductance (Gs) and water use efficiency (WUE) of Rhododendron fortunei seedlings were significantly increased by inoculation with ERM fungi.While reducing the intercellular CO2concentration (Ci),Compared with non-inoculation treatment,the difference was obvious.Under the condition of continuous drought,the rhizome of Rhododendron fortunei increased the utilization efficiency of water by promoting the synthesis and accumulation of chlorophyll in leaves,to slow down the decline of photosynthetic parameters of seedlings,so that seedlings maintain a high photosynthetic capacity;After rewatering, mycorrhizal fungi could restore their photosynthetic centers faster,and the recovery ability was significantly higher than that of control seedlings,and TY 35 and TY 29 could recover to the pre-drought level.It can be seen that inoculation of ERM fungi can significantly improve the photosynthetic capacity and water use efficiency of Rhododendron fortunei seedlings,so as to enhance the drought resistance of Rhododendron fortunei,and TY 35 and TY 29 has the best performance.
Ericoid mycorrhizal(ERM);Rhododendronannae; chlorophyll content;gas exchange parameters; drought stress
2017-02-24
国家自然科学基金项目(31560223;31460136);贵州省科技计划项目(黔科合支撑[2016]2522号);贵州省社发攻关项目(黔科合SY字[2015]3022号);贵州科学院科技计划项目(黔科院[2015]01号)。
龙海燕(1991—),女,在读硕士研究生,研究方向:园林植物与观赏园艺;E-mail:861032569@qq.com。
欧 静(1974—),女,教授,硕士研究生导师,主要从事园林植物资源利用与规划研究;E-mail:coloroj@126.com。
10.16590/j.cnki.1001-4705.2017.06.014
S 685.21
A
1001-4705(2017)06-0014-06
猜你喜欢
辽宁林业科技(2022年2期)2022-11-24
心声歌刊(2021年3期)2021-08-05
小小说月刊(2021年3期)2021-03-25
食药用菌(2021年4期)2021-01-08
饮食保健(2020年6期)2020-04-03
小聪仔(幼儿版)(2020年12期)2020-02-01
民族音乐(2018年5期)2018-11-17
乡村地理(2018年3期)2018-11-06
中国食用菌(2014年3期)2014-07-18
中成药(2014年8期)2014-02-28
