
黄河上游高寒草甸矮嵩草生理特征对流域不同生境的响应
2018-01-19 07:24李积兰李希来马福俊
草业学报 2018年1期
李积兰,李希来,马福俊
(1.青海大学农牧学院,青海 西宁 810016;2.青海大学生态环境工程学院,青海 西宁810016)
高寒矮嵩草(Kobresiahumilis)草甸是青藏高原主要植被类型之一,约占高原面积的33%[1],矮嵩草是构成矮嵩草草甸的主要建群种,属寒冷中生短根茎地下芽多年生莎草科嵩草属草本植物[2],是典型的密丛型草类,属高抗逆性物种[3],具有耐寒、耐旱、耐辐射、耐强风及耐贫瘠的特点[4],在长期对寒冷的适应进化和对人为放牧干扰的对抗过程中,形成了形态、营养、干旱、耐践踏等的稳定性维持机制,具有较高的系统稳定性与自我调控能力[5],并在放牧条件下,矮嵩草能成为群落优势种[6]。它是青藏高原高寒草甸草地的优良牧草,草质鲜嫩,营养价值丰富,具有“四高一低”典型特性,其总营养物质的含量在各经济类群中居首位,一年四季均为家畜喜食[7],是高寒草甸地区生态和经济价值兼优的牧草[8]。在所有的草地类型中矮嵩草、杂类草草地型生态系统服务价值最大[9],它的良好生长不仅对维持青藏高原的生态平衡起着重要作用[10]。但由于各种自然因素和人为因素的影响,高寒草甸生态系统被破坏,草地退化严重,其中以矮嵩草草甸的破坏最为典型,草甸出现秃斑,草地变为“黑土滩”[5]。
植物与其生存环境的适应关系是矛盾统一体的两个方面,一方面导致植物体一系列形态学、生理学、生物化学和分子生物学的变化,影响植物的生长,甚至危害其生存;另一方面,植物为了生存会使它的机体结构和功能发生改变[11]。有关矮嵩草环境适应性的研究从80年代至今一直在其生物生态学特性、繁殖特性、遗传特性以及对放牧、施肥、刈割等干扰环境的适应以及适应环境的形态结构、生理生态特征等方面进行了系统研究[12-19],但是多数研究集中在对不同海拔梯度的适应性,三江源区以山地地貌为主,长江、黄河及其支流、澜沧江河谷切割较深,形成许多台地和谷底[20],形成许多大小不等的流域,由于地形的不同,造成不同生境下水、热、养分因子的空间异质性,从而使得植被分布存在一定的差异性,而在此生境下矮嵩草生理性的研究尚未见报道。因此本研究选择三江源区小流域不同生境,分析不同立地生境各生态因子综合作用下矮嵩草生理生化指标的变化特征,并探索测定指标与土壤环境因子之间的相互关系,试图探讨矮嵩草适应流域不同生境的生理机制,为进一步研究高寒草甸优势种矮嵩草的适应性提供一定的参考。
1 材料与方法
1.1 研究区自然概况
研究区位于青海省河南蒙古族自治县南旗村黄河二级支流夏吾曲河小流域。地理坐标34°51.571′ N,101°28.102′ E,平均海拔3590 m。该区域属高原大陆性气候,全年四季特征不明显,每年5-10月温暖、多雨,11月至次年4月寒冷、干燥、多大风天气。年平均气温为0.0 ℃,最冷月平均气温为-10.6 ℃,最热月的平均气温为9.4 ℃。年降水量597.1~615.5 mm,年平均蒸发量为1349.7 mm。年日照时数2551.3~2577.2 h,日照率58.0%。
1.2 样地选择
2015年7月29日-8月10日,选择南旗村黄河二级支流夏吾曲河小流域1个,该样地为自然放牧地,放牧压力一致。在小流域中地形依次设立山顶(hill top,HT)、山坡(hill slope,HS)、滩地(beach land,BD)和1级阶地[21](terrace 1,T1)、2级阶地(terrace 2,T2),地形生境描述见图1。
采用样线法和网格法相结合的取样方法,在小流域每种生境设大样方50 m×50 m,重复3次,进行矮嵩草取样。每个大样方的对角线上设3个1 m×1 m的小样方进行植被调查,调查样方内出现的物种、数量及其盖度,并剪取植物地上生物量在60 ℃下24 h烘干称重,矮嵩草重要值=(相对高度+相对生物量+相对盖度)/3,植被特征见表1。同时进行土样取样,采用对角线法,用直径为3 cm不锈钢土钻进行采样,样地土壤环境因子见表2,小流域50 m×50 m样方有15个,小样方有45个。

图1 小流域地形及生境描述Fig.1 Description of basin habitats ①河道;②1级阶地,属河漫滩湿地,以藏嵩草为主;③2级阶地,属阶地湿地[21],以青藏苔草、藏嵩草和海韭菜为主;④滩地,属高寒草甸,以矮嵩草、高山嵩草和紫花针茅、垂穗披碱草为主。⑤陡坡地(坡度24°),属高寒草甸,以紫花针茅、垂穗披碱草、高山嵩草和矮嵩草为主;⑥山顶,属高寒草甸,以高山嵩草、紫花针茅和矮嵩草为主。①River; ②Terrace 1 (T1), floodplain wetland, it is dominated by cold-tolerant species such as Kobresia tibetica; ③ Terrace 2 (T2), terrace wetland, it is dominated by species such as Kobresia tibetica, Cara moocroftii, Triglochin maritimum; ④Beach land (BD), alpine meadow, it is dominated by species such as Kobresia humilis, Elymus nutans, Potentilla anserine; ⑤Hill slope (HS), alpine meadow,it is dominated by species such as Stipa purpurea, Elymus nutans; ⑥Hill top (HT), alpine meadow, it is dominated by species such as Kobresia pygmaea, Stipa purpurea.
1.3 样品采集与处理
采样时间为植物生长旺季,7月底-8月初,每个大样方内采集矮嵩草茎叶,装入自封袋密封,并立即放入自制冰盒中,低温下(<4 ℃以下)带回实验室测定。同时采取样方中植物根系主要分布的0~30 cm土层的土壤,带回实验室测定土壤理化性质指标。
1.4 测定指标及方法
土壤指标的测定:土壤温度、湿度用美国TETRACAM公司生产的Easy Test便携式土壤三参数测量仪(测定时间和生境土壤取样时间同步)和WatchDog1400土壤温湿度记录仪测定;土壤有机质的测定用重铬酸钾容量法;全氮的测定用凯氏定氮法;速效氮的测定用硫酸钠—纳氏试剂比色法;全磷的测定用酸溶-钼锑抗比色法;速效磷的测定用Olsen法;全钾、速效钾的测定采用原子吸收法;pH用pH计测定[22]。
植物指标的测定:用北京普析通用仪器有限责任公司生产的T6系列紫外分光光度计测定,可溶性蛋白的测定采用考马斯亮蓝G-250染色法;可溶性糖含量的测定采用苯酚法;脯氨酸含量的测定用磺基水杨酸法;丙二醛含量的测定用硫代巴比妥酸法;过氧化氢酶活性采用紫外吸收法;叶绿素含量测定用丙酮乙醇混合液法[23]。
1.5 数据处理
采用Excel 2010对数据进行整理,用SPSS for Windows 20.0进行方差分析和相关分析,结果用平均值±标准偏差表示,用单因素方差分析(ANOVA)和最小显著差异法(LSD)比较不同数据组间的差异,差异显著性水平设定为α=0.05;相关显著性用Pearson进行检验,用Sigma Plot 10进行绘图。

2 结果与分析
2.1 流域不同生境矮嵩草生理特征
研究结果表明(图2),矮嵩草丙二醛含量由坡地、山顶、2级阶地及滩地依次降低,其含量由坡地的10.24 nmol·g-1降低至山顶的8.38 nmol·g-1、2级阶地的7.01 nmol·g-1、滩地的4.80 nmol·g-1,坡地较滩地增加了2.1倍。坡地、山顶、滩地、2级阶地间均差异显著(P<0.05)。
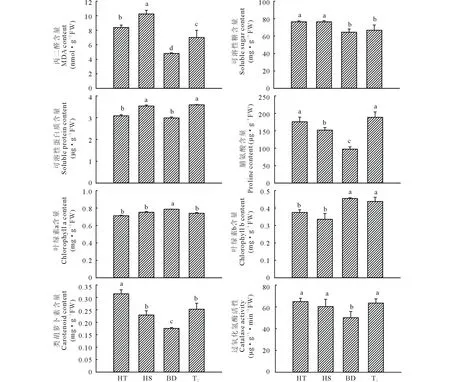
图2 流域不同地形生境下矮嵩草叶片生理指标变化特征Fig.2 Physiological characteristics of K. humilis at different sites HT:山顶 Hill top;HS:坡地 Hill slope;BD:滩地 Beach land;T2:2级阶地 Terrace 2.图中不同小写字母表示处理间多重比较差异显著(P<0.05)。Different lowercase letters in the Figure show significant difference (P<0.05) of multiple comparisons between the treatments.
矮嵩草可溶性糖含量由山顶、坡地及2级阶地、滩地依次降低,其含量由山顶、坡地平均76.31 mg·g-1降低至2级阶地、滩地平均的65.57 mg·g-1,山顶、坡地间差异不显著(P>0.05),与2级阶地、滩地间差异显著(P<0.05)。矮嵩草可溶性蛋白含量在坡地、2级阶地含量较高,二者间差异不显著(P>0.05),与其他生境间差异显著(P<0.05),滩地含量最低,此含量较坡地、2级阶地平均降低了0.58 μg·g-1,滩地与山顶间差异不显著(P>0.05)。矮嵩草脯氨酸含量2级阶地>山顶>坡地>滩地,由2级阶地、山顶平均的182.21 μg·g-1降低至坡地的150.51 μg·g-1、滩地的97.29 μg·g-1。除山顶、2级阶地间差异不显著外(P>0.05),其他生境间均差异显著(P<0.05)。
矮嵩草叶绿素a、b含量均在滩地生境最高,山地和坡地较低,其中叶绿素a含量在滩地与其他生境间差异显著(P<0.05),其他生境间差异不显著(P>0.05)。其含量由滩地的0.78 mg·g-1降低至2级阶地、坡地、山顶平均的0.73 mg·g-1。叶绿素b含量在滩地与2级阶地、山顶与坡地间差异不显著(P>0.05),其他生境间均差异显著(P<0.05),其含量由滩地、2级阶地平均的0.45 mg·g-1降低至山顶、坡地平均的0.35 mg·g-1。
矮嵩草类胡萝卜素含量山顶最高,与其他生境差异显著(P<0.05),滩地含量最低,与其他生境差异显著(P<0.05),坡地与2级阶地间差异不显著(P>0.05)。其含量由山顶的0.31 mg·g-1分别降低至2级阶地、坡地平均的0.23 mg·g-1,滩地的0.18 mg·g-1。
矮嵩草过氧化氢酶活性(CAT)在山顶最高,与坡地、2级阶地间差异不显著(P>0.05),滩地含量最低,与其他生境差异显著(P<0.05),其活性由山顶、坡地、2级阶地平均的62.85 μg·g-1·min-1减小至滩地的50.05 μg·g-1·min-1,降低了25.56%。
2.2 矮嵩草生理指标与土壤因子相关性分析
通过对流域不同生境矮嵩草生理指标与土壤因子相关性分析结果表明(表3),矮嵩草生理生化指标与土壤中的全钾、pH、湿度相关性较大,其中土壤中的全钾与可溶性蛋白质呈极显著负相关(P<0.01),与叶绿素a和b、类胡萝卜素呈显著负相关(P<0.05);pH与可溶性蛋白、脯氨酸、类胡萝卜素呈极显著正相关(P<0.01),与过氧化氢酶呈显著正相关(P<0.05);土壤湿度除与叶绿素a和b呈显著正相关(P<0.05),与其他指标均呈负相关,其中与可溶性蛋白质呈极显著负相关(P<0.01)。土壤中的全磷与叶绿素b呈显著正相关(P<0.05),其他生理指标与土壤因子间无显著相关性(P>0.05)。

表3 矮嵩草生理指标与土壤环境因子相关性Table 3 Physiological indexes of K. humilis associated with soil factors
注:*在0.05水平(双侧)上显著相关,**在0.01水平(双侧)上极显著相关。
Note:* Significant correlation was found at the 0.05 level (bilateral),**Significant correlation was found at the 0.01 level (bilateral).
各生理指标间的相关分析结果表明,可溶性蛋白质与叶绿素a和过氧化氢酶呈显著正相关(P<0.05),与类胡萝卜素呈极显著正相关(P<0.01);脯氨酸与过氧化氢酶呈显著正相关(P<0.05);丙二醛与叶绿素b呈显著正相关(P<0.05);叶绿素a与叶绿素b、类胡萝卜素呈极显著正相关(P<0.01),与过氧化氢酶呈显著正相关;叶绿素b与类胡萝卜素呈极显著正相关(P<0.01);类胡萝卜素与过氧化氢酶呈显著正相关(P<0.05)。
3 讨论
3.1 植被结构对小流域生境的响应
环境控制和塑造了植物的生理过程、形态特征和地理分布;植物则在适应环境的同时,改造和影响着环境,二者相互影响、相互制约。在不同的光照、热量、水分等环境条件下,植物的群落结构、形态特征、生理过程和地理分布等方面有很大的差异性[24]。
小流域不同地形下生境土壤理化性质中土壤湿度、全氮、全磷、碱解氮、速效磷、有机质由1级阶地、2级阶地、山顶、滩地、坡地依次降低,土壤pH由山顶、坡地、1级阶地、2级阶地、滩地降低,全钾含量在坡地和滩地较高,1级阶地、2级阶地较低,速效钾含量滩地最高,坡地最低,总变化趋势是2级阶地、1级阶地、滩地、山顶中较高,坡地的含量较低(表2),这可能由于小流域中从山顶到1级阶地有明显的高度落差,降水后形成的地表径流、壤中流导致水分、养分积累于地势低洼区域,且水分蒸发量较少,因物理作用使得滩地、1级阶地、2级阶地土壤中含水量较高,土壤肥力较好。由于流域生境状况的差异,物种数由山顶、坡地、滩地、2级阶地、1级阶地依次降低,而总生物量在2级阶地、1级阶地、滩地中较高,山顶和坡地最低,其变化梯度与流域生境土壤理化性质变化梯度一致,说明滩地、2级阶地、1级阶地植物生产力较高,生境条件较好,这与土壤湿度适宜、营养物质含量高时,植物生长良好的[25-27]研究结果一致。从矮嵩草分布范围来看,矮嵩草植物除在1级阶地河漫滩湿地无分布外,其他生境均有分布,且在滩地为优势种,其盖度、重要值、生物量及生物量比重均最高,坡地次之,在2级阶地的阶地湿地最低(表1),呈零星分布,这可能与生境土壤湿度有关。因此,矮嵩草生态幅较广,具有适应不同生境的能力,这与前人研究的植物在不同植被带不同侵蚀环境下样地内出现频率较高,预示着此物种可以适应不同的生境,有着较强的适应性研究结果一致[28-29]。从小流域不同地形的生境植被结构来看,在正常放牧情况下矮嵩草植物在滩地所在生境下生物量、盖度、重要值最高,坡地次之,则矮嵩草植物在滩地处适合度较高,结合前人研究的结果矮嵩草不放牧的情况下与群落中的高大禾本科植物竞争时处于劣势,而在较重的放牧情况下,采食作用可消除不耐牧的禾本科植物的竞争影响[30],因此如果禁牧或土壤水分含量(>38%)继续增加,矮嵩草植物与其他植物竞争作用下,滩地高寒植被生境可能不利于矮嵩草植物的生长,如果放牧压力增大、土壤含水量适当的情况下,滩地高寒草甸可能成为矮嵩草植物生长的适宜生境。
3.2 矮嵩草生理特性对小流域生境的响应
植物在长期的进化中为了适应环境,不仅在植物形态结构表型上表现出可塑性,同时调节着自身体内生理平衡[31]。当植物受到环境胁迫时,植物细胞内活性氧自由基的产生与清除的动态平衡被破坏,致使活性氧自由基大量积累,对植物造成伤害,从而使膜脂发生过氧化作用或膜脂脱脂作用,形成丙二醛(MDA),MDA反映了植物细胞膜的受伤害程度[32],本研究中,矮嵩草MDA含量在坡地、山顶较高,滩地最低,说明矮嵩草在滩地处受伤害程度最小,细胞膜未受到破坏。
渗透调节是植物抗旱性的一种重要生理机制,植物有机渗透物质主要有可溶性糖、可溶性蛋白和脯氨酸。可溶性糖含量增加可使原生质脱水后仍可保持其基本结构,同时增大了细胞液浓度,提高了对水分的吸收能力[33]。可溶性蛋白含量的变化可以反映细胞内蛋白质合成、变性及降解等信息,也可结合水分降低细胞的渗透势以维持细胞的膨压[34]。脯氨酸在植物抗逆过程中的主要作用有两点:一是作为渗透调节物质保持原生质与环境的渗透平衡,有效吸水和防止水分散失;二是与蛋白质相互作用增加蛋白质的可溶性和减少可溶性蛋白的沉淀,从而保持细胞膜结构的完整性和使酶免受伤害[35]。本研究发现山顶、坡地和2级阶地中矮嵩草处于胁迫状态,体内可溶性糖、可溶性蛋白、脯氨酸含量增加,尤其是脯氨酸含量变化较明显,可能是用以调节植物体内的水分平衡,同时减小环境对植物细胞的伤害,植物受到水分胁迫,使得体内可溶性糖、可溶性蛋白和脯氨酸含量增大,植物的渗透调节能力提高,而这些物质的积累可以降低植物细胞内的水势,防止水分外流,起到保护细胞内大分子结构的作用[32]。此含量在滩地中最低,说明滩地高寒草甸生境条件适宜,矮嵩草尚未处于胁迫状态。
过氧化氢酶(CAT)活性是植物体内酶催活性氧自由基清除系统之一,可有效地消除活性氧对细胞的伤害,阻止膜的过氧化和被破坏[36]。植物体过氧化物体和线粒体中的H2O2由CAT催化分解成H2O和O2,本研究结果表明,山顶、坡地、2级阶地中的CAT活性较高,滩地中的CAT最低,说明矮嵩草在山顶、坡地和2级阶地受到的胁迫已经干扰到了植物活性氧清除系统的正常循环。
叶绿素含量可以反映植物生长发育状况,反映植物潜在光合作用能力大小,胁迫会加速叶片衰老,抑制叶绿素合成,并加速其分解,使叶绿素浓度降低[35],本研究中叶绿素a、b含量均在滩地生境最高,山地、坡地和2级阶地较低。
植物体内存在的非酶促清除系统类胡萝卜素(Car),可以直接或间接地清除系统中的活性氧[26],本研究中矮嵩草Car含量山顶、坡地、2级阶地较高,滩地含量最低。由于生境的差异,在植物干旱强光照的山顶、山坡植物细胞膜受到胁迫最严重,细胞膜受的伤害程度最大,而且干旱使叶绿素分解作用加强,同时水分胁迫也影响叶绿素的合成速率,干旱胁迫会打破植物体内活性氧的产生和清除系统之间的平衡,细胞内活性氧增多导致细胞膜稳定性降低和膜脂过氧化等,严重损害植物的生长[36]。
总之矮嵩草渗透调节物质、细胞伤害程度物质、抗逆酶活性物质和非酶活性物质表现为山顶、坡地、2级阶地>滩地,叶绿素a,b含量呈相反趋势。说明在山顶、坡地、2级阶地矮嵩草植物受伤害程度有所增加,表现出了抗逆性增强的趋势。这可能是由于坡地坡度大、光线强,同时该生境下土壤侵蚀造成的水分养分流失,植物生长受到的干扰强度较大,使得该生境下矮嵩草细胞内积累超氧根离子量、丙二醛、可溶性糖、可溶性蛋白、脯氨酸、类胡萝卜素、CAT量增加,叶绿素a及叶绿素b含量减小,而在生境较好的滩地上述生理指标呈相反趋势,有研究表明土壤营养物质含量高,植物渗透调节物质含量均不会很高,植物不处于逆境中,还未启动保护酶等调节自身抗性,故在水分和养分较高的滩地矮嵩草植物尚未表现出抗性。山顶、2级阶地虽然土壤养分、水分含量相对较大,但山顶空气流动速度快、光照较强,且2级阶地表层土壤含水量较低(5.74%),故矮嵩草在山顶和2级阶地生理指标变化与坡地相似。矮嵩草在不同生境下的生理变化特征与其他学者研究胁迫生境下植物生理反应结果类似[35,37-40]。
3.3 矮嵩草生理特性与土壤环境因子的关系
矮嵩草生理特性与流域生境的土壤环境因子相关性表明,矮嵩草植物生理特性与土壤湿度、pH和全钾相关性较高,因此土壤湿度、pH和全钾可能是影响矮嵩草生长的关键因子,在滩地中土壤湿度适中,pH最低,全钾、速效钾含量最高。土壤pH与矮嵩草植物可溶性蛋白质、叶绿素a、类胡萝卜素和过氧化氢酶呈显著正相关,这与崔喜艳等[41]研究的随着pH值从4.5上升到7.5,烤烟叶中蛋白质含量、 叶绿素含量和酶活性呈上升趋势的结果一致。钾是植物必需营养元素之一,有研究表明钾是植物细胞中最丰富的离子,在植物的生长和发育中起重要作用,如促进酶的活化、蛋白质合成、光合作用,维持渗透压等[41]。而本研究土壤中的全钾含量与可溶性蛋白质、脯氨酸、类胡萝卜素、过氧化氢酶含量呈显著负相关,流域生境山顶、2级阶地全钾含量较低,而在坡地、滩地含量较高(表1)。青藏高原的低温、缺水环境可导致植物受到水分胁迫的影响[42],干旱胁迫常导致植物内活性氧的积累,从而对植物造成氧化胁迫。研究认为丙二醛、脯氨酸、可溶性蛋白质、可溶性糖、抗氧化物酶活性等可以作为植物抗逆性生理评价的重要指标,细胞积累渗透调节物质以保持膨压维持植物的生理过程,MDA 含量反映植物细胞膜的受伤害程度,可溶性糖的积累有利于提高细胞膨压,脯氨酸含量增加可防止细胞脱水等[43-44],强光和水分胁迫下叶绿素含量降低[45]。本研究表明土壤湿度与矮嵩草可溶性蛋白质、丙二醛、类胡萝卜素、过氧化氢酶活性呈显著负相关,与叶绿素a、b呈显著正相关。说明矮嵩草植物在逆境生境(水分含量低、土壤养分贫瘠)中通过生理可塑性减小组织代谢活动、增加抗逆物质积累来提高自身抗逆性以适应不同生境。
细胞内越高的可溶性蛋白含量,能够使细胞维持越低渗透势,渗透调节作用与可溶性蛋白含量大小密切相关,高的可溶性蛋白含量能够抵抗逆境胁迫对细胞形成的损害[46]。矮嵩草体内生理指标间的相关表明在逆境中通过增加其可溶性蛋白含量,维持低渗透性来抵抗生境。同时也有研究表明植物竞争通过增加体内的渗透调节物质和增强体内的抗氧化酶系统来适应岩溶高原干旱、少土、寒冷的恶劣环境[46]。因此矮嵩草植物在滩地中未受到胁迫影响,适宜生长在水热养分环境适宜的滩地生境。
4 结论
流域不同生境土壤理化性质含量在1级阶地、2级阶地、滩地、山顶中较高,在坡地的含量较低。由于流域生境状况的差异,物种数由山顶、坡地、滩地、2级阶地、1级阶地依次降低,而总生物量在2级阶地、1级阶地、滩地中较高,坡地最低,其变化梯度与流域生境土壤理化性质变化梯度一致,因此1级阶地、2级阶地、滩地所在生境条件较好。而矮嵩草植物除在1级阶地无分布外,其他生境均有分布,且在滩地为优势种,其盖度、重要值、生物量及生物量比重均在滩地最高,坡地次之,在2级阶地最低。
山顶、坡地光线强,加之该环境下土壤侵蚀造成的水分养分流失,以及2级阶地生境表层土壤比较干旱,矮嵩草植物生长受到的干扰强度较大,使得该生境下矮嵩草体内丙二醛含量、可溶性蛋白含量、可溶性糖含量、脯氨酸含量、类胡萝卜素含量、过氧化氢酶活性增加,叶绿素a、b含量减小,而在滩地生境中以上指标呈现相反趋势。
矮嵩草生理生化特性与流域生境的土壤环境因子中的土壤湿度、pH和全钾相关性较高,因此土壤湿度、pH和全钾可能是影响矮嵩草生长的关键因子。
因此,通过对小流域内矮嵩草分布状况及生理生化特征的研究,矮嵩草生态幅较广,喜适于pH较低、水热养分条件适宜的滩地高寒草甸植被生境;在胁迫生境下如土壤含水量较低、贫瘠,通过增加体内抗逆物质活性和竞争来适应立地生境。
致谢:夏略、东霞、程世军、朱恩泽对论文给予了实验帮助,特此致谢。
References:
[1] Lin L, Li Y K, Zhang F W,etal. Principal component analysis on alpineKobresiahumilismeadow degradation succession in Qinghai-Tibetan plateau. Chinese Journal of Grassland, 2012, 34(1): 24-30.
林丽, 李以康, 张法伟, 等. 青藏高原高寒矮嵩草草甸退化演替主成分分析. 中国草地学报, 2012, 34(1): 24-30.
[2] Zhou X M, Wang Q J, Zhang Y Q,etal. Quantitative analysis of succession law of the alpine meadow under the different grazing intensities. Acta Phytoecologicaet Geobotanica Sinica, 1987, 11(4): 276-285.
周兴民, 王启基, 张堰青, 等. 不同放牧强度下高寒草甸植被演替规律的数量分析. 植物生态学与地植物学报, 1987, 11(4): 276-285.
[3] Wang C T, Long R J, Ding L M. Study of alpine meadow of basic characteristic in Qinghai Tibet Plateau. Pratacultural Science, 2004, 21(8): 16-19.
王长庭, 龙瑞军, 丁路明. 青藏高原高寒嵩草草甸基本特征的研究. 草业科学, 2004, 21(8): 16-19.
[4] Li Q X, Zhao Q F, Ma S R,etal. Research progress onKobresiaspecies. Journal of Northwest Normal University (Natural Science Edition), 2006, 42(6): 78-82.
李巧峡, 赵庆芳, 马世荣, 等. 嵩草属植物研究进展.西北师范大学学报(自然科学版), 2006, 42(6): 78-82.
[5] Cao G M, Long R J. System stability and its self-maintaining mechanism by grazing in alpineKobresiameadow. Chinese Journal of Agrometeorology, 2009, 30(4): 553-559.
曹广民, 龙瑞军. 放牧高寒嵩草草甸的稳定性及自我维持机制. 中国农业气象, 2009, 30(4): 553-559.
[6] Zhu Z H, Sun S Q. Characteristic responsiveness on modular populations ofKobresiahumilisto grazing in alpine meadow. Aata Betanica Sinica, 1996, 38(8): 653-660.
朱志红, 孙尚奇. 高寒草甸矮嵩草种群的放牧中构件种群的反应特性. 植物学报, 1996, 38(8): 653-660.
[7] Yang Y W, Li X L, Li J L,etal. Growth response ofKobresiahumilisto grazing disturbance on alpine meadow. Acta Agriculturae Boreali-Occidentalis Sinica, 2011, 20(9): 18-24.
杨元武, 李希来, 李积兰, 等. 高寒草甸矮嵩草对放牧扰动的生长反应. 西北农业学报, 2011, 20(9): 18-24.
[8] Yang Y W, Li X L. Pilot study of clonal growth and reproduction ofKobresiahumilis. Journal of Qinghai University (Nature Science Edition), 2008, 26(1): 31-34.
杨元武, 李希来. 矮嵩草克隆生长与繁殖的初步研究. 青海大学学报(自然科学版), 2008, 26(1): 31-34.
[9] He L. The evaluation on ecosystem services value of natural meadow in Maqin county. Journal of Anhui Agricultural Sciences, 2011, 39(10): 6045-6047.
何玲. 玛沁县天然草地生态系统服务功能价值评价. 安徽农业科学, 2011, 39(10): 6045-6047.
[10] Li X L, Zhu Z H, Qiao Y M,etal. Study on ramet modular ofKobresiahumilisclonal under different stocking intensity in alpine meadow. Chinese Qinghai Journal of Animal and Veterinary Science, 2001, 31(3): 7-9.
李希来, 朱志红, 乔有明, 等. 不同放牧强度下高寒草甸矮嵩草无性系分株构件的研究. 青海畜牧兽医杂志, 2001, 31(3): 7-9.
[11] Jian L C, Wang H. Adversity Plant Cell Biology. Beijing: Science Press, 2009.
简令成, 王红. 逆境植物细胞生物学. 北京: 科学出版社, 2009.
[12] Zhou X M. A preliminary study of morphological-ecological characteristics of eight species of genusKobresiain Qinghai-Tibet Plateau. Acta Botanica Sinica, 1979, 21(2): 135-142.
周兴民. 青藏高原嵩草属八种植物的形态—生态学特性的初步研究. 植物学报, 1979, 21(2): 135-142.
[13] Jiang G C, Jia X H. Comparison of the microstructures of vegetative organs of three species ofKobresiagrowing in different altitude. Journal of Henan University (Natural Science Edition), 1999, 29(1): 63-68.
姜罡丞, 贾晓红. 不同海拔高度上的嵩草属三个种营养器官显微结构的比较. 河南大学学报(自然科学版), 1999, 29(1): 63-68.
[14] Han F, Ben G Y, Shi S B,etal. Comparative study on the resistance ofKobresiahumilisgrown at different altitudes in Qinghai-Xizang plateau. Acta Ecologica Sinica, 1998, 18(6): 654-659.
韩发, 贲桂英, 师生波, 等. 青藏高原不同海拔矮嵩草抗逆性的比较研究. 生态学报, 1998, 18(6): 654-659.
[15] Zhu Z H, Li X L, Sun H Q,etal. Effects of different grazing intensity on green up ofKobresiahumilismeadow. Pratacultural Science, 2002, 19(8): 52-57.
朱志红, 李希来, 孙海群, 等. 不同放牧强度对矮嵩草草甸群落返青的影响. 草业科学, 2002, 19(8): 52-57.
[16] Li Y K, Lin L, Zhang F W,etal.Kobresiapygmaeacommunity-disclimax of alpine meadow zonal vegetation in the pressure of grazing. Journal of Mountain Science, 2010, 28(3): 257-265.
李以康, 林丽, 张法伟, 等. 小嵩草群落——高寒草甸地带性植被放牧压力下的偏途顶极群落. 山地学报, 2010, 28(3): 257-265.
[17] Zhu Z H, Wang G, Wang X A. Grading responses of clonalKobresiahumilisto grass cutting. Acta Botanica Boreali-Occidentalia Sinica, 2005, 25(9): 1833-1839.
朱志红, 王刚, 王孝安. 克隆植物矮嵩草在刈割条件下的等级反应研究. 西北植物学报, 2005, 25(9): 1833-1839.
[18] Cui Y. Eco-anatomical Study onKobresiaWilld.of Eastern Qinghai-Tibet Plateau. Lanzhou: Northwest Normal University, 2006.
崔艳. 青藏高原东部嵩草属植物的生态解剖学研究. 兰州: 西北师范大学, 2006.
[19] Li J L, Li X L. Research progress on environmental adaptability ofKobresiahumilisin alpine meadow. Ecological Science, 2016, 35(2): 156-165.
李积兰, 李希来. 高寒草甸矮嵩草的环境适应性研究进展. 生态科学, 2016, 35(2): 156-165.
[20] Chen G, Li X L, Gao J Q,etal. Wetland Types and Evolution and Rehabilitation in the Sanjiangyuan Region. Xining: Qinghai people’s Publishing House, 2011.
陈刚, 李希来, 高家庆, 等. 三江源区湿地类型与演变和修复. 西宁: 青海人民出版社, 2011.
[21] Gao J, Li X L, Gary B,etal. Geomorphic-centered classification of wetlands on the Qinghai-Tibet Plateau, Western China. Journal of Mountain Science, 2013, 10(4): 632-642.
[22] Bao S D. Soil Agricultural Chemistry Analysis. Beijing: China Agriculture Press, 2010.
鲍士旦. 土壤农化分析. 北京: 中国农业出版社, 2010.
[23] Kong X S, Yi X F. Experimental Techniques of Plant Physiology. Beijing: China Agriculture Press, 2008.
孔祥生, 易现峰. 植物生理学实验技术. 北京: 中国农业出版社, 2008.
[24] Li F L, Bao W K. Responses of the morphological and anatomical structure of the plant leaf to environmental change. Chinese Bulletin of Botany, 2005, 22(Supplement 1): 118-127.
[25] Ye C J, Zhao K F. Effects of adaptation to elevated salinity on some enzymes’ salt-tolerance in vitro and physiological changes of eelgrass. Acta Botanica Sinica, 2002, 44(3): 788-794.
[26] Pan H L, Li M H, Cai X H,etal. Responses of growth and eco-physiology of plants to altitude. Ecology and Environmental Sciences, 2009, 18(2): 722-730.
潘红丽, 李迈和, 蔡小虎, 等. 海拔梯度上的植物生长与生理生态特性. 生态环境学报, 2009, 18(2): 722-730.
[27] Zhang X H. Study on Population Distribution, Morphological Characteristics and Physiological-biochemical Characteristics ofDactylisglomeratain East and West Section of Tianshan Mountain North Slope. Wulumuqi: Xinjiang Agricultural University, 2015.
张鲜花. 天山北坡东段与西段野生鸭茅的种群分布、形态特征及生理生化特性研究. 乌鲁木齐: 新疆农业大学, 2015.
[28] Brown J H. On the relationship between abundance and distribution of species. The American Naturalist, 1984, 124: 255-279.
[29] Rosenzweig M. Species Diversity in Space and Time. Cambridge: Cambridge University Press, 1995.
[30] Chen B, Zhou X M. Analysis of niche breadths and overlaps of several plant species in theKobresiacommunities of an alpine meadow. Acta Phytoecologica Sinica, 1995, 19(2): 158-169.
陈波, 周兴民. 三种嵩草群落中若干植物种的生态位宽度与重要分析. 植物生态学报, 1995, 19(2): 158-169.
[31] Martinez J P, Kient J M, Bajji M,etal. NaCl alleviates polyethylene glycol-induced water stress in the halophyte speciesAtriplexhalimusL. Journal of Experimental Botany, 2005, 56(419): 405-410.
[32] Johnson S M, Doherty S J, Croy R R D. Biphasic superoxide generation in potato tubers: a self amplifying response to stress. Plant Physiology, 2003, 13: 1440-1449.
[33] Iannucci A, Russo M, Arena L,etal. Water deficit effects on osmotic adjustment and solute accumulation in leaves of annual clovers. European Journal of Agronomy, 2002, 16: 111-122.
[34] Irigoyen J J, Einerich D W, Sánchez-Díaz M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicagosativa) plants. Physiologia Plantarum, 1992, 84: 55-60.
[35] Turner N C, Kramer P J. Adaptation of Plants to Water and High Temperature. New York: John Wiley and Sons, 1980.
[36] Du H D. Ecological Responses of Foliar Anatomical Structure and Physiological Characteristics of Dominant Plants at Different Site Conditions in North Shanxi Loss Plateau. Yangling: North West Agriculture and Forestry University, 2010.
杜华栋. 陕北黄土高原优势物种叶片结构与生理特性不同立地环境的生态响应. 杨凌: 西北农林科技大学, 2010.
[37] Sairam R K, Vasanthan B, Ajay A. Calcium regulates Gladiolus flower senescence by influencing anti-oxidative enzymes activity. Acta Physiologiae Plantarum, 2011, 33: 1897-1904.
[38] Du H D. Study on Adaptations of Dominant Plants to Different Soil Erosion Environments in Hill-gully Region of Loess Plateau——from Community, Individual, Tissue to Physiology. Beijing: The University of Chinese Academy of Sciences, 2013.
杜华栋. 黄土丘陵沟壑优势植物对不同侵蚀环境的适应研究——从群落、个体、组织到生理. 北京: 中国科学院大学, 2013.
[39] Hu S, Jiao J Y, Du H D,etal. Antioxidant properties of plants in the hilly-gullied loess plateau. Acta Prataculturae Sinica, 2014, 23(5): 1-12.
胡澍, 焦菊英, 杜华栋, 等. 黄土丘陵沟壑区不同立地环境下植物的抗氧化特性. 草业学报, 2014, 23(5): 1-12.
[40] Hu S. Antioxidant Properties and Osmotic Adjustments of Plants in the Hilly-Gullied Loess Plateau. Yangling: North West Agriculture and Forestry University, 2014.
胡澍. 黄土丘陵沟壑区植物抗氧化特性和渗透调节作用研究. 杨凌: 西北农林科技大学, 2014.
[41] Cui X Y, Chen Z Y, Zhang M S,etal. Effect of soil pH on physiological-biochemical characteristics in flue-cured tobacco (NicotianatabacumL.) leaves. Plant Physiology Communications, 2005, 41(6): 737-740.
崔喜艳, 陈展宇, 张美善, 等. 土壤 pH 值对烤烟叶片生理生化特性的影响. 植物生理学通讯, 2005, 41(6): 737-740.
[42] Tang L, Shi W M, Wang X C. Cloning of potassium transport genes in higher plants, and crop molecular breeding. Plant Nutrition and Fertilizer Science, 2001, 7(4): 467-473.
汤利, 施卫明, 王校常. 植物钾吸收转运基因的克隆与作物遗传改良. 植物营养与肥料学报, 2001, 7(4): 467-473.
[43] Souza R P, Machado E C, Silva J A B,etal. Photosynthetic gas exchange, chlorophyll fluorescence and some associated metabolic changes in cowpea (Vignaunguiculata) during water stress and recovery. Environmental and Experimental Botany, 2004, 51(1): 45-56.
[44] Pan X, Qiu Q, Li J Y,etal. Physiological indexes of six plant species from the Tibetan plateau under drought stress. Acta Ecologica Sinica, 2014, 34(13): 3558-3567.
潘昕, 邱权, 李吉跃, 等. 干旱胁迫对青藏高原 6 种植物生理指标的影响. 生态学报, 2014, 34(13): 3558-3567.
[45] Zhu J J, Zhang J L, Liu H C,etal. Photosynthesis non-photochemical path ways and activities of antioxidant enzymes in a resilient ever green oak under different climatic conditions from a valley-savanna in Southwest China. Physiologia Plantarum, 2009, 135: 62-72.
[46] Liang X. The Study on Physiological and Ecological Adaption of Typical Plant in Karst Rocky Desertification of Heqing, Yunnan. Guilin: Guangxi Normal University, 2011.
梁晓. 云南鹤庆县石漠化地区典型植物生理生态适应性研究. 桂林: 广西师范大学, 2011.
猜你喜欢
热带作物学报(2022年8期)2022-09-16
落叶果树(2021年6期)2021-02-12
中国农村水利水电(2018年12期)2018-12-28
幸福·婚姻版(2017年9期)2017-09-22
中国水土保持(2017年6期)2017-01-21
中国水土保持科学(2016年4期)2016-09-21
体育科技(2016年2期)2016-02-28
西华师范大学学报(自然科学版)(2015年3期)2015-02-27
应用海洋学学报(2014年2期)2014-11-26
植物营养与肥料学报(2012年1期)2012-10-26
