
利用qPCR法检测OsPIMT1转基因水稻中的外源片段拷贝数
2018-01-25 08:07魏毅东罗曦吴方喜林悦龙何炜连玲谢华安张建福
福建稻麦科技 2017年4期
魏毅东,罗曦,吴方喜,林悦龙,何炜,连玲,谢华安*,张建福*
(1.福建省农业科学院水稻研究所,福建福州350019;2.农业部华南杂交水稻种质创新与分子育种重点实验室/福州(国家)水稻改良分中心/福建省作物种质创新与分子育种省部共建国家重点实验室培育基地/杂交水稻国家重点实验室华南研究基地/福建省作物分子育种工程实验室/福建省水稻分子育种重点实验室,福建福州350003)
植物转化已经广泛应用于植物功能基因研究与作物品种改良。目前常用的农杆菌介导的转化法,由于片段是随机插入到基因组中,当多个片段重组到基因组中将影响外源基因的表达,甚至会造成外源基因沉默[1],且不利于后续的遗传分析。为了获得单一插入片段的转基因植株,早期的研究人员不得不依赖于费时费力,且需要大量基因组DNA的Southern 杂交法进行拷贝数鉴定。随着实时荧光定量 PCR(Real Time Fluorescence Quantitative PCR,qPCR)技术的不断发展,其已经成为快速、灵敏、高效的拷贝数鉴定方法。目前,qPCR技术已经被用于转基因棉花,小麦,玉米等作物的拷贝数鉴定[2-4]。本研究选取保守的单拷贝基因甘油醛-3-磷酸脱氢酶GAPDH2作为内参基因[5,6],利用基于2-ΔCT相对定量法的荧光定量PCR技术,建立了快速,高效的不依赖于标准曲线的水稻除草剂草铵膦抗性基因的拷贝数分析体系,从而简化了转基因水稻拷贝数鉴定的流程并提高了检测通量。
1 材料与方法
1.1 材料
质粒模版:pOE-PIMT1(含有草铵膦抗性基因bar),GAPDH2基因PCR产物和UBQ5基因PCR产物。
水稻材料:籼稻航2号对照,含有bar基因的OsPIMT1转基因克隆,依次命名为AL1~AL9。
1.2 引物设计与合成
水稻内参基因UBQ5,GAPDH2和抗性基因bar的引物使用Primer premier5软件设计,由铂尚生物技术(上海)有限公司合成。
bar基因:
上游引物:GCACCATCGTCAACCACTAC,
下游引物GTCGTCCGTCCACTCCTG;
GAPDH2基因:
上游引物:AAGCCAGCATCCTATGATCAGATT,
下游引物:
CGTAACCCAGAATACCCTTGAGTTT;
UBQ5基因:
上游引物:ACCACTTCGACCGCCACTACT;
下游引物:ACGCCTAAGCCTGCTGGTT。
1.3 基因组DNA提取
采用CTAB法提取水稻叶片DNA[7],对提取的DNA进行琼脂糖电泳以及紫外分光法检测,保证DNA未降解且无杂质污染。
1.4 荧光定量PCR 扩增
实时定量PCR反应在96孔板中进行,50 μL反应体系包含以下成分:25μL 2× FastStart Universal SYBR Green Master (ROX),上下游引物终浓度各为0.3μM,5μL稀释的DNA(50 ng/μL)。反应条件:95℃预变性10 min;95℃变性15 s,60℃退火延伸60 s,40个循环。反应结束后根据ABI7500仪器标准程序进行溶解曲线分析。
1.5 内参基因ubc5和外源基因bar标准曲线制作
以含有草铵膦抗性基因bar的质粒pOE-PIMT1作为标准品,将其进行5×梯度稀释,浓度分别为10 pg/μL,2 pg/μL,0.4 pg/μL, 80 fg/μL,16 fg/μL。同时以GAPDH2和UBQ5基因的PCR产物为标准品,稀释终浓度为0.5 pg/μL,0.1 pg/μL,20 f/μL,4 pg/μL,0.8 fg/μL。
反应扩增程序按1.3进行,最后分别制作标准曲线。
1.6 转基因水稻中内参基因和外源基因的定量分析
利用已验证的含bar基因的纯合单拷贝株系的gDNA用作参比模版,9个PCR阳性的OsPIMT1转基因克隆植株以及亲本航2号的gDNA为待测样品模板,以内参基因与抗性基因的检测引物进行qPCR 检测,所得CT值利用2-△△CT法进行定量分析。
1.7 转基因植株的Southern杂交分析
将提取的水稻gDNA利用PstI(Takara)进行酶切(时间6 h,温度37℃,体积250 μL,gDNA 150 μg)。待酶切结束后,在酶切体系中加入2.5倍体积冰乙醇和100 μL体积3M NaCl溶液混合后-20℃放置30 min,于16 000 g低温离心10 min。吸弃去上清后,加入55 μL ddH2O溶解DNA,将溶解的DNA与6μL 10× DNA loading buffer混合后上样电泳。电泳参数为:0.7%琼脂糖,2.5 V/cm电泳8 h左右,直至溴酚蓝指示带跑至底部位置。参照分子克隆第三版[8],采用毛细管向上转膜法进行DNA印迹,杂交、探针制备和显色使用AlkPhos Direct Labeling and Detection System (Amersham)进行,操作步骤参照厂家说明书。
1.8 转基因后代的分离比分析
籼稻航2号种子和T1代转基因种子经过1 d浸种后,用湿布包裹于37℃催芽。将发芽的种子播种到盆土中,待第2片叶完全展开后喷施500 mg/mL草铵膦,5 d后将幼苗取出,统计死亡和存活的幼苗数量,所得数据用SPSS19进行卡方统计分析。
2 结果与分析
2.1 荧光定量扩增曲线与溶解曲线
3个基因bar,UBQ5和GAPDH2的扩增曲线如图1所示,结果表明扩增曲线平滑,扩增稳定。同时我们对扩增产物进行溶解曲线测定,bar、UBQ5和GAPDH2基因的Tm值分别为81.8℃,80.5℃,和78.3℃,溶解曲线均为单一峰,说明引物的特异性较强,在反应中无引物二聚体等非特异性扩增,适合用于qPCR分析。

图1 bar, UBQ5, GAPDH2 基因扩增曲线与溶解曲线
2.2 荧光定量标准曲线
以起始模板拷贝数的对数为纵坐标,CT值为横坐标绘制标准曲线,分别获得bar,UBQ5和GAPDH2的标准曲线(图2)。由于2-ΔΔCT对两个基因的扩增效率要求很高,需要扩增效率一致,而bar基因引物和GAPDH2引物扩增效率相近,约95%,符合实验要求,因此选取GAPDH2这个内参基因用于后续的拷贝数分析。

图2 内参基因UBQ5和GAPDH2与外源抗性基因bar的标准曲线
2.3 基于2-ΔΔCT鉴定T0代转基因水稻的拷贝数
将所提DNA用于荧光定量PCR分析,由于待测样品为T0代植株,因此其插入位点为杂合型,纯合单拷贝对照样品的RQ值应为T0代单拷贝植株RQ值的2倍,而与双拷贝植株的RQ值相等。通过2-△△CT法将所有样品与纯合单拷贝的参比对照进行比较,结果表明L1,L2,L4,L5,L6,L8为单拷贝植株,而L3,L7,L9为双拷贝植株,其余植株为单拷贝转基因植株(图3)。
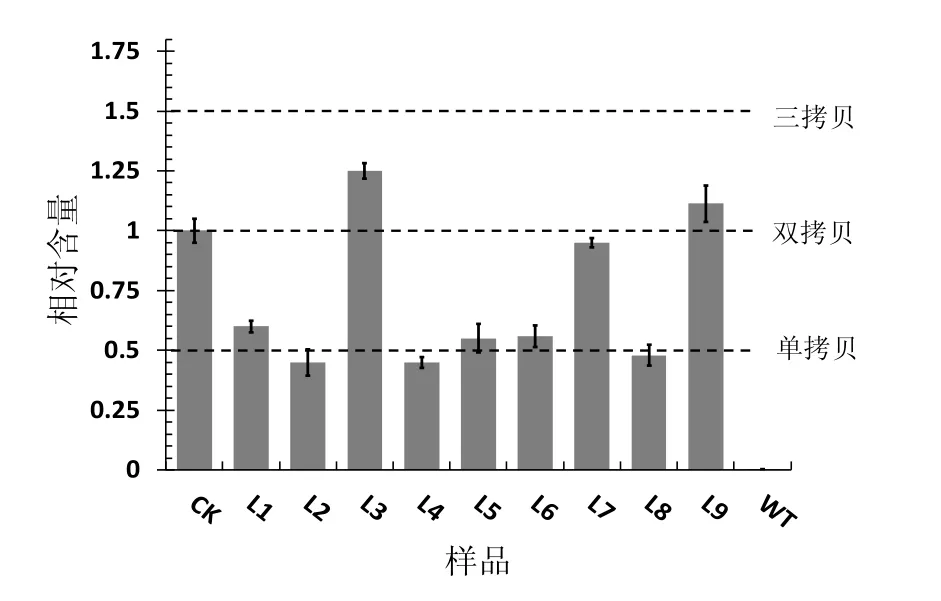
图3 荧光定量鉴定转基因材料拷贝数
2.4 转基因水稻southern杂交验证与分离比分析
从9个转基因克隆中选取5个转基因植株进行Southern杂交拷贝数验证,结果表明L1,L2,L4和L5为单拷贝植株,而L3为双拷贝植株(图4),这与qPCR结果完全一致。为了进一步验证qPCR所得的拷贝数结果,对T1代转基因进行草铵膦筛选,阴性与阳性植株数量进行卡方分析,单拷贝植株符合1/3分离比,双拷贝植株符合1/15分离比(表1)。
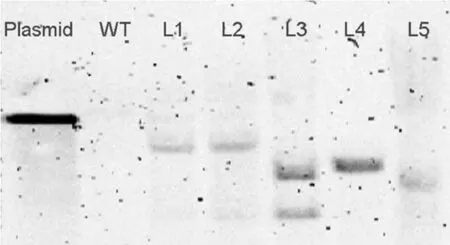
图4 转基因植株的Southern杂交检测

表1 转基因水稻后代分离比分析
3 讨论与结论
早期的水稻转化过程中,由于当时基因组测序尚未完成且水稻组培转化的难度较大,需要Southern杂交鉴定转化事件和拷贝数分析。随着基因组测序的技术的快速发展,许多植物,包含水稻的全基因组序列都已测序完成,内源基因的拷贝数以及序列结构都已十分明确。利用荧光定量PCR技术,借助于基因组数据选取合适的内参基因能够准确确定转基因作物对的拷贝数。近年的研究结果表明,Southern杂交所得的拷贝数并不准确[9-11]。排除Southern杂交实验误差,如Southern杂交中酶切不完全,不同片段转膜效率不同等因素;农杆菌介导的水稻转化本身存在不确定性,会发生大量的基因重排或片段重组[12],可能导致插入位置附近区域大片段的复制或部分重组的发生,因此需要其他方法进一步验证Southern blot拷贝数[13]。
此外,由于本研究采用2-△△CT法而不依赖于非标准曲线法,提高了检测效率和通量,但应尽可能排除样品方面的影响,需要样品纯度较高,扩增体系要相对稳定,重复性好。经过2-△△CT法计算,单拷贝,双拷贝和三拷贝的RQ值应该分别为0.5,1.0以及1.5。实际定量过程中,一般单拷贝RQ值在0.25~0.75,双拷贝RQ值在0.75~1.25,三拷贝为1.25~1.75。因此,对于每个样品的CT值误差应当控制在0.25以内,否则结果难以确定。虽然本研究选用了bar基因进行检测,但只要更换外源基因引物即可检测潮霉素抗性基因HPTII,新霉素抗性基因NPTII等多种抗性基因。
[1] Stam,M.,J.N.Mol,J.M. Kooter, Review article:the silence of genes in transgenic plants[J].Annals of Botany,1997,79(1):3-12.
[2] 李淑洁,张正英.REAL-TIME PCR方法测定转基因小麦中外源基因拷贝数[J].中国生物工程杂志,2010,30(3): 90-94.
[3] 杨晓杰,刘传亮,张朝军,等.不同转化方法获得的转基因棉花外源基因拷贝数分析[J].农业生物技术学报,2011,19(2):221-229.
[4] Shepherd,C.T.,A.N.M. Lauter,M.P. Scott,Determination of transgene copy number by real-time quantitative PCR[J]. Transgenic Maize: Methods and Protocols,2009 :129-134.
[5] 魏毅东,陈玉,郭海萍,等.水稻缺素胁迫下实时荧光定量 RT-PCR 的内参基因的选择[J].农业生物技术学报,2013,21(11):1302-1312.
[6] Jain,M.,A. Nijhawan,A.K.Tyagi,et al. Validation of housekeeping genes as internal control for studying gene expression in rice by quantitative real-time PCR[J].Biochemical and biophysical research communications,2006,345(2): 646-651.
[7] Porebski,S.,L.G.Bailey, B.R. Baum,Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components[J].Plant molecular biology reporter, 1997,15(1): 8-15.
[8] DY, J. and M. Li,Molecular Cloning-A Laboratory Manual(分子克隆实验指南). 2002,2nd,Beijing: Science Press.
[9] Ingham,D.J.,S. Beer,S. Money, et al. Quantitative real-time PCR assay for determining transgene copy number in transformed plants[J].Biotechniques, 2001,31(1):132-141.
[10] Livak, K.J.,T.D. Schmittgen, Analysis of relative gene expression data using real-time quantitative PCR and the 2- ΔΔCT method[J].methods, 2001,25(4): 402-408.
[11] Faize,M,L. Faize, L. Burgos, Using quantitative realtime PCR to detect chimeras in transgenic tobacco and apricot and to monitor their dissociation[J]. BMC Biotechnol, 2010,10:53.
[12] Jeon,J.S,S. Lee, K.H. Jung, et al. T‐DNA insertional mutagenesis for functional genomics in rice[J].The Plant Journal,2000,22(6): 561-570.
[13] Mason,G.,P. Provero, A.M. Vaira, et al. Estimating the number of integrations in transformed plants by quantitative real-time PCR[J].BMC biotechnology,2002,2(1):20.
猜你喜欢
学与玩(2022年10期)2022-11-23
今日农业(2022年3期)2022-06-05
河北医学(2021年10期)2021-10-27
中国临床医学影像杂志(2019年6期)2019-08-27
中国生殖健康(2018年1期)2018-11-06
今日中国·中文版(2017年8期)2017-09-03
中华实验和临床病毒学杂志(2016年3期)2016-08-09
创新科技(2015年1期)2015-12-24
医学综述(2014年24期)2014-03-08
中国科技信息(2011年12期)2011-02-17
