
动植物蛋白源及牛磺酸对大菱鲆摄食、生长及体组成的影响❋
2018-03-16 03:25刘兴旺麦康森刘付志国艾庆辉
中国海洋大学学报(自然科学版) 2018年5期
刘兴旺, 麦康森, 刘付志国, 艾庆辉❋❋
(1.教育部海水养殖重点实验室(中国海洋大学),山东 青岛266003;2. 盐城恒兴饲料有限公司,江苏 盐城 224111)
近年来,以植物蛋白源作为鱼粉的替代物在肉食性鱼类中使用已成为水产动物营养研究的热点问题[1]。然而,氨基酸组成不平衡、适口性差、抗营养因子等因素限制了植物蛋白在肉食性鱼类中的用量[2]。最近研究发现,植物蛋白中缺乏牛磺酸可能也是限制其在肉食性鱼类饲料中使用量的重要因素[3]。牛磺酸(Taurine)又称牛胆碱、牛胆酸或牛胆素,是一种小分子β-含硫氨基酸。鱼类合成牛磺酸的能力因种类而不同,也可能与鱼的发育阶段相关[4]。高植物蛋白条件下,饲料中补充牛磺酸可促进鱼类生长、提高饲料转化,改善海水鱼“绿肝”等现象[5-7]。因此,牛磺酸在一定条件下是某些海水鱼的必需营养素[8]。已有研究表明,大菱鲆仔稚鱼的生长与饵料中牛磺酸含量正相关[9]。且在高植物蛋白水平条件下添加牛磺酸能够改善大菱鲆(Scophthalmusmaximus)生长性能并调控牛磺酸转运载体mRNA的表达[10]。本研究的目的:在以鱼粉或大豆浓缩蛋白为主要蛋白源的情况下,比较饲料中添加不同含量牛磺酸对大菱鲆幼鱼摄食、生长及脂肪代谢的影响,以期为牛磺酸在大菱鲆配合饲料中的合理应用提供更多理论支持。
1 材料与方法
1.1 实验饲料
分别以鱼粉(FM,粗蛋白74.3%)和大豆浓缩蛋白(SPC,粗蛋白70.1%)作为蛋白源,配制8种等氮等能的饲料(分别标记为T1~T8)。T1~T4主要蛋白源为FM,T5~T8用57%的SPC替代相应蛋白的FM,在2种蛋白源饲料中分别添加0%、0.5%、1.0%和2.0%的牛磺酸。实验饲料配方及化学成分见表1。所有原料经粉碎过80目筛,按表1配方混合均匀,然后再加入鱼油和大豆卵磷脂(先溶解于鱼油中),手工将油脂微小颗粒搓散,于V型立式混合机(上海天祥V-0.5型)中混合均匀,最后加入约30%蒸馏水形成硬团,全自动鱼用饵料机(260,山东威海友谊机械厂)中将饲料挤压成直径3.0 mm的颗粒,在50℃恒温下干燥12 h,干燥颗粒置-20℃冰箱备用。

表1 试验饲料配方和化学成分(干重)分析Table 1 Formulation and proximate composition (dry weight) of eight experimental diets /%
1.鱼粉购自七好生物科技有限公司(中国山东),粗蛋白74.3%,粗脂肪7.3%;大豆浓缩蛋白由山东东营万得福植物蛋白生物科技有限公司提供,粗蛋白70.1%;粗脂肪0.8%。Fish meal, obtained from Qihao Bio-tech Company (Shandong, China), crude protein, 74.3% dry matter, crude lipid 7.3% dry matter;SPC supplied by Dong Ying Wonderful Vegetable Protein Science and Technology Co., Ltd. (Shandong, China), crude protein, 70.1% dry matter, crude lipid, 0.8% dry matter.
2.维生素预混料(mg或IU/kg饲料):维生素A醋酸酯,16 000IU;维生素D3,2 500IU;维生素E醋酸酯,200mg;维生素K3(MSB),5.1mg;维生素B1,24.5mg;核黄素,36mg;泛酸钙,58.8mg;维生素B6,19.8mg;维生素B12,0.1mg;烟酸,198mg;叶酸,19.6mg;生物素,1.2mg;肌醇,784mg。Vitamin premix (mg or IU/kg diet): retinol acetate, 16 000IU; cholecalciferol, 2 500IU; alpha-tocopherol, 200 mg; vitamin K3, 5.1 mg; thiamin 24.5 mg; riboflavin, 36 mg; pantothenic acid, 58.8 mg; pyridoxine HCl, 19.8 mg; vitamin B12, 0.1 mg; niacin acid, 198 mg; folic acid, 19.6 mg; biotin, 1.20 mg; inositol, 784 mg.
3.矿物质预混料(mg/kg饲料):MgSO4·7H2O,1 200mg;CuSO4·5H2O,10mg;ZnSO4·H2O,50mg;FeSO4·H2O,80mg;MnSO4·H2O,45mg;CoCl(1%),50mg;Na2SeO3(1%),20mg;Ca(IO3)2(1%),60mg;沸石粉,8 485mg。Mineral premix (mg/kg diet): MgSO4·7H2O,1 200mg;CuSO4·5H2O,10mg;ZnSO4·H2O,50mg;FeSO4·H2O,80mg;MnSO4·H2O,45mg;CoCl(1%),50mg;Na2SeO3(1%),20mg;Ca(IO3)2(1%),60mg;Zoelite, 8 485mg.
4.牛磺酸:湖北永安集团潜江永安药业股份有限公司生产。Supplied by Hubei Qianjiang Pharmaceutical Co., Ltd.
1.2 养殖管理
大菱鲆幼鱼购自胶南市大珠山镇古镇营村大菱鲆育苗场,为当年孵化同批鱼苗。养殖实验在中国海洋大学鳌山卫实验基地进行。实验开始前大菱鲆禁食24 h,选择大小均匀((38.19±0.09)g)、体格健壮的幼鱼随机分配于24个养殖桶(每种饲料随机投喂3组),每桶18尾。每天早晚投饵2次(08:30,18:30),达饱食水平,投喂结束后30min,吸出残饵及粪便。实验采用养殖桶(总体积400 L,水体体积300 L)均为玻璃纤维桶。养殖所用海水为沙滤水,采用循环流水系统,每桶进水量为1.0 L/min。实验期间水温控制为12.0~18.0°C,盐度为28.5~32,溶氧维持在7mg/L左右。试验共进行8周。
1.3 样品收集及化学分析
养殖试验结束后,停止喂食24 h,开始收集大菱鲆。分别称量每桶鱼体总重,记录鱼体个数,计算每桶大菱鲆平均体重。每桶取5~7尾鱼用于体常规分析。此后,每桶随机抽取6尾大菱鲆,分别测定其体重、体长、内脏和肝脏重量,以计算肥满度、脏体比和肝体比。同时以注射器自尾静脉取血,离心后分离血清并于-75℃保存。
鱼体和饲料常规成分分析均采用AOAC的方法[12]。水分的测定为105℃烘干恒重法(24 h);粗蛋白的测定为凯氏定氮法,采用瑞典TECATOR公司1030型蛋白质自动分析仪;粗脂肪的测定为索氏抽提法;粗灰分的测定为箱式电阻炉550℃灼烧法(16 h)。总能采用氧弹式热量计(Parr 1281,美国)测定。甘油三酯和总胆固醇含量采用试剂盒测定(南京建成)。
1.4 计算与统计分析
大菱鲆特定生长率、摄食率、蛋白质效率和饲料效率参照以下公式计算:
特定生长率(Specific growth rate, SGR, %/d)=100×[ln (终末体重)-ln (初始体重)]/实验天数;
成活率(Survival rate, SR, %)=100×实验结束每桶鱼尾数/实验开始每桶鱼尾数;
摄食率(Feed intake, FI, %/d)=100×每尾鱼摄食的饲料干物质重/[(实验结束时鱼体重+实验开始时鱼体重)/2×养殖实验天数];
饲料效率(Feed efficiency, FE)=(实验结束时鱼体重-实验开始时鱼体重)/每尾鱼摄食的饲料干物质重;
蛋白效率比(Protein efficiency ratio, PER)=(终末体重-初始体重)/蛋白摄入量;
肥满度(Condition factor, CF, g/cm3)=100×实验终体重(g)/(鱼体长度)3(cm3);
肝指数(Hepatosomatic index, HSI, %)=100×肝脏重(g)/鱼体重(g);
内脏指数(Viscerosomatic index, VSI, %)=100×内脏团重(g)/鱼体重(g);
所有百分率在分析统计前均先经Arcsine转换。试验结果采用单因素方差分析(ANOVA)与双因素方差分析同时进行,当单因素方差分析处理之间差异显著(P< 0.05)时,用Tukey’s检验进行多重比较分析。所有统计分析均用SPSS 11.0软件进行。
2 结 果
2.1 生长及饲料利用率
各处理组成活率在92.59%~98.15%之间,且差异不显著(P>0.05)(见表2)。饲料中不同的蛋白源1.S.E.M. 标准误Standard error of means。
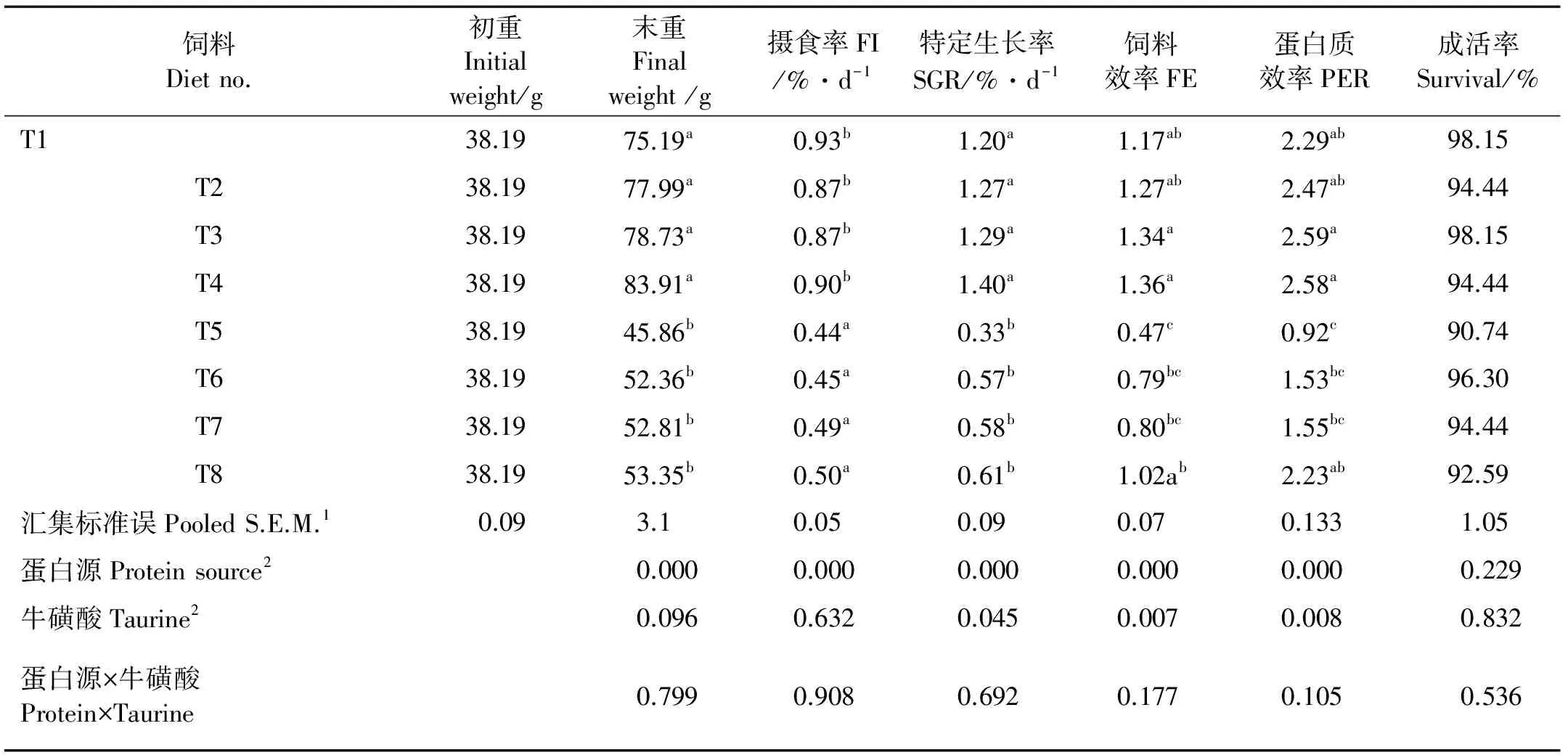
表2 不同基础蛋白源饲料中添加牛磺酸对大菱鲆摄食和生长性能的影响Table 2 Growth performance of Turbot fed various protein sources-based diets with or without supplemental taurine
注:表中数据为平均值(n=3),同一列右上角不同英文上标字母表示有显著差异(P< 0.05)。
Note: Values are presented as means of triplication. Means in the same column with different superscripts are significantly different from each other (P< 0.05).
2.分别采用单因素方差分析或双因素方差分析。One-way or two-way analysis of variance.
极显著(P<0.001)得影响了大菱鲆的末重、FI、SGR、FE和PER。与鱼粉组相比,SPC组末重、FI、SGR、FE和PER均显著下降(P<0.001)。饲料中添加牛磺酸对大菱鲆末重和FI没有显著影响(P>0.05),却显著影响了SGR、FE和PER。随着饲料中牛磺酸的添加,投喂2种蛋白源饲料大菱鲆SGR、FE和PER均显著升高(P<0.05)。饲料蛋白源和牛磺酸对大菱鲆生长及饲料利用未发现有交互作用(P>0.05)。
2.2 鱼体生化组成
不同蛋白源及牛磺酸水平对大菱鲆体组成的影响见表3。饲料蛋白源极显著影响了大菱鲆鱼体水分、粗蛋白及粗脂肪含量(P<0.001),对灰分无显著影响(P>0.05)。与鱼粉组相比,SPC组鱼体水分升高、粗蛋白和粗脂肪降低。而饲料中添加牛磺酸对大菱鲆鱼体水分及粗蛋白含量有显著影响,随牛磺酸添加量升高,鱼体水分有降低的趋势,而粗蛋白有升高的趋势(P<0.05),但牛磺酸对鱼体粗脂肪和灰分无显著影响。不同蛋白源和牛磺酸对大菱鲆鱼体组成的影响不存在交互作用(P>0.05)。
注:表中数据为平均值(n=3),同一列右上角不同英文上标字母表示有显著差异(P< 0.05)。
Note: Values are presented as means of triplication. Means in the same column with different superscripts are significantly different from each other (P<0.05).
1.S.E.M. 标准误Standard error of means。
2.分别采用单因素方差分析或双因素方差分析。One-way or two-way analysis of variance.
2.3 肥满度、脏体比及血清生化指标
饲料中不同蛋白源和添加牛磺酸对大菱鲆肥满度和内脏指数无显著影响(P>0.05)(见表4)。与鱼粉组相比,SPC组大菱鲆肝体比显著降低(P<0.05)。SPC组肝体比有随着牛磺酸添加而逐渐升高的趋势,但差异不显著(P>0.05)。与鱼粉组相比,SPC组大菱鲆血清总胆固醇含量显著降低(P<0.001),但饲料中添加牛磺酸能显著提高SPC组血清总胆固醇水平(P<0.05)。SPC组大菱鲆与鱼粉组相比血清甘油三酯含量(P<0.01)显著降低,但随饲料中牛磺酸的添加,2种蛋白源处理组大菱鲆血清甘油三酯含量均有升高的趋势,但差异不显著(P>0.05)。对血清甘油三酯含量来讲,蛋白源与牛磺酸之间有一定的交互作用(P<0.05)。
3 讨论
饲料中高水平植物蛋白源与鱼粉相比显著抑制了大菱鲆的摄食率、特定生长率、饲料效率及蛋白质效率(P<0.05)(见表2),该结果与之前的研究结果相似[13]。Deng等[14]在牙鲆(Paralichthysolivaceus)、Kissil等[15]在乌颊鱼(Sparusaurata)上的研究也发现随着饲料中大豆蛋白添加量升高,实验鱼生长率及饲料转化率呈直线下降。Day等[16]的研究却发现,饲料中SPC能够替代25%的鱼粉蛋白而对大菱鲆摄食、生长、FE和PER不造成显著影响。该研究结果与前述研究的差异可能与其基础饲料中较高的脂肪水平(22.7%)有关。在牙鲆上的研究显示,高水平的植物蛋白替代鱼粉会对鱼类的脂肪代谢造成显著影响[17]。因此,通过添加牛磺酸改善因植物蛋白替代鱼粉导致造成的脂肪代谢异常问题成为改善植物蛋白在鱼类中应用的一个重要手段[18]。
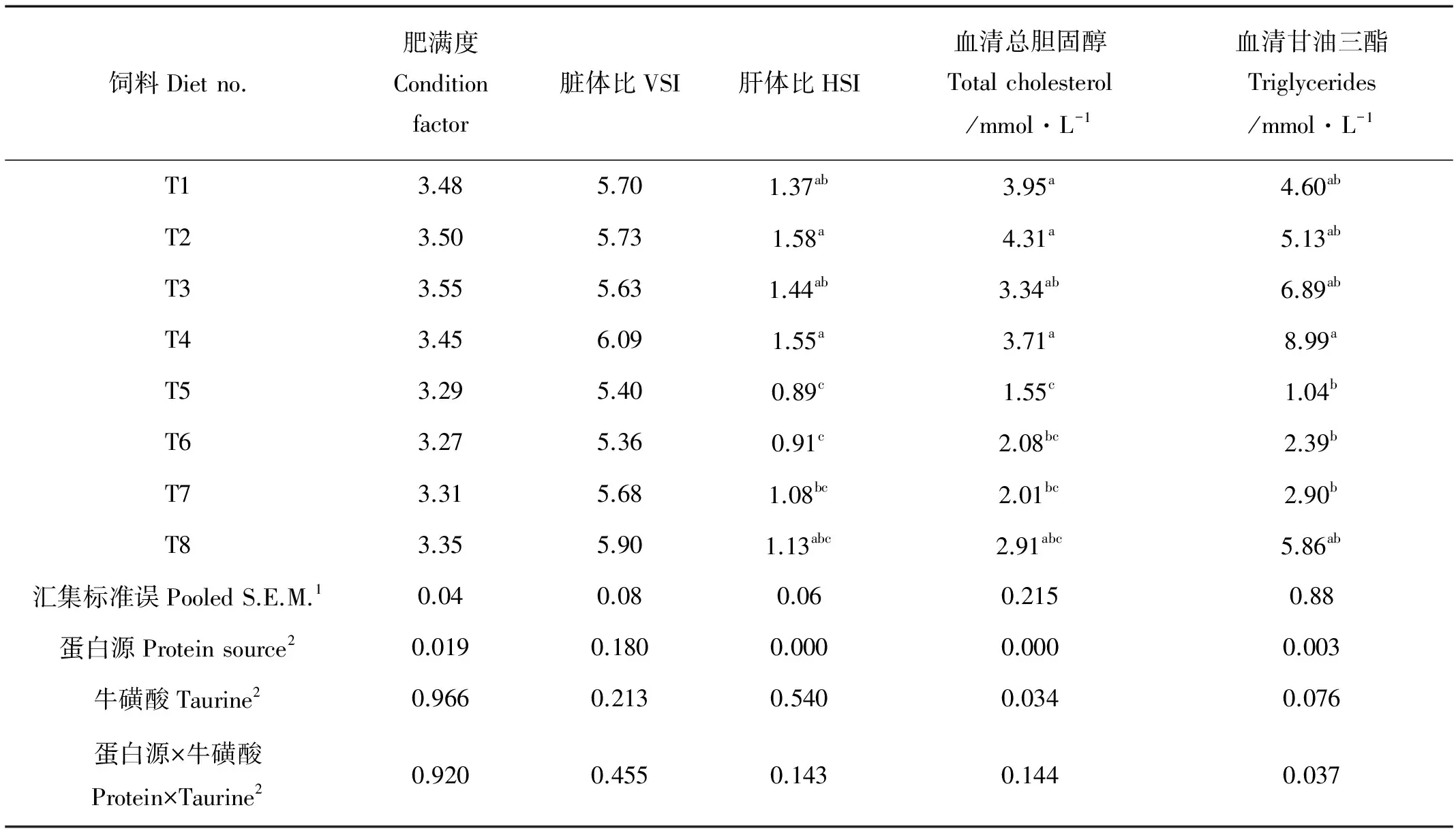
表4 不同基础蛋白源饲料中添加牛磺酸对相关生理指标的影响Table 4 Hematocrit and biometric indices of Turbot fed various protein sources-based diets with or without supplemental taurine
注:表中数据为平均值(n=3),同一列右上角不同英文上标字母表示有显著差异(P< 0.05)。
Note: Values are presented as means of triplication. Means in the same column with different superscripts are significantly different from each other (P< 0.05).
1.S.E.M. 标准误Standard error of means。
2.分别采用单因素方差分析或双因素方差分析。One-way or two-way analysis of variance.
本研究中,随着饲料中牛磺酸梯度升高,大菱鲆特定生长率、饲料效率及蛋白质效率显著升高(P<0.05)。在牙鲆[4]、虹鳟(Oncorhynchusmykiss)[19]、欧洲鲈鱼(Dicentrarchuslabrax)[20]和五条鰤(Seriolaquinqueradiata)[21]等鱼类上的研究也得到类似结果。这一方面与牛磺酸调节消化酶活性促进对营养物质的消化吸收有关[10],也与牛磺酸的添加节约了从其他含硫氨基酸向牛磺酸的合成转化有关[22]。Wang等研究发现大菱鲆具有将蛋氨酸和胱氨酸转化为牛磺酸的能力,但是这种能力不足以满足大菱鲆快速生长的需要,需要在饲料中补充牛磺酸以促进鱼类的生长[22]。齐国山曾研究发现初始体重6.3g的大菱鲆幼鱼饲料中牛磺酸最适需求量为1.0%[23]。但在五条鰤上的研究发现,在蛋白源为SPC时实验鱼对牛磺酸的需求量高于饲料蛋白源为鱼粉时[24]。这是大豆蛋白影响粪便中牛磺酸和胆汁酸的量而降低鱼类血清中牛磺酸水平而造成的[19]。因此,植物蛋白对鱼类牛磺酸需求的影响有两方面:一方面是提高牛磺酸的排泄,另一方面抑制牛磺酸的合成。大菱鲆在不同生长阶段、不同基础配方条件下牛磺酸需求量是否存在变化,值得进一步进行研究。
在本研究中,SPC组大菱鲆与鱼粉组相比体组织粗蛋白和粗脂肪水平、肝体比及大菱鲆血清甘油三酯和胆固醇含量显著降低(P<0.05)。该结果与在其他鱼类上的研究结果相似[17]。很多研究发现,SPC会影响海水鱼类的脂肪代谢,一方面是由于大豆蛋白中的非淀粉多糖能够引起肠炎,导致鱼对脂肪吸收下降,从而引起鱼体脂肪含量下降[25];同时,植物蛋白源中其他非蛋白成分(如大豆低聚糖)具有降胆固醇的作用[17]。欧洲鲈鱼上的研究发现,SPC会通过影响鱼类肝脏中某些脂肪代谢相关酶而影响鱼的脂肪代谢[26]。也有学者发现SPC不仅仅通过影响脂肪酶而影响鱼类的脂肪代谢,也通过胆汁酸代谢途径影响鱼类的脂肪代谢[24,27]。因此,在SPC为基础的饲料中可以通过添加牛磺酸改善胆汁酸代谢而改善鱼类的脂肪代谢。本研究的结果显示饲料中添加不同水平牛磺酸时,肝体比、血清甘油三酯和总胆固醇含量都有随着牛磺酸水平升高而升高的趋势,说明饲料中添加牛磺酸能够明显改善SPC造成的鱼类脂肪代谢异常。最近的研究显示,饲料中牛磺酸的作用与鱼类的脂肪代谢密切相关[11],因此值得进一步的研究。
4 结论
植物蛋白替代鱼粉条件下补充牛磺酸能够促进大菱鲆生长,并对植物蛋白导致的脂肪代谢异常有一定缓解作用。
[1] Wang J, Tao Q, Wang Z, et al. Effects of fish meal replacement by soybean meal with supplementation of functional compound additives on intestinal morphology and microbiome of Japanese seabass (Lateolabraxjaponicus) [J]. Aquaculture Research, 2017, 48(5): 2186-2197.
[2] 刘运正, 何艮, 麦康森, 等. 新型复合动植物蛋白源部分替代鱼粉对大菱鲆幼鱼生长和肉质的影响[J]. 中国海洋大学学报(自然科学版), 2016(1): 33-39.
Liu Y Z, He G, Mai K S, et al. Effect of partial replacement of dietary fishmeal with an animal and plant protein mixture on the growth performance and muscle texture of juvenile turbot (ScophthalmusmaximusL. ) [J]. Periodical of Ocean University of China, 2016(1): 33-39.
[3] Lunger A N, McLean E, Gaylord T G, et al. Taurine supplementation to alternative dietary proteins used in fish meal replacement enhances growth of juvenile cobia (Rachycentroncanadum) [J]. Aquaculture, 2007, 271(1): 401-410.
[4] Kim S K, Takeuchi T, Akimoto A, et al. Effect of taurine supplemented practical diet on growth performance and taurine contents in whole body and tissues of juvenile Japanese flounderParalichthysolivaceus[J]. Fisheries Science, 2005, 71(3): 627-632.
[5] 骆艺文, 艾庆辉, 麦康森, 等. 饲料中添加牛磺酸和胆固醇对军曹鱼生长、体组成和血液指标的影响[J]. 中国海洋大学学报(自然科学版), 2013(8): 31-36.
Luo Y W, Ai Q H, Mai K S, et al. Effects of dietary taurine and cholesterol on growth performance, body composition and plasma metabolites in diets of cobia (RachycentronCanadumL. )[J]. Periodical of Ocean University of China, 2013(8): 31-36.
[6] Takagi S, Murata H, Goto T, et al. Necessity of dietary taurine supplementation for preventing green liver symptom and improving growth performance in yearling red sea breamPagrusmajorfed nonfishmeal diets based on soy protein concentrate [J]. Fisheries Science, 2010, 76(1): 119-130.
[7] Li M, Lai H, Li Q, et al. Effects of dietary taurine on growth, immunity and hyperammonemia in juvenile yellow catfishPelteobagrusfulvidracofed all-plant protein diets [J]. Aquaculture, 2016, 450: 349-355.
[8] Chatzifotis S, Polemitou I, Divanach P, et al. Effect of dietary taurine supplementation on growth performance and bile salt activated lipase activity of common dentex,Dentexdentex, fed a fish meal/soy protein concentrate-based diet [J]. Aquaculture, 2008, 275: 201-208.
[10] 柳茜, 梁萌青, 郑珂珂, 等. 牛磺酸及相关氨基酸对大菱鲆幼鱼生长性能及TauT mRNA表达的影响[J]. 水生生物学报, 2017, 41(1): 165-173.
Liu X, Liang M Q, Zheng K K, et al. Effects of dietary taurine and related amino acids on growth performance and taurine transporter (TAUT) mRNA expression of juvenile turbot (ScophthalmusmaximusL. ) [J]. Acta Hydrobiologica Sinica, 2017, 41(1): 165-173.
[11] Hoseini S M, Hosseini S A, Eskandari S, et al. Effect of dietary taurine and methionine supplementation on growth performance, body composition, taurine retention and lipid status of Persian sturgeon,Acipenserpersicus(Borodin, 1897), fed with plant-based diet[J/OL]. Aquaculture Nutrition,2017, DOI: 10. 1111/anu. 12563.http://onlinelibrary.wiley.com/doi/10.1111/anu.12563/full.
[12] Association of Official Analytical Chemists (AOAC). Official methods of analysis of AOAC international,16th edition[S]. Arlington, Virginia: AOAC International, 1995.
[13] 刘兴旺, 艾庆辉, 麦康森, 等. 大豆浓缩蛋白替代鱼粉对大菱鲆摄食、生长及体组成的影响[J]. 水产学报, 2014, 38(1): 112-119.
Liu X W, Ai Q H, Mai K K, et al. Effects of replacing fish meal with soy protein concentrate on feed intake and growth of turbot (Scophthalmusmaximus) [J]. Journal of Fisheries of China, 2014, 38(1): 91-98.
[14] Deng J, Mai K, Ai Q, et al. Effects of replacing fish meal with soy protein concentrate on feed intake and growth of juvenile Japanese flounder,Paralichthysolivaceus[J]. Aquaculture, 2006, 258(1): 503-513.
[15] Kissil G W, Lupatsch I, Higgs D A, et al. Dietary substitution of soy and rapeseed protein concentrates for fish meal, and their effects on growth and nutrient utilization in gilthead seabreamSparusaurataL [J]. Aquaculture Research, 2000, 31(7): 595-601.
[16] Day O J, Gonzalez H G P. Soybean protein concentrate as a protein source for turbotScophthalmusmaximusL [J]. Aquacture Nutrition, 2000, 6(4): 221-228.
[17] Deng J, Mai K, Ai Q, et al. Interactive effects of dietary cholesterol and protein sources on growth performance and cholesterol metabolism of Japanese flounder (Paralichthysolivaceus) [J]. Aquaculture Nutrition, 2010, 16(4): 419-429.
[18] Salze G P, Davis D A. Aurine: a critical nutrient for future fish feeds [J]. Aquaculture, 2015, 437(1): 215-229.
[19] Gaylord T G, Teague A M, Barrows F T. Taurine supplementation of all-plant protein diets for rainbow trout (Oncorhynchusmykiss) [J]. Journal of the World Aquacul-ture Society, 2006, 37(4): 509-517.
[20] Martinez J B, Chatzifotis S, Divanach P, et al. Effect of dietary taurine supplementation on growth performance and feed selection of sea bassDicentrarchuslabraxfry fed with demand-feeders [J]. Fisheries Science, 2004, 70(1): 74-79.
[21] Matsunari H, Takeuchi T, Takahashi M, et al. Effect of dietary taurine supplementation on growth performance of yellowtail juvenilesSeriolaquinqueradiata[J]. Fisheries Science, 2005, 71: 1131-1135.
[22] Wang Q, He G, Wang X, et al. Dietary sulfur amino acid modulations of taurine biosynthesis in juvenile turbot (Psettamaxima) [J]. Aquaculture, 2014, 422-423: 141-145.
[23] Qi G S, Ai Q H, Mai K S, et al . Effects of dietary taurine supplementation to a casein-based diet on growth performance and taurine distribution in two sizes of juvenile turbot (Scophthalmus maximus L.)[J]. Aquaculture, 2012(358-359): 122-128.
[24] Takagi S, Murata H, Goto T, et al. Taurine is an essential nutrient for yellowtailSeriolaquinqueradiatafed non-fish meal diets based on soy protein concentrate [J]. Aquaculture, 2008, 280: 198-205.
[25] Refstie S, Storebakken T, Baeverfjord G, et al. Longterm protein and lipid growth of Atlantic salmon (Salmosalar) fed diets with partial replacement of fish meal by soy protein products at medium or high lipid level [J]. Aquaculture, 2001, 193: 91-106.
[26] Dias J, Alvarez M J, Arzel J, et al. Dietary protein source affects lipid metabolism in the European seabass (Dicentrarchuslabrax) [J]. Comparative Biochemistry and Physiology, 2005, 142: 19-31.
[27] Yun B, Ai Q, Mai K, et al. Synergistic effects of dietary cholesterol and taurine on growth performance and cholesterol metabolism in juvenile turbot (ScophthalmusmaximusL.) fed high plant protein diets [J]. Aquaculture, 2012, 324-325: 85-91.
猜你喜欢
当代水产(2022年8期)2022-09-20
国外畜牧学(猪与禽)(2022年1期)2022-07-10
电脑报(2022年25期)2022-07-05
当代水产(2022年6期)2022-06-29
饲料工业(2021年20期)2021-11-18
饲料工业(2021年6期)2021-04-16
食品与健康(2020年7期)2020-07-09
当代水产(2019年6期)2019-07-25
当代水产(2019年1期)2019-05-16
中国水产(2019年3期)2019-03-25
