
多种神经元细胞耦合体系同步状态的最优调控
2018-03-23 03:44张健生张季谦谢朔俏黄守芳
西安文理学院学报(自然科学版) 2018年2期
张健生, 张季谦, 谢朔俏, 徐 飞, 黄守芳
(安徽师范大学 物理与电子信息学院,芜湖 241000)
近年来,耦合神经元体系的动力学行为及其调控作用研究一直以来是科研工作者关注的研究热点之一,复杂体系的同步振荡现象在物理、生物、通信网络体系中是常见的动力学行为[1-2].人们提出了许多理论模型来模拟单个以及多个神经元耦合体系的放电活动规律及其动力学调控机制[3-6].其中,针对一种北极鹅的肌肉纤维研究结果而建立了Hodgkin-Huxley(HH)模型[7],随后,将其改进为Morris-Lecar(ML)模型,并利用三维ML模型研究神经元的放电节律及其动力学行为[8].在此基础上,1961年FitzHugh和Nagumo共同提出FitzHugh-Nagumo(FHN)模型[4,9],利用这些模型,可以很方便地观察到了丰富的放电行为[10].
众所周知,在实际生物体系中,各神经元彼此之间并不是孤立存在的,而是通过合适的耦合作用构成一个有机整体,以便能有效地完成对外部各类刺激做出响应和传递信息等功能.例如,在同种单元种类构成的耦合体系中,两个ML神经元在电耦合和化学耦合下可以实现同步[11-12],延迟耦合对FHN神经元系统动力学行为有重要的调控作用[13-14].特别地,最近,人们发现,不同神经元之间也存在一定的耦合作用,它们之间协同作用可以完成一定的功能.例如,人大脑中不同神经系统之间的元同步,或者心肺神经元之间的同步,均属于不同的结构神经元系统的同步[15].本文以ML模型与FHN模型为基础,作为对比研究,一方面先考察同种神经元(例如,仅含ML神经元)体系下耦合强度对同步状态的影响;另一方面通过电耦合的方式将两种不同类型的神经元构成一种多神经元混合模型,通过理论模拟,设计适当的耦合构型,合理调节神经元的控制参量和体系的耦合参数,最终实现在两种神经元协同作用下系统达到最优同步.同时,引入同步误差对体系耦合作用结果进行对比分析,上述结果为深入理解体系的同步形成和调控机理提供一定的理论指导作用.
1 模型描述
1.1 ML神经元模型
ML神经元模型是描述北极鹅肌肉纤维电活动的模型.本文中以改进的三维ML神经元模型为研究对象,其动力学特性由下面的微分方程给出:
(1)
其中:M∞,W∞和λ(V)分别表示如下
(2)
式中:V,W,I分别表示神经元的膜电位、恢复变量、慢变调节电流;C表示膜电容,α表示时间尺度因子;gL、gCa、gK分别表示漏电流通道、钙离子通道和钾离子通道的最大电导;VL、VCa、VK分别表示漏电流通道、钙离子通道和钾离子通道的反转电压;M∞、W∞分别表示钙离子通道和钾离子通道打开的概率的稳态值;v11,v22,v33,v44分别表示M∞=0.5时候的电势、电压对M∞的斜率的倒数、W∞=0.5时候的电势、电压对W∞的斜率的倒数.其他参数取值为:VL=-0.5 mV,VK=-0.7 mV,VCa=1.0 mV,gL=0.5 mS/cm2,gK=2.0 mS/cm2,gCa=1.2 mS/cm2,v11=-0.01 mV,v22=0.15 mV,v33=0.10 mV,v44=0.05 mV,C=1.0 mF,α=0.005.详细描述可参考文献[16].
1.2 FHN神经元模型
二维的FHN神经元模型如下[17-18]:
(3)
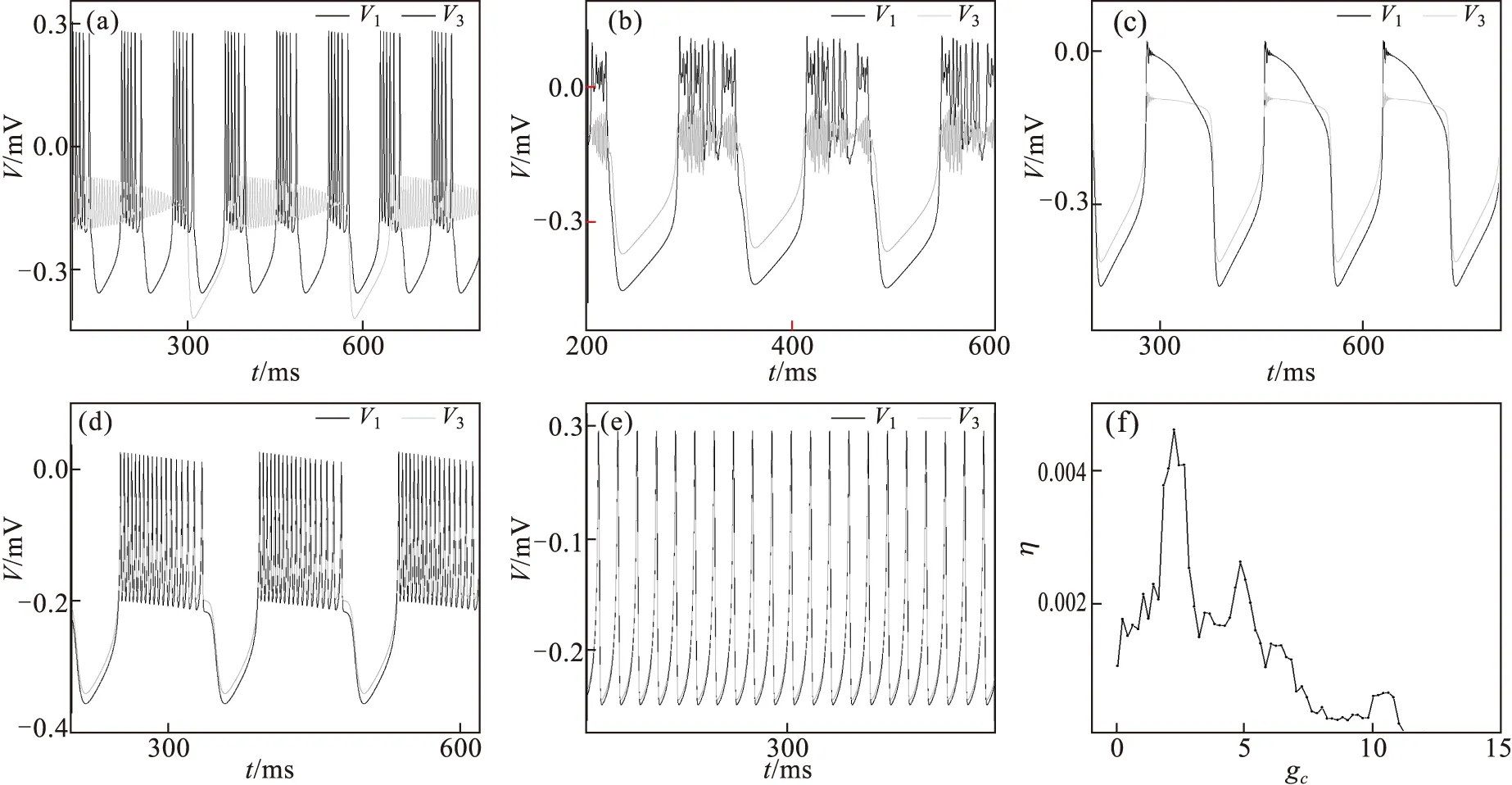
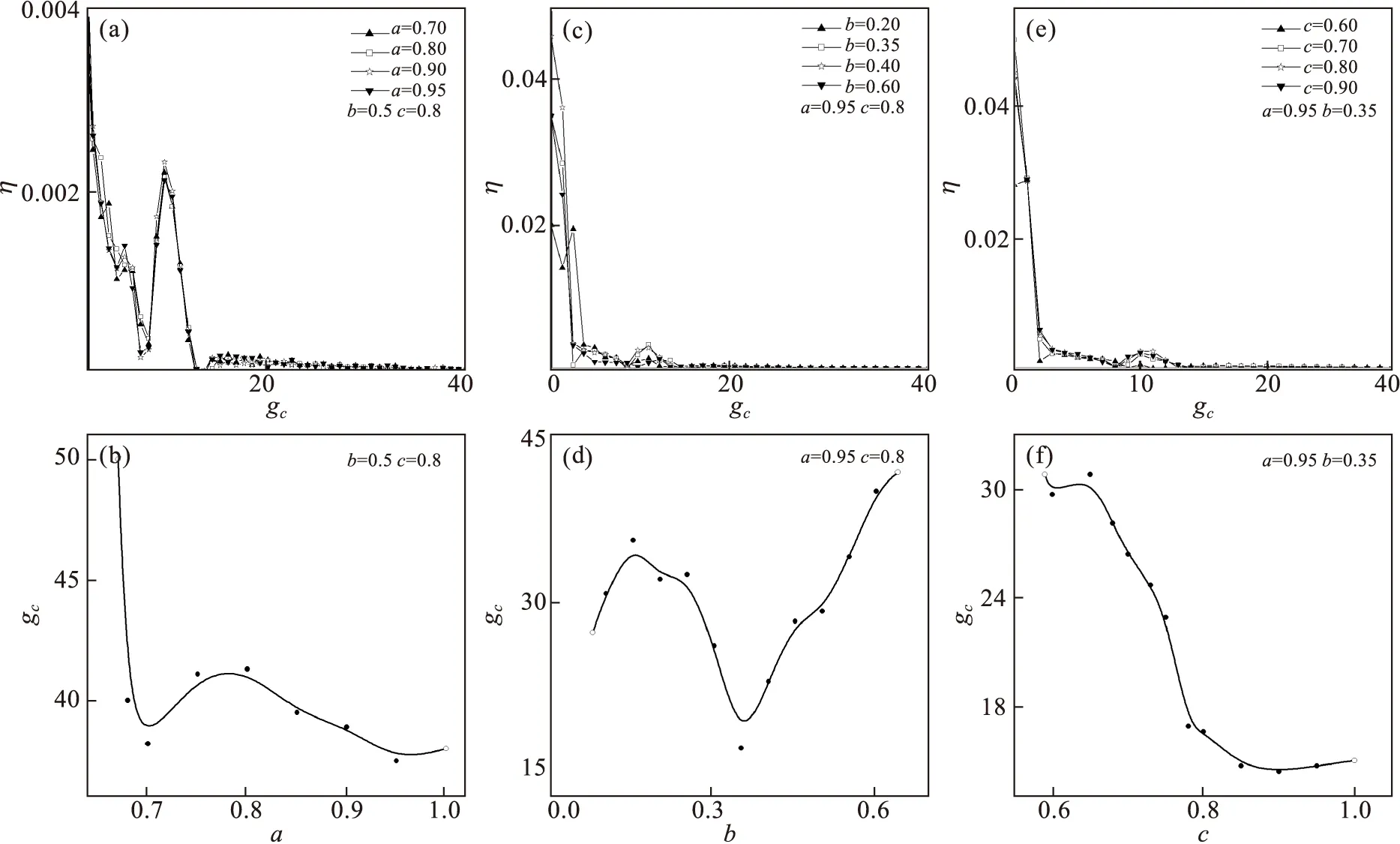
其中,V,Y分别表示神经元细胞膜电压以及恢复变量.常数a,b,c取值范围是:0 现以ML神经元为对象,研究同种类神经元耦合系统的同步行为.3个(i=1,2,3)ML神经元耦合体系的动力学行为可由如下微分方程组描述: (4) 其中:gc代表耦合强度,耦合项fi分别为f1=gc(V2-V1),f2=gc(V1+V3-2V2),f3=gc(V2-V3),其他参数代表意义和取值与式(1)、式(2)相同. 图1 不同耦合强度下两个神经元耦合同步示意图及同步误差随耦合强度变化关系图 (a)~(e):耦合强度gc=0.001,2.0,4.0,8.0,12.0下两端神经元的膜电压与时间t的峰序列;(f)同步误差随着耦合强度变化. 设置各细胞处于不同初始状态,状态参量分别取VK1=-0.7,VK2=-200,VK3=-400,同时各细胞处于不同初始点火模式.调节耦合强度,选取几个有代表性的值gc=0.001,2.0,4.0,8.0,12.0,记录首尾两个细胞在不同耦合强度下膜电压的时间序列如图1(a)~(e)所示.从图中可以看出,当耦合强度较小时,两细胞按照各自初始条件进行独立振荡,处于不同步状态,结果如图1(a)所示.随着耦合强度的增加,体系可以处于不同的同步状态,并且同步效果不断得到增强.如图1(b)中gc=2.0时开始出现混沌同步,图1(c)中出现尖峰同步,图1(d)中出现簇同步.当耦合强度进一步增大时,神经元可以达到完全同步状态,如图1(e)所示.为观察耦合体系同步效果随耦合强度变化的规律,我们引入如下同步误差来进行分析: (5) 其中:η(t)表示两神经元t时刻膜电压的均方差,〈*〉表示对观察的总时间求平均,[*]表示经过50次重复计算后取平均值.改变耦合强度,模拟计算同步时同步误差随耦合强度变化如图1(f)所示,从图中可以看出,随着耦合强度的增加,同步误差不断减小.值得注意的是由于神经元细胞膜电压会呈现出尖峰振荡、簇振荡以及混沌等不同振荡模式,因而随着耦合强度的增加会有不同的同步状态,表现在图中有多处极小值出现,如图1(f)所示.从图中可以看出,当耦合强度进一步增加超过一定值时,体系将达到完全同步状态.要使该同种细胞耦合体系处于同步状态,不管中间细胞的内部参数取值如何,细胞之间的耦合强度只需要取gc=12即可达到较好的同步效果. 在上述同种细胞耦合模型的基础上我们再引入另外一种神经元,例如FHN神经元,通过电耦合方式构建一种新的混合神经元系统模型,即ML-FHN-ML耦合体系,通过调控FHN神经元中的参数来观察系统的同步特性.ML和FHN神经元模型的动力学方程分别描述如下: (6) (7) 式(6)中:j=1,3,细胞1耦合项f1(V1,V2)=gc(V2-V1),细胞3耦合项f3(V2,V3)=gc(V2-V3),细胞2耦合项f2(V1,V2,V3)=gc(V1+V3-2V2),其他参数与式(1)、式(2)一致.下面利用该混合神经元耦合体系,考察体系的同步作用机理.由于FHN神经元的动力学行为与模型中的三个参数a,b,c有关,因此下面我们分别考察和讨论这些参数作用范围内的最佳取值. 在ML-FHN-ML模型中,由于FHN神经元模型内部参数a,b,c的取值均有一定的限定范围,因此,首先,本文中我们在合理范围内随机设定b值和c值,例如b=0.5,c=0.8.选取参数a为控制变量,考察系统同步状态随着该参数变化的行为和规律. 我们在a∈(2/3,1)范围内调节a的大小,分别选取几个有代表性的值a=0.70,0.80,0.90,0.95,体系的同步因子随耦合强度gc变化的行为如图2(a)所示.从图中可以看出,当a取某一值时,调节耦合强度,在gc较小区域,同步因子做不规则的振荡变化.其原因在于,引入新的神经元FHN后,其ML神经元的膜电压产生了类似图1(a)~(e)所示的多种振荡模式,因而可以同步到不同的振荡模式状态上.随着耦合强度的增加,同步因子将不断降低,表明耦合体系逐渐趋于同步.当耦合强度增加到一定值时,体系呈现出完全同步状态.改变a值大小,重复上述模拟过程,均能观察到类似现象.不过,对于不同的a值,使得体系处于同步状态时对应的耦合强度有所不同.图2(b)描述的是a在有意义的范围内取不同值时,能将体系调节到最佳同步状态所对应的耦合强度值gc.从图中可以看出,当a取0.95时,有一个最小的耦合强度gc=37.5,使得体系处于最佳同步状态.即0.95是参数a的最佳取值点. 其次,设定a=0.95,c=0.8,选取参数b为控制参量,在b的取值范围(0.075,0.64)内选取几个有代表性的点,考察参数b和gc对于组合模型系统同步的影响. 如图2(c)所示,在不同的参数b情况下系统达到同步时所需要的最小耦合强度gc不同,从图2(d)中可看出,随着b的增加,gc值开始略有上升,接着逐步下降并达到一个最小值,但当进一步增加b值时,gc值反而上升.这表明,参数b=0.35时系统达到同步所需要耦合强度gc值最小即gc=16.9.因此,b=0.35是参数b的最佳值点. 最后,考察参数c以及耦合强度对体系同步现象的调控作用.按照上述同样方法,设定a=0.95,b=0.35,选取参数c作为控制变量,在c∈(0.351/2,1)有意义范围内,考察不同c值对系统同步状态的影响.所得结果如图2(e)~(f)所示. 从图2(e)中可以看出,选取c为不同值时,系统达到同步所需要耦合强度gc的大小也不相同.gc随整个参数c区间变化示意图如图2(f)所示.随着c的增加,最佳耦合强度gc也不断减小,当c增加到0.9时,相应的耦合强度减小到最小值gc=14.4,随后,若进一步增加c的值,耦合强度gc反而上升.表明c的最佳值可取为0.90. 图2 不同参数a,b,c条件下同步误差随耦合强度变化示意图及同步状态时的耦合强度gc随参数a,b,c值变化关系图 (a)(c)(e):不同参数a,b,c条件下同步误差随耦合强度变化;(b)(d)(f):同步状态时的最佳耦合强度gc随参数a,b,c值变化 本文选取ML及FHN等神经元细胞为研究对象,构建多种神经元混杂的耦合模型,分别选取体系中的参数为控制参量,考察这些参数对耦合体系同步特性的调控作用.研究结果发现,其一,同种细胞构成的耦合体系(例如ML-ML-ML体系),调节耦合强度等参数就很容易使系统达到同步状态;若体系中包含有其他种类的神经元细胞时(例如ML-FHN-ML体系),则不仅要调节细胞之间的耦合参数,而且还要将体系内部的参数调节到合适的范围才行;其二,对于多种神经元耦合的非线性系统,由于有多种类型的神经元参与,使得细胞膜电压出现多种振动模式,因而,调节适当的参数,可以实现该种体系达到不同模式下的同步状态.上述结果表明,这种耦合体系在对外部刺激信号产生同步响应时,振荡模式越多,能够感受的外部刺激种类也就越多.实际上同步特性也与掺杂的第二种神经元的个数有关,即当引入的FHN神经元细胞个数适当时,可进一步提升同步响应效果(数据未给出).本文模拟实现了不同结构组合的神经元之间的最优同步,其结果不仅对将来深入理解耦合体系的同步机理有着一定的理论意义和指导作用,也为揭示大脑神经系统不同脑区之间的分工与协作机制提供一定的启示. [1] BORISYUK R,BORISYUK G,KAZANOVICH Y.Temporal structure of neural activity and modelling of information processing in the brain[J].Lecture Notes in Computer Science,2001,2036(3):237-254. [2] XU B,GONG Y B,WANG L,et al.Multiple synchronization transitions due to periodic coupling strength in delayed Newman-Watts networks of chaotic bursting neurons[J].Nonlinear Dynamics,2013,72(1-2):79-86. [3] CASADO J M,BALTANAS J P.Phase switching in a system of two noisy Hodgkin-Huxley neurons coupled by a diffusive interaction[J].Phys.Rev.E,2003,68(6):1-10. [4] FITZHUGH R.Impulses and physiological states in theoretical modelsof nerve membrane[J].Biophysical J,1961,1(6):445-466. [5] ZHANG J Q,HUANG S F,PANG S T.Optimizing calculations of coupling matrix in Hindmarsh-Rose neural network[J].Nonlinear Dyn,2016,84(3):1303-1310. [6] LIANG L S,ZHANG J Q,XU G X,et al.Effect of topology structures on synchronization transition in coupled neuron cells system[J].Commun.Theor.Phys,2013(60):380-386. [7] MORRIS C,LECAR H.Voltage oscillations in the barnacle giant muscle-fiber[J].Biophys J,1981,35(1):193-213. [8] WANG H X,LU Q S,WANG Q Y.Generation of firing rhythm,patterns and synchronization in the Morris-Lecar neuronModel[J].Int J Nonlin Sci Num,2005,6(1):7-12. [9] NAGUMO J,ARIMORO S,YOSHIZAWA S.An active pulse transmission line simulating nerve axion[J].Proc Ire,1962,50(10):2061-2070. [10] 刘勇,谢勇.分数阶FitzHugh-Nagumo模型神经元的动力学特性及其同步[J].物理学报,2010,59(3):2147-2155. [11] WANG Q Y,LU Q S.Transition to complete synchronization via near-synchronization in two coupled chaotic neurons[J].Chin.Phys,2005,14(11):2189-2195. [12] WANG Q,GONG Y B,LI H Y.Effects of channel noise on synchronization transitions in Newman-Watts neuronal networkwith time delays[J].Nonlinear Dynamics,2015,81(4):1689-1697. [13] XU C,WU Y,LU L.Dynamics in a coupled FHN model with two different delays[J].J of Computers,2014,9(8):1834-42. [14] LIN Y.Periodic oscillation analysis for a coupled FHN network model with delays[J].Abstract & Applied Analysis,2013,2013(54):1-13. [15] BARTSCH R,KANTELHARDT J W,PENZEL T,et al.Experimental evidence for phase synchronization transitions in the human cardiorespiratory system[J].Phys.Rev.Lett,2007,98(5):054102. [16] UPADHYAY R K,MONDAL A,TEKA W W.Mixed mode oscillations and synchronous activity in noise induced modified Morris-Lecar neural system.Intern[J].Jof Bifurcation & Chaos,2017,27(5):1730019. [17] XU B,BINCZAK S,JACQUIR S,et al.Parameters analysis of FitzHugh-Nagumo model for a reliable simulation[J].Conf Proc IEEE Eng Med Biol Soc,2014(2014):4334-4337. [18] WANG Q,GONG Y B.Multiple coherence resonance and synchronization transitions induced by autaptic self-feedbackactivity in Newman-Watts neuron networks[J].Applied Mathematical Modelling,2016,40:7147-7155.2 模拟结果与分析
2.1 同种神经元耦合情形
2.2 不同种类神经元耦合情形
3 结论
猜你喜欢
空气动力学学报(2022年4期)2022-08-23
数学物理学报(2022年2期)2022-04-26
黑龙江大学自然科学学报(2022年1期)2022-03-29
电子制作(2019年16期)2019-09-27
现代装饰(2018年5期)2018-05-26
中国惯性技术学报(2015年1期)2015-12-19
中国生化药物杂志(2015年4期)2015-07-07
浙江大学学报(工学版)(2015年2期)2015-05-30
航空学报(2015年4期)2015-05-07
弹箭与制导学报(2015年1期)2015-03-11
