
基于事件相关电位P300的动物认知研究进展
2018-05-28 04:27张小琴鲜雪梅方光战
四川动物 2018年3期
张小琴, 鲜雪梅, 方光战
(1. 阿坝师范学院资源与环境学院,四川汶川623002; 2. 中国科学院成都生物研究所两栖爬行动物研究室,成都610041)
事件相关电位(event-related potential,ERP)是感觉运动或认知事件在大脑内诱发的、具有锁时关系的电位变化,与认知过程(包括注意、辨别、判断、选择和决策等)密切相关,其幅度和潜伏期可用于考察大脑对信息的加工过程、加工效率和时间进程。ERP作为研究认知功能的电位生理学方法,在临床及动物认知研究中有广泛的应用前景(Duncanetal. ,2009)。ERP包含多种成分,其中由英国人Sutton 1965年发现的第三个正相波是大脑诱发电位的晚成分,因其与认知过程密切相关,故被称为认知诱发电位(cognitive evoked potential);同时该成分通常于刺激后300 ms左右出现,故被简称为P300(Suttonetal. ,1965)。一般用振幅(幅度或波幅)、潜伏期、头皮分布等参数来表征P300。振幅有2种常用测量方法:峰振幅测量,即特定时间窗内的最大峰值;平均振幅测量,即特定时间窗内所有振幅的平均值(Luck,2005)。对于特定ERP成分,潜伏期通常是指从刺激开始到最大振幅之间的持续时间(Johnson,1993),常用峰潜伏期(对应峰振幅)和半面积潜伏期(对应平均振幅)来描述。头皮分布是指ERP成分在不同脑区(对应于不同电极)的空间分布。
注意是心理活动对一定对象的指向和集中,伴随着感知觉、记忆、思维、想象等心理过程,具有指向性和集中性2个基本特征。Posner(1980)将注意分为内源性注意(endogenous)和外源性注意(exogenous):前者指根据观察者的行为目标或意图来分配注意,后者指由外部信息引起的注意定向。P300是ERP的主要内源性成分(Polich,2007),是在注意某一客体并进行认知加工时产生的,是高级思维活动如感觉、知觉、识别和判断等过程的结果,亦是大脑各皮质间综合活动的一种表现(Twomeyetal. ,2015)。因此,P300是研究人类(Tsolakietal. ,2015)、灵长类(Pineda & Westerfield,1993;Swicketal. ,1994a)、哺乳类(Ehlersetal. ,2014;Grupeetal. ,2014)和两栖爬行类(Elliott & Kelley,2007;Fangetal. ,2015)等认知加工及脑机制的重要电生理指标(魏景汉,罗跃嘉,2010),在神经病学、精神病学、心理学及其他临床学科、动物认知和大脑功能、脑机接口、测谎、毒理药理等研究领域具有广泛的应用价值。
P300通常由Oddball实验范式的偏差刺激诱发,即以不同概率呈现刺激,大概率者称为标准刺激,小概率者称为偏差刺激或靶刺激(图1)。P300可在视觉、听觉、嗅觉、体感等多种模态或视听双模态下诱发,其幅度可用来辨别刺激差异(Johnson,1989b;Miltneretal. ,1989;Yamaguchi & Knight,1991b)。与听觉、嗅觉2种模态相比,视觉P300的幅度和潜伏期均较大,但P300在视觉、听觉、嗅觉3种模态下的头皮分布无本质差别(Pictonetal. ,1984),表明3种模态下的P300均由共同的神经系统产生(Snyderetal. ,1980),但这一推论尚存争论。动物模型ERP的研究也显示存在类似的P300成分,其潜伏期和振幅具有任务特异性(Hattorietal. ,2010);在实际的动物研究中,往往参考人类P300的潜伏期。P300家族中最常见的亚成分是P3a,其潜伏期通常较短[对人而言,不同研究采用的范围略有不同,如视觉任务中取280~360 ms(Barceló & Rubia,1998)、听觉任务中取250~400 ms(Wronkaetal. ,2012)等],主要分布于额叶-中央区域,是定向反应出现的标志(Friedmanetal. ,2001);同时P3a与大脑抑制以及新颖刺激加工有关,反映新颖刺激处理过程的早期阶段。经典的P300实际上是P3b,其潜伏期较长[对人而言,不同研究采用的范围亦略有差异,如视觉任务中取450~600 ms(Barceló & Rubia,1998)、听觉任务中取400~700 ms(Wronkaetal. ,2012)等],依赖于被试主动辨别刺激类型,即由任务相关的靶刺激诱发,它要求被试在接受靶刺激后立即执行某种特定任务,且在顶叶波幅最大。P3a是对信号的初始反应,反映了从刺激出现到大脑开始意识到刺激信号并达到觉醒状态所需的时间,而P3b反映了大脑对信号的注意和记忆过程所需时间(Polich,2007)。Soltani和Knight(2000)认为,P3a与新颖刺激的自动觉察有关,而P3b与靶刺激的主动觉察有关。人类P300潜伏期与识别需要的时间及对刺激的反应时间都有关(Allen,2002)。
1 基于P300的动物认知研究
动物相关研究往往采用经典的Oddball范式,通常与操作性条件反射、经典条件反射和被动观看、训练动物参与任务等技术结合。现有的动物实验表明,包括哺乳动物在内的多个类群在多种模态中均存在类似人类P300成分,通常称为类P300(like-P300,lP300),其潜伏期和波幅随着任务不同而变化(Imada,2013)。理想的动物模型是脑疾病机理研究和药物开发的重要基础,亦被当作连续记录ERP特征和大脑皮层功能定位的工具(Pineda & Westerfield,1993),虽然这一技术存在2个重要局限:(1)在一个物种上观察到的ERP和另外一个物种是否相同,很难证明;(2)人类的一些ERP成分不能直接泛化到动物上,因为它们可能不适用,比如语言特殊反应。为了规避这些局限,研究者通过一系列任务操作来证明在动物上发现的ERP成分与人类相同或相似;此时要求动物和人类的行为及大脑结构必须存在对应的关联。
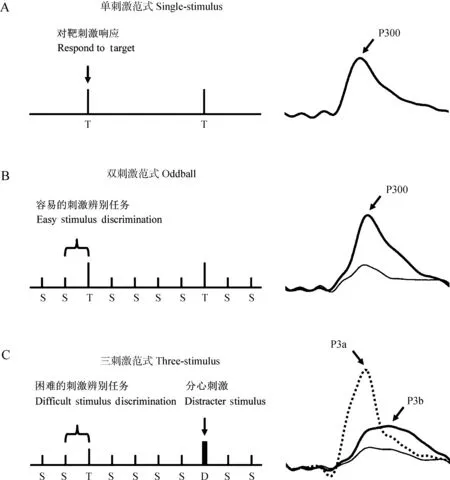
图1 诱发P300的实验范式
Fig. 1 Schematic illustration of paradigms eliciting P300
A、B、C分别对应于单刺激范式、双刺激范式和三刺激范式; 每个子图右侧呈现相应的P300成分; 在单刺激范式中,仅呈现概率小的靶刺激(T), 而无其他刺激; 在双刺激范式中, 按随机顺序呈现2种不同刺激, 其中一种刺激的出现概率少于另一种[前者为靶刺激(T), 后者为标准刺激(S)]; 三刺激范式类似于双刺激范式, 只是另外增加了一个呈现概率小的分心刺激(D); 在每一项任务中, 被试仅对靶刺激T响应; 三刺激范式中的分心刺激会诱发出P3a, 靶刺激会诱发出P3b(Polich & Criado, 2006)
The single-stimulus (A), oddball (B), and three-stimulus (C) paradigms are shown with the elicited ERPs from the stimuli of each task presented at the right; the single-stimulus task presents an infrequent target (T) in the absence of any other stimulus; the oddball task presents 2 different stimuli in a random sequence, with one occurring less frequently than the other (target=T, standard=S); the three-stimulus task is similar to the oddball with a compelling distracter (D) stimulus that occurs infrequently; in each task, the subject is instructed to respond only to the target and to refrain from responding otherwise; the distracter in the three-stimulus task elicits a P3a, and target elicits a P3b (Polich & Criado, 2006)
训练豚尾猴Macacanemestina从高概率呈现的纯音(非靶刺激2.5 kHz,持续时间400 ms)中辨别稀少的纯音(靶刺激1 kHz,持续时间400 ms),然后利用这2种刺激进行ERP实验,发现豚尾猴的事件相关正成分lP300类似于人类的P300,且振幅与刺激概率成反比(Arthur & Starr,1984)。同样猫Feliscatus(O’connor & Starr,1985)亦有相似的听觉lP300响应。对松鼠猴Saimirisciureus肌肉注射可乐定(α-2肾上腺素能受体激动剂,能减少去甲肾上腺素释放和抑制蓝斑神经元活动),发现其lP300幅度显著减少,潜伏期显著增加,但对ERP早期成分(P1、N1、P2)无影响(Swicketal. ,1994a),说明蓝斑及其传出投射系统在调制lP300活动中起重要作用。
在视觉研究方面,给松鼠猴呈现小的蓝色矩形作为标准刺激(90%或50%),小的黄色矩形作为偏差刺激(10%或50%),刺激间隔为2 s,发现概率为10%的偏差刺激诱发的lP300波幅比50%的更大,同时lP300幅度对刺激序列和任务敏感(Pineda & Swick,1992),这一结果类似于人类视觉P300。同样短尾猴Macacaarctoides(Snyderetal. ,1979)和食蟹猴Macacafascicularis(Ghilardietal. ,1987;Gloveretal. ,1991)的视觉lP300亦与人类类似。给松鼠猴呈现3 kHz纯音(70 db,持续时间50 ms)作为听觉标准刺激,1 kHz纯音(70 db,持续时间50 ms)作为偏差刺激,同时1个黄色矩形(持续时间50 ms)作为稀少视觉刺激,采集给予安慰剂或不同浓度α肾上腺素受体拮抗剂时的ERP波形,结果发现,听觉偏差刺激和视觉刺激均诱发出大的lP300,两者具有不同的分布且有不同的来源,支持2个独立源的存在;药物操作导致顶叶区域lP300幅度降低。这些数据支持去甲肾上腺素是一种调节位于顶叶区域的听觉和视觉lP300共有的神经递质这种假设;另外,中线和颞叶对应的lP300不受去甲肾上腺素活性的影响(Pineda & Westerfield,1993)。注射可乐定后,相对于听觉lP300,偏差刺激的差异大小更易影响松鼠猴视觉lP300的振幅大小,即当视觉偏差刺激与背景差异大时,更易获得显著的振幅差异。表明相对于听觉lP300,松鼠猴的视觉lP300对可乐定更为敏感。松鼠猴内侧颞叶的双侧损伤对硬脑膜外lP300没有影响(Palleretal. ,1988),双边内侧颞叶病变的松鼠猴仍能产生lP300,这表明内侧颞叶并非lP300产生的关键。同样,松鼠猴的lP300也受类胆碱和肾上腺素的影响(O’Neilletal. ,2000)。因此,灵长类存在与人类相似的P300这一事实为进一步探索该成分潜在的神经机制和解剖学基础提供了可能(Arthur & Starr,1984)。在被动非任务条件和主动任务条件下,分别记录食蟹猴的听觉lP300,发现其对偏差刺激的响应十分类似于相似条件下的人类P300,且可能与记忆功能和内侧颞叶活动有关,尤其是海马(Palleretal. ,1988;Swicketal. ,1994b)。依次切除双侧海马、杏仁核和皮层,食蟹猴出现类似于人类的遗忘症(Zola-Morganetal. ,1982),说明灵长类与人类的大脑功能具有同源性。被动条件和主动条件下的2种实验可能反映了lP300的2个成分P3a和P3b,但要严格区分这2个成分是不可能的。在听觉任务中(无论是被动条件还是主动条件),呈现概率小的音调能诱发出灵长类的晚正成分,该成分的波形特征、潜伏期、极性和头皮分布均类似于人类的P300,提示其功能特征的相似性。在连续双任务中,对第一个任务的lP300波幅响应随着第二个任务处理需求的增加而减少,亦与人类类似;虽然相对于任务条件,在非任务条件下,食蟹猴的lP300波幅较人类的更小,但其他特征与人类的极度相似性进一步为跨物种相似提供了支持,同时也反映了常见(或同源)的神经生理学过程。食蟹猴的lP300诱发不需要海马、杏仁核或邻近皮质的完整性,如双侧内侧颞叶病变的食蟹猴诱发的lP300与健康个体的lP300基本相似。这种人类与灵长类研究结果的融合有望提高动物模型的有效性,并进一步完善对P300神经基础和相关认知过程的理解(Palleretal. ,1988)。
研究发现,边缘系统、缘上回和海马都对猫听觉lP300的产生有贡献,膈区损毁后,猫的lP300消失(O’connor & Starr,1985);同时lP300的潜伏期在头骨记录、硬膜外记录和颅内记录三者间具有明显差异。损伤初级听觉皮层对猫颅内lP300的振幅没有明显影响(Harrisonetal. ,1986),提示猫的lP300存在多个起源,这一点与人类研究结果类似。而宽吻海豚Tursiopstruncatus对听觉偏差刺激产生一个长潜伏期的增强型正向成分(P550),类似于人类的P300(Woodsetal. ,1986)。对家兔Oryctolaguscuniculus的听觉ERP研究表明,在前扣带皮层和丘脑均能记录到lP300电位,而且lP300取决于皮质毒蕈碱的受体激活,在顶叶皮层注射盐酸利多卡因(酰胺类局麻药)(Wangetal. ,1998)或莨菪碱(乙酰胆碱拮抗剂)(Wangetal. ,1999)均能导致顶叶lP300振幅减少,说明顶叶皮层是局部lP300产生的必需结构(Wangetal. ,1998,1999)。另外,Meynert基底核可能亦是家兔lP300的起源之一(Wangetal. ,1997)。
通过被动实验范式和主动Go/No-Go范式,重点分析大鼠Rattusnorvegicus前额和枕叶等注意相关脑区的ERP成分,帮助判别各种峰值的功能意义以及比较大鼠和人类之间的异同(Imada,2013;Ehlersetal. ,2014)。研究显示,大鼠与人类具有相同的ERP成分和极性,且主动范式诱发的lP300波幅大于被动实验范式诱发的lP300(Hattorietal. ,2010)。据此推断,大鼠和人类可能有着相似的注意ERP成分的发生源(Sambethetal. ,2003)。与人类类似,大鼠在主动任务和被动任务下诱发出的ERP幅度不同(前者更大),说明大鼠和人类在感觉信息加工上可能存在相似性(Utheretal. ,2003)。
有意思的是,在诸如蛙类这样低等的动物中亦存在lP300。以等概率范式给峨眉仙琴蛙Babinadaunchina呈现白噪声、2种同种鸣叫(从洞内发出和洞外发生的鸣叫),发现白噪声诱发的lP300幅度显著大于同种鸣叫,而2种同种鸣叫间的lP300幅度无显著差异,说明仙琴蛙能对声音进行分类,即将白噪声从同种鸣叫中区分开来(Fangetal. ,2015)。具体来说,每种刺激的呈现概率占33.3%,同种鸣叫的呈现概率就是66.7%,为大概率的标准刺激;白噪声为偏差刺激,能诱发出更大的lP300。该研究表明,lP300非常适合于研究动物物种识别和个体识别等认知行为,对描述动物认知过程及其神经机制起着不可或缺的作用。
2 P300的起源
正常人、患者及多个动物类群均可诱发出P300(Polich,1987;Nieuwenhuisetal. ,2005)。对脑内源进行研究有助于揭示P300的生理、心理实质和进一步认识大脑功能(Johnson,1993)。目前对于P300的起源有着多种假说:
一是起源于海马。海马位于人类和其他灵长类动物大脑颞叶内部,具有调节情绪、参与学习记忆等功能。P3a依赖于参与注意力维持的额叶和参与处理新颖信息的海马(Knight,1997;Polich,2003);而P3b可能起源于海马(Slaweckietal. ,2000;Polich & Criado,2006)。但在双侧颞叶切除和海马毁损的实验中(Johnson,1989a)并未观察到lP300波幅明显减少,说明海马并非lP300的直接起源(Nieuwenhuisetal. ,2005)。
二是起源于前扣带回(anterior cingulated gyrus,ACG)(Hattorietal. ,2010)。ACG属于边缘系统,包括胼胝体、扣带回以内的环状部分,是一非均质结构,功能不尽相同。因此有研究认为,P3a往往在刺激间有较大差异时出现(不论被试是否作出反应);无需执行任务,是被试对随机出现的、与背景刺激不同的偏差刺激在大脑中的被动响应,它的起源更倾向于前额-中央皮质(Ehlersetal. ,1994)。
三是多部位起源(Isrealetal. ,1980)。Halgren等(1998)记录P300时同步记录颅内脑电图(intracranial electroencephalogram,iEEG),发现多个脑区(包括额颞叶前部、颞叶上部、后顶叶的广泛皮层及海马区)的iEEG活跃,认为P300在脑内的起源十分广泛,涉及各个系统。切除癫痫患者单侧颞叶后,其P300波幅并没有明显降低(Johnson,1989);双内侧颞叶损毁的食蟹猴依然能产生lP300(Palleretal. ,1988),这些结果表明颞叶并非是产生lP300的关键脑区。对松鼠猴听觉和视觉ERP的研究发现,lP300的起源在左右半球是相对独立的(Pineda & Westerfield,1993);切除食蟹猴的双侧海马、杏仁核及上覆皮质结构(Palleretal. ,1988)或猫的初级听觉皮层和联合皮层(Harrisonetal. ,1990),lP300并未受到显著影响,说明这些脑区并非lP300起源的必要脑区。这些结果表明,单一脑结构不能解释不同认知任务下的lP300产生(Utheretal. ,2003),药理学研究亦发现lP300可能具有多个神经起源(Pineda & Westerfield,1993)。另外,P3a和P3b不仅是P300的2个功能独特的电位成分,而且有各自独立的多重皮层起源(Soltani & Knight,2000)。前者主要定位在额叶皮层和前扣带回,后者起源包括顶上小叶和部分后扣带回(Wronkaetal. ,2012)。因此,Polich(2007)提出一种认知模型:P3a起源于任务处理过程中刺激驱动的前额注意机制;而P3b起源于注意相关的颞顶活动,且与记忆处理相关。
3 影响P300的因素
波幅一般反映神经活动范围的信息,波面积代表大脑神经元在单位时间内的发放总和,并且面积与记忆清晰度相关(Hermensetal. ,2010)。影响P300波幅的因素包括刺激呈现概率(Calcusetal. ,2014)、刺激顺序、刺激质量和刺激间隔(Duncanetal. ,2009;Wangetal. ,2015)、刺激强度(Lindínetal. ,2005)、注意力和刺激任务的相关性(Patel & Azzam,2005)、刺激意义、任务难度、动机和警觉(Sommer & Matt,2010)等。潜伏期表明激活的开始,P300潜伏期反映了记忆场景更新时,大脑对刺激的辨认速度和选择过程(Dienetal. ,2004)。潜伏期与被试对刺激做出辨别时中枢所需的皮层突触数量相关,参与的突触数量越多,潜伏期越长(Joosetal. ,2014);同时也可能和心理功能的速度相关,较短的潜伏期对应较快的认知功能:随着正常儿童发育期、正常老龄化和痴呆的顺序,潜伏期增加(van Dinterenetal. ,2014)。
P300波幅和潜伏期均与任务难度正相关(Polich,1987),这是因为任务难度越大,包含信息越多,从而需要投入更多的认知加工资源,这种效应也称为任务效应。刺激模态不同,产生的P300亦不同,如,视觉刺激对应的潜伏期较躯体刺激短,听觉刺激对应的P300则较稳定、变异小、波形易辨认、无视力影响因素,目前应用广泛;刺激强度越大,间期越长(Junetal. ,1999),或偏差刺激概率越小(Snyderetal. ,1980),P300波幅越大,而潜伏期一般无变化。视觉、听觉和触觉P300波幅和潜伏期均与年龄呈负相关(Yamaguchi & Knight,1991a;Tsolakietal. ,2015):从10岁左右开始,随着年龄的增长,潜伏期逐渐缩短,波幅逐渐增高,20岁左右时潜伏期最短、波幅最高;之后随年龄增长,潜伏期又以每年1~2 ms的幅度增宽,波幅逐渐降低(Johnson,1989b),但反应时间没有明显改变(Pictonetal. ,1984;Yamaguchi & Knight,1991a)。P300波幅还受遗传因素影响,是高度遗传的表型,其遗传度为39%~79%(Euseretal. ,2012)。关于P300的性别差异尚存争论,部分关于人和蛙类的研究均显示其无性别差异(Husteretal. ,2011;Fangetal. ,2015),虽然在执行Oddball任务中发现男性的反应时间更短(Glaseretal. ,2012;Tsolakietal. ,2015);但另一些研究发现,性别会影响P300的波幅和潜伏期(Arnoneetal. ,2011;Glaseretal. ,2012)。另外,睡眠、认知语境、唤醒度、注意水平(Halgrenetal. ,1980;Swicketal. ,1994b;Polich & Kok,1995)、利手、情绪效价及唤醒度的性别差异均会影响P300的波幅和潜伏期(Arnoneetal. ,2011;Glaseretal. ,2012)。
4 展望
目前关于P300神经基础研究已有较大进展,但其大脑起源及各起源之间的作用机制仍然不甚明晰。首先,P300的起源有哪些?这些起源或多个处理系统如何连接产生定向反应或侦测行为的?不同模态下的起源是否具有共性和特异性差异?可考虑用脑网络方法对此进行深入研究。其次,P300的特征、起源及与大脑认知功能的联系在不同物种中存在何种差异?利用ERP方法在不同物种中进行比较生理学研究,从而揭示其中的差异。这类研究将有助于确定非人动物的lP300研究成果在多大程度上可以向其他物种特别是人类泛化。最后,P300在不同物种间的差异与大脑功能进化和生物多样性形成的关系如何?P300的特征在不同种群中是否存在差异?同域分布与否和环形种与否是否影响P300的特征?可从进化的角度对这一问题进行深入研究,进一步了解P300与动物认知能力之间、P300与种群扩散之间及P300与生物多样性形成之间的关联。
:
魏景汉, 罗跃嘉. 2010. 事件相关电位原理与技术[M]. 北京: 科学出版社: 52-55.
Allen JJB. 2002. The role of psychophysicology in clinical assessment: ERPs in the evaluation of memory[J]. Psychophysiology, 39(3): 261-280.
Arnone B, Pompili A, Tavares MC,etal. 2011. Sex-related memory recall and talkativeness for emotional stimuli[J]. Frontiers in Behavioral Neuroscience, 5: 52.
Arthur DL, Starr A. 1984. Task-relevant late positive component of the auditory event-related potential in monkeys resembles P300 in humans[J]. Science, 223(4632): 186-188.
Barceló F, Rubia FJ. 1998. Non-frontal P3b-like activity evoked by the Wisconsin Card Sorting Test[J]. Neuroreport, 9(4): 747-751.
Calcus A, Deltenre P, Hoonhorst I,etal. 2014. MMN and P300 are both modulated by the featured/featureless nature of deviant stimuli[J]. Clinical Neurophysiology, 126(9): 1727-1734.
Dien J, Spencer KM, Donchin E. 2004. Parsing the late positive complex: mental chronometry and the ERP components that inhabit the neighborhood of the P300[J]. Psychophysiology, 41(5): 665-678.
Duncan CC, Barry RJ, Connolly JF,etal. 2009. Event-related potentials in clinical research: guidelines for eliciting, recording, and quantifying mismatch negativity, P300, and N400[J]. Clinical Neurophysiology, 120(11): 1883-1908.
Ehlers CL, Desikan A, Wills DN. 2014. Event-related potential responses to the acute and chronic effects of alcohol in adolescent and adult Wistar rats[J]. Alcoholism Clinical and Experimental Research, 38(3): 749-759.
Ehlers CL, Kaneko WM, Robledo P,etal. 1994. Long-latency event-related potentials in rats: effects of task and stimulus paramenters[J]. Neuroscience, 62(3): 759-769.
Elliott TM, Kelley DB. 2007. Male discrimination of receptive and unreceptive female calls by temporal features[J]. Journal of Experimental Biology, 210(Pt 16): 2836-2842.
Euser AS, Arends LR, Evans BE,etal. 2012. The P300event-related brain potential as a neurobiological endophenotype for substance use disorders: a meta-analytic investigation[J]. Neuroscience & Biobehavioral Reviews, 36(1): 572-603.
Fang GZ, Yang P, Xue F,etal. 2015. Sound classification and call discrimination are decoded in order as revealed by event-related potential components in frogs[J]. Brain Behavior Evolution, 86(3-4): 232-245.
Friedman D, Cycowicz YM, Gaeta H. 2001. The novelty P3: an event-related brain potential (ERP) sign of the brain’s evaluation of novelty[J]. Neuroscience & Biobehavioral Reviews, 25(4): 355-373.
Ghilardi MF, Marx MS, Onofrj MC,etal. 1987. Scalp distribution of pattern visual evoked potentials in normal and hemianopic monkeys[J]. Physiology & Behavior, 41: 297-302.
Glaser E, Mendrek A, Germain M,etal. 2012. Sex differences in memory of emotional images: a behavioral and electrophysiological investigation[J]. International Journal of Psychophysiology, 85(1): 17-26.
Glover A, Ghilardi MF, Bodis-Wollner I,etal. 1991. Visual ‘cognitive’ evoked potentials in the behaving monkey[J]. Electroencephalography Clinical Neurophysiology, 90(1):65-72.
Grupe M, Grunnet M, Laursen B,etal. 2014. Neuropharmacological modulation of the P3-like event-related potential in a rat two-tone auditory discrimination task with modafinil and NS9283, a positive allosteric modulator of alpha4 beta2 nAChRs[J]. Neuropharmacology, 79: 444-455.
Halgren E, Marinkovic K, Chauvel P. 1998. Generators of the late cognitive potentials in auditory and visual oddball tasks[J]. Electroencephalography and Clinical Neurophysiology, 106(2): 156-164.
Halgren E, Squires N, Wilson C,etal. 1980. Endogenous potentials generated in the human hippocampal formation and amygdala by infrequent events[J]. Science, 210(4471): 803-805.
Harrison J, Buchwald J, Kaga K. 1986. Cat P300 present after primary auditory cortex ablation[J]. Electroencephalography & Clinical Neurophysiology, 63(2): 180-187.
Harrison JB, Dickerson LW, Song S,etal. 1990. Cat-P300 present after association cortex ablation[J]. Brain Research Bulletin, 24(4): 551-560.
Hattori M, Onoda K, Sakata S. 2010. Identification of rat P3-like processes in the anterior cingulate cortex and hippocampus[J]. Neuroscience Letters, 472(1): 43-46.
Hermens DF, Ward PB, Hodge M,etal. 2010. Impaired MMN/P3a complex in first-episode psychosis: cognitive and psychosocial associations[J]. Progress in Neuro-Psychopharmacology & Biological Psychiatry, 34(6): 822-829.
Huster RJ, Westerhausen R, Herrmann CS. 2011. Sex differences in cognitive control are associated with midcingulate and callosal morphology[J]. Brain Structure & Function, 215(3-4): 225-235.
Imada A. 2013. Analysis of rat event related potentials in frontal and parietal lobes as a possible neural correlate of attention in passive Oddball and active Go/No-Go paradigms[D]. Massachusetts: Wellesley College: 11-20.
Isreal JB, Chesney GL, Wickens CD,etal. 1980. P300 and tracking difficulty: evidence for multiple resources in dual-task performance[J]. Psychophysiology, 17(3): 259-273.
Johnson RJ. 1989a. Auditory and visual P300s in temproal lobectomy patients-evidence for modailtiy-dependent generators[J]. Psychophysiology, 26(6): 633-650.
Johnson RJ. 1989b. Developmental evidence for modality-dependent P300 generators: a normative study[J]. Psychophysiology, 26(6): 651-667.
Johnson RJ. 1993. On the neural generators of the P300 component of the event-related potential[J]. Psychophysiology, 30(1): 90-97.
Joos K, Gilles A, Van de Heyning,etal. 2014. From sensation to percept: the neural signature of auditory event-related potentials[J]. Neuroscience & Biobehavioral Reviews, 42(5): 148-156.
Jun J, Porjesz B, Begleiter H,etal. 1999. P300: the similarities and differences in the Scalp distribution of visual and auditory modality[J]. Brain Topography, 11: 315-327.
Knight RT. 1997. Distributed cortical network for visual attention[J]. Journal of Cognitive Neuroscience, 9(1): 75-91.
Lindín M, Zurrón M, Díaz F. 2005. Stimulus intensity effects on P300 amplitude across repetitions of a standard auditory oddball task[J]. Biological Psychology, 69(3): 375-385.
Luck SJ. 2005. An introduction to the event-related potential technique[M]. Cambridge, Massachusetts: MIT Press:229-235.
Miltner W, Johnson JR, Braun C,etal. 1989. Somatosensory event-related potentials to painful and non-painful stimuli: effects of attention[J]. Pain, 38(3): 303-312.
Nieuwenhuis S, Aston-Jones G, Cohen JD. 2005. Decision making, the P3, and the locus coeruleus-norepinephrine system[J]. Psychological Bulletin, 131(4): 510-532.
O’connor TA, Starr A. 1985. Intracranial potentials correlated with an event-related potential, P300, in the cat[J]. Brain Research, 339(1): 27-38.
O’Neill J, Halgren E, Marinkovic K,etal. 2000. Effects of muscarinic and adrenergic agonism on auditory P300 in the macaque[J]. Physiology & Behavior, 70(1-2): 163-170.
Paller KA, Zolamorgan S, Squire LR,etal. 1988. P3-like brain waves in normal monkeys and in monkeys with medial temporal lesions[J]. Behavioral Neuroscience, 102(5): 714-725.
Patel SH, Azzam PN. 2005. Characterization of N200 and P300: selected studies of the event-related potential[J]. International Journal of Medical Sciences, 2(4): 147-154.
Picton TW, Stuss DT, Champagne SC,etal. 1984. The effects of age on human event-related potentials[J]. Psychophysiology, 21(3): 312-326.
Pineda JA, Swick D. 1992. Visual P3-like potentials in squirrel monkey: effects of a noradrenergic agonist[J]. Brain Research Bulletin, 28(3): 485-491.
Pineda JA, Westerfield M. 1993. Monkey P3 in an “oddball” paradigm: pharmacological support for multiple neural sources[J]. Brain Research Bulletin, 31(6): 689-696.
Polich J, Criado JR. 2006. Neuropsychology and neuropharmacology of P3a and P3b[J]. International Journal of Psychophysiology, 60(2): 172-185.
Polich J, Kok A. 1995. Cognitive and biological determinants of P300: an integrative review[J]. Biological Psychology, 41(2): 103-146.
Polich J. 1987. Task difficulty, probability, and inter-stimulus interval as determinants of P300 from auditory stimuli[J]. Electroencephalography & Clinical Neurophysiology, 68(4): 311-320.
Polich J. 2003. Theoretical overview of P3a and P3b[M]// Polich J. Detection of change: event-related potential and fMRI findings. Boston: Kluwer Academic Press: 83-98.
Polich J. 2007. Updating P300: an integrative theory of P3a and P3b[J]. Clinical Neurophysiology, 118(10): 2128-2148.
Posner MI. 1980. Orienting of attention[J]. Quarterly Journal of Experimental Psychology, 32(1): 3-25.
Sambeth A, Maes JH, van Luijtelaar G,etal. 2003. Auditory event-related potentials in humans and rats: effects of task manipulation[J]. Psychophysiology, 40(1): 60-68.
Slawecki CJ, Thomas JD, Riley EP,etal. 2000. Neonatal nicotine exposure alters hippocampal EEG and event-related potentials (ERPs) in rats[J]. Pharmacology Biochemistry & Behavior, 65(4): 711-718.
Snyder E, Hillyard SA, Galambos R. 1980. Similarities and differences among the P3 waves to detected signals in three modalities[J]. Psychophysiology, 17(2): 112-122.
Snyder EW, Beck EC, Dustman RE. 1979. Visual evoked potentials in monkeys[J]. Electroencephalography & Clinical Neurophysiology, 47(4): 430-440.
Soltani M, Knight RT. 2000. Neural origins of the P300[J]. Critical Reviews in Neurobiology, 14(3-4): 199-224.
Sommer W, Matt J. 2010. Awareness of P300-related cognitive processes: a signal detection approach[J]. Psychophysiology, 27(5): 575-585.
Sutton S, Braren M, Zubin J,etal. 1965. Evoked potential correlates of stimulus uncertainty[J]. Science,150:1187-1188.
Swick D, Pineda JA, Foote SL. 1994a. Effects of systemic clonidine on auditory event-related potentials in squirrel monkeys[J]. Brain Research Bulletin, 33(1): 79-86.
Swick D, Pineda JA, Schacher S,etal. 1994b. Locus coeruleus neuronal activity in awake monkeys: relationship to auditory P300-like potentials and spontaneous EEG[J]. Experimental Brain Research, 101(1): 86-92.
Tsolaki A, Kosmidou V, Hadjileontiadis L,etal. 2015. Brain source localization of MMN, P300 and N400: aging and gender differences[J]. Brain Research, 1603: 32-49.
Twomey DM, Murphy PR, Kelly SP,etal. 2015. The classic P300 encodes a build-to-threshold decision variable[J]. European Journal of Neuroscience, 42(1): 1636-1643.
Uther M, Jansen DHJ, Huotilainen M,etal. 2003. Mismatch negativity indexes auditory temporal resolution: evidence from event-related potential (ERP) and event-related field (ERF) recordings[J]. Brain Research Cognitive Brain Research, 17(3): 685-691.
van Dinteren R, Arns M, Jongsma ML,etal. 2014. Combined frontal and parietal P300 amplitudes indicate compensated cognitive processing across the lifespan[J]. Frontiers in Aging Neuroscience, 6(6): 294.
Wang F, He Y, Pan J,etal. 2015. A novel audiovisual brain-computer interface and its application in awareness detection[J]. Scientific Reports, 5: 9962.
Wang Y, Kawai Y, Nakashima K. 1999. Rabbit P300-like potential depends on cortical muscarinic receptor activation[J]. Neuroscience, 89(2): 423-427.
Wang Y, Nakashima K, Shiraishi Y,etal. 1997. P300-like potential disappears in rabbits with lesions in the nucleus basalis of Meynert[J]. Experimental Brain Research, 114(2): 288-292.
Wang Y, Shiraishi Y, Kawai Y,etal. 1998. Cortical anesthesia reduced the amplitude of local P300 event-related potential in rabbits by auditory oddball paradigm[J]. Neuroscience Letters, 244: 25-28.
Woods DL, Ridgway SH, Bullock TH. 1986. Middle- and long-latency auditory event-related potentials in dolphins[M]// Schusterman JR, Thomas JA, Wood FG. Dolphin cognition and behavior: a comparative perspective. London: Lawrence Erlbaum Associates: 61-77.
Wronka E, Kaiser J, Coenen AM. 2012. Neural generators of the auditory evoked potential components P3a and P3b[J]. Acta Neurobiologiae Experimentalis, 72(1): 51-64.
Yamaguchi S, Knight RT. 1991a. Age effects on the P300 to novel somatosensory stimuli[J]. Electroencephalography & Clinical Neurophysiology, 78(4): 297-301.
Yamaguchi S, Knight RT. 1991b. P300 generation by novel somatosensory stimuli[J]. Electroencephalography & Clinical Neurophysiology, 78(1): 50-55.
Zola-Morgan S, Squire LR, Mishkin M. 1982. The neuroanatomy of amnesia: amygdala-hippocampus versus temporal stem[J]. Science, 218(4579): 1337-1339.
猜你喜欢
哈尔滨工程大学学报(2022年8期)2022-09-11
小学科学(学生版)(2021年12期)2021-12-31
保健与生活(2021年21期)2021-11-11
小学科学(学生版)(2020年7期)2020-07-28
家庭科学·新健康(2019年9期)2019-10-21
价值工程(2018年25期)2018-09-26
中国漫画(2017年10期)2018-01-25
意林·少年版(2018年24期)2018-01-05
中国现代医生(2017年24期)2017-10-17
中国漫画(2017年4期)2017-06-30
