
草地生态系统生物和功能多样性及其优化管理
2018-07-04 01:59宋明华刘丽萍陈锦张宪洲
生态环境学报 2018年6期
宋明华,刘丽萍 ,陈锦 ,张宪洲
草地生态系统是自然生态系统的重要组成部分,对维系生态平衡、地区经济、人文历史具有重要的价值。中国拥有天然草地面积近4亿公顷,是世界第二草地大国,分布区域广泛,其中以西藏自治区草地面积最大(Chen et al.,2000)。草地蕴涵了丰富而多样的物种资源,是地球上生物多样性的主要承载体之一,同时多样的物种也为草地生态系统提供了多样的生态和服务功能。伴随着探究物种多样性——生态系统生产力二者之间的关系,也即揭示生态系统结构——功能、格局——过程关系的发展历程,生态学的研究已经从简单二元变量关系拟合转向关注多元变量之间复杂多维网络联系,由从单一的结构推断和功能转向基于结构、组分之间机理联系预测和优化生态系统多元化功能网络。因此,揭示草地物种共存、多样性维持机制,探寻引起物种共存与多样性变化的生物和非生物因素,建立生物多样性与生态系统多功能之间的联系势在必行,也是当前草地生态学关注的前沿和面临的挑战(图1)。随着认识的不断深入,必将为草地生态系统的管理和可持续发展提供可行的科学依据,解释不同情境下草地生态系统多功能之间的权衡关系,实现科学管理方式调控下的草地生态系统多功能的预测以及生态与经济效益兼顾的优化发展模式(表1)。
1 生物多样性
生物多样性是生物与环境长期作用的结果,因此生物多样性的研究伴随着生物适应方式、策略的研究。在多样和多变的环境条件下,生物在不同的构件水平上对环境、资源等表现出了多样化的适应和应对方式,各构件之间存在着资源消费、分配的权衡,使得不同组分机能正常运行,提高整个机体的适合度(fitness)。
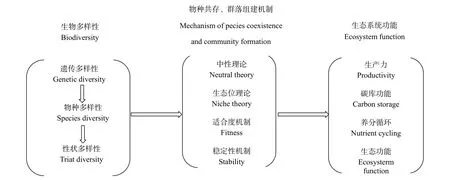
图1 生物多样性与生态系统多功能关系概念图Fig. 1 Conceptual map of the multifunctional relationship between biodiversity and ecosystems

表1 草地生态系统理论研究与利用管理的年代发展事件Fig. 1 The events of theory and the development of utilization and management of grassland ecosystem in the era
在讨论生物多样性之前,必须先明确什么是生物多样性。许多人认为生物多样性非常复杂,难以将其表述清楚,所以往往回避“生物多样性”的说法。生物多样性分为不同的层次,与生态学研究密切相关的生物多样性大体包括遗传多样性、物种多样性、性状多样性(Gillison et al.,2013)。这三方面是生态学问题关注的主要领域,其研究意义不仅体现于草地生态系统生物多样性的保护,也适用于其他类型的生态系统,将生物多样性和生态系统服务纳入多功能景观管理,以帮助我们更好地理解生物多样性在提供生态系统功能和人类-自然耦合系统服务方面的作用(Gamfeldt et al.,2013)。
1.1 生态学关注的生物多样性
1.1.1 遗传多样性
遗传多样性是生命进化和适应的基础。遗传多样性可能通过改变物种间相互作用的性质间接影响生态系统中物种的组成和功能(Fridley et al.,2010)。一般而言,种内遗传多样性越丰富,物种对环境变化的适应能力也越强,所以遗传多样性推动了物种进化(Byrne et al.,2000)。但是,近年来,各种因素导致了遗传多样性的丧失,如有效居群大小的变化(等位基因);近交繁殖使纯合性增加和后代适应能力下降;遗传漂变使杂合性降低和物种的微进化、基因流的减少或远交衰退使有效居群大小降低,形成恶性循环,居群变得格外脆弱,容易在环境发生变化时灭绝。此外,人类活动对遗传多样性的丧失也具有很大影响,如生境破碎化导致居群遗传结构的变化,加速了物种灭绝的进程(Lande,1988)。
种内遗传多样性对生态系统进程的影响受到越来越多的关注。种内遗传变异对生态系统进程的影响显著而广泛(Bailey et al.,2009)。因此,虽然世界范围内物种多样性下降的负面影响已被广泛承认(Hooper et al.,2005),群落遗传学认为,种内遗传多样性的丧失对多种生态系统的演化进程和群落带来了从未为人所知的后果(Whitham et al.,2006)。由于环境条件的影响,遗传多样性的影响在空间和时间上可能有所不同。这种情况下,在低多样性和高多样性的地块中,植物之间的适应度可能会产生差异,从而导致自然选择的异质性和进化轨迹的差异。Hughes et al.(2008)探讨了关于人们对遗传多样性带来的生态影响认识的主要差异。例如,有必要将遗传多样性的重要性与其他生态因素的重要性进行比较(Hughes et al.,2009)。虽然先前的研究通过展现遗传多样性的潜在重要性开辟了新的研究前沿,但这些研究往往在其实验中人为地减少了环境变化。因此,下一步研究是比较种内遗传多样性与其他生态因素的重要性,从而确立群落遗传学观点在理解群落结构和生态系统过程中的相关性(Tack et al.,2011)。
从进化过程中的物种选择和适者生存的角度,我们可以判断这样一个事实:遗传的均一性将最终威胁群体或物种的生存,导致生态系统中物种的濒危或灭绝。从遗传多样性来看,种内遗传多样性的保持有助于保持物种和整个生态系统的多样性,可以减慢由于适应和进化所导致的灭绝过程。遗传多样性和物种多样性是生态系统提供多功能服务的基础。遗传多样性越高,生态系统的稳定性相对更高。遗传多样性的维持可以增强生态系统的抗干扰能力,有利于生物多样性的保护,尤其是有助于人类对濒危物种的保护,将在育种、繁殖、基因保存与利用中达到保护珍稀物种和促进生物多样性的目的。同时,遗传多样性在调节种内和种间相互作用中有利于协助改善群落组成结构,影响群落演替、生态系统的发展进程和生态系统功能的发挥。而且,从目前对遗传多样性的研究来看,从单纯的遗传因素的研究到把遗传因素与其他因素的研究进行结合和比较,将丰富遗传多样性的研究课题和内容。
1.1.2 物种多样性
物种丰富度是指一定面积内种的总数目,它反映了物种的生态适应、扩散能力及进化历史(Kreft et al.,2006)。物种丰富度在大尺度分布格局上具有一定规律性,物种的丰富程度跟纬度呈明显的反比关系,离赤道越远,物种就越稀少(Qian et al.,2005;Huston,1994;Zhang et al.,1985);也受海拔影响,如物种的垂直分布(Bachman et al.,2004;Vetaas et al.,2002)。
过去,我们在研究草地生物多样性时,更多地考虑了植物物种数目和植食动物通过采食活动对植物种类和生物量所产生的影响,然后去评估草地生态系统功能的发挥。然而,如果不考虑到生态系统高营养水平层次的多样性,对生物多样性——生态系统功能关系的评估将是不完整的(Cardinale et al.,2006)。植物物种组成是生态系统功能的重要调节因子(Loreau et al.,2001;Chapin et al.,1997)。在草地生态系统中,由于禾草类的竞争优势是影响植物物种多样性和生态系统功能水平的重要因素(Schmitz,2008),捕食者的级联效应可能导致植物组成和生态系统功能的巨大变化。植物优势效应一般是生态系统结构和功能的重要决定因素(Hillebrand et al.,2008;Wilsey et al.,2005;Smith et al.,2004),捕食者功能多样性经常与生态系统功能的变化联系在一起(Schmitz,2009)。
要全面了解食物网变化对生态系统功能的影响,需将生物多样性——生态系统功能理论与关于营养结构影响的知识更深入地结合起来(Srivastava et al.,2009)。草地生态系统的物质流动和能量循环功能的研究离不开生产者、消费者和分解者的共同作用,因为营养级之间的能量和营养物质流动既受食物网营养结构的影响,也受营养级内物种多样性的影响。营养结构理论表明,营养级之间的能量流动关键取决于消费者的多样性。研究表明,物种丰富的群落和生态系统会表现出更强的抵御干扰能力或对干扰进行恢复的能力(Ollinger,2005),由此可见,物种间的多样性对维持生态系统的稳定性功能和长期的可持续性至关重要。在草地生态系统中,物种丰富度还有利于草地生产力的提高。一个生物群落中的生物数量是否对生态系统过程的性质有可预测性的影响是生态学中一个长期主题。
1.1.3 性状多样性
从植物性状多样性来看,植物适应环境变化会通过不同性状之间可塑性的权衡关系来实现,在适应环境过程中,多样性的提高增强了生态系统功能的稳定性(Roscher et al.,2013)。研究表明,多样性高的生态系统会提高系统的多功能,生态系统通常也具有丰富的功能性状多样性。
性状多样性对生态系统功能的影响主要体现在人类对生态系统发展进程和功能发挥的预测方面。物种的生态性状决定了它在生态系统功能中的作用。植物的功能性状是指植物体具有的与其定植、存活、生长和消亡紧密相关的一系列核心植物属性。根据与生态系统的关系,植物功能性状可以分为响应性状和影响性状。其中,影响性状决定了植物对生态系统功能的影响,如对生物地球化学循环的影响(孟婷婷等,2007),物种的功能性状间普遍存在着权衡关系,有利于植物选择最适的生存策略。植物性状多样性是生物对环境适应的结果,与环境和生态系统服务功能息息相关(雷羚洁等,2016)。因为生态环境变化引起了植物性状的变化,植物性状及功能的改变实际上反映了植被对环境变化的响应,也反映了该地区土壤、温度、水分、光照等外界因素的变化,所以性状多样性能够提供有效的生态指示作用,有助于及时发现生态问题,制定有效的生态对策防止环境进一步恶化,在维持生态系统的稳定性方面发挥着重要作用。通过对叶片性状(如叶面积、叶片营养元素等)和繁殖性状(种子、根系等)的研究有助于理解生态系统净初级生产力和营养循环的发展和变化。
因为物种之间的功能性状不同,物种组成的变化可以改变生态系统的功能,探讨不同性状以及性状在多个营养水平上的分布,对于深入了解生物多样性的丧失过程中,某些生态系统功能发生剧烈变化而另一些生态系统的功能仍保持复原力的差异至关重要,并可能有助于预测生态系统对环境变化的反应(Heilpern et al.,2018)。基于性状的方法可为我们提供更多有价值的信息,因为功能性状通过对物种生长、繁殖和生存的影响为生态学家提供了一个客观衡量生物体在生态系统功能中作用的指标(Bregman et al.,2015;Violle et al.,2007;Diaz et al.,2001)。
近年来,有许多生态学家利用系统发育多样性预测生物多样性对生态系统功能的影响。然而,系统发育多样性和功能多样性预测能力的比较实验尚不完整。为了探讨系统发育多样性和功能性状变异如何控制生物多样性对生物量生产的影响,Flynn et al.(2011)总结了草地29种植物实验的结果,实验中观测了植物的系统发育情况,为我们提供了大量的植物性状数据。因为系统发育多样性和功能多样性都可以作为生物多样性对生态系统功能影响的有价值的预测因子,因此,关注群落特征多样性和进化历程可以提高人们对生物多样性丧失导致的生态后果的认识。最近的研究表明,系统发育多样性可以作为衡量群落功能多样性的指标,二者之间的这种关系基于进化多样化产生的性状或特征多样化的合理假设,可能导致更强大的生态位互补,这一理论得到了生物多样性——生态系统功能研究的元分析的支持,虽然已有大量的实验证据表明生物多样性对生态系统功能产生了积极影响,特别是对草地地上植物生物量生产的影响(Duffy,2009;Balvanera et al.,2006),但是生物多样性的哪些方面对生态系统的功能影响最大,仍然是一个受争论的话题。
在预测生物多样性的影响时,应该直接比较系统发育多样性和群落功能多样性以及功能性状差异是如何影响生态系统发生变化的。利用有关物种性状功能特征和进化历史的信息资源,可以为预测物种灭绝在生态系统层面上产生的后果提供更有力的手段。然而,利用系统发育多样性预测生态系统功能仍然面临草原生物多样性——生态系统功能研究的具体问题。
1.2 生物多样性的研究历史和发展概要
自然生态系统中往往是多物种共存的,单一物种的群落很少见。那么物种为什么会牺牲自己的种群个体多度换取多物种共存呢?最早研究这个问题的是植物种群存活率随密度增加而变化的-3/2自疏法则,自疏导致固着生长的植物随密度增加引起的竞争加剧而淘汰较小的个体(Norberg,1988)。随后,研究发现多物种共存群落生产力高于单种生产量,也就是所谓的超产(overyield)现象,这引起了生态学家对物种多样性与生产力二元变量相关关系的关注(Grime,1979)以及对物种共存、多样性维持内在机制的深入探究。基于Lotka-Volterra的种间竞争模型,理论生态学提出了物种共存的适合度机制(fitness)和稳定性机制(stability),提出如此多样的物种之所以能够在有限的空间内共存是因为物种间存在生态位的分异,这尤其是稳态环境中物种得以实现共存的先决和必要条件。另外,环境的波动及不同物种对环境变化的非线性响应的异步性(asynchrony)也是实现物种共存的重要条件(Norberg,1988)。此外,储能效应(storage effect),即物种在适合生长的条件下积蓄资源与能量以供应不适合生长的条件下种群的维持与更新,也是实现物种恢复、更新、建植的重要方式。生态系统中物种共存实现的条件,即多物种共存的优势,一方面多物种共存可以通过物种之间生态位分异的互补性地充分利用生境资源,从而提高生产力(Vandermeer,1972);另一方面,不同植物种对环境条件变化的异步和非线性响应可以缓冲生态系统结构和功能的波动,增强整个系统的稳定性。多物种共存还可以充分发挥不同物种之间的协助性相互作用,减弱竞争作用(Norberg,1988)。然而,在考虑有限空间的资源有限性时,传统生态位理论却面临挑战。中性理论明确强调扩散限制对群落物种多样性的影响,近年来,生态学家对此理论进行了推论和改进。目前,更多的生态学家致力于将二者整合于一体,以更好地理解群落构建机理。除此之外,生物多样性的研究发展出了数学与多模型研究、元分析、生物信息学研究、多因素综合研究等新方法。
2 生态系统的多样性、功能与研究方法
生物多样性对生态系统功能和服务具有重要意义。在物种丰富的生态系统中,理解生物多样性与生态系统功能的关系是一项艰巨的任务(Brophy et al.,2017)。对生物多样性和生态系统功能关系的广泛研究已达成共识(图1),即增加生态系统的生物多样性可提高生态系统维持或增强功能的能力(Finn et al.,2013;Duffy,2009;Bell et al.,2005)。
面对全球生物多样性的下降,人们日益关注生物多样性与生态系统功能之间的关系(Chesson,2000)。生物多样性与生态系统功能和服务之间的关系引起了生态学家的极大兴趣。在过去的 20年里,生态学家进行了大量的理论和实验研究。许多生物多样性——生态系统功能(BEF)实验的结果表明,物种丰富度与生态系统功能之间存在着正向、饱和的关系(Cardinale et al.,2011,2007;Balvanera et al.,2006)。在考虑多种功能和服务时,生物多样性越来越重要(Isbell et al.,2011;Zavaleta et al.,2010)。
多样性研究的出发点就是探究物种多样性与生产力二元变量之间的相关性。一直以来,环境压力和干扰被认为是影响物种共存和多样性维持的主要因素,Grime(1979)提出这两个因子可以被组合为单一的群落性质即最大总生物量,并进一步发展形成了物种多样性与生产力之间的humped-back模型。接下来的30多年,生态学家们在不同生态系统广泛收集数据,试图建立物种多样性与生产力之间普适性的二元变量相关关系。虽然,大量研究发现了正相关、负相关、humped-back、不相关等多种类型的相关性存在,然而,并没有得到二者之间普适性的变化规律,这不得不让生态学家们思考一个问题,即物种多样性和生产力可能受到不同因子的驱动,其背后的作用机制可能各不相同,与其不懈地寻找物种多样性与生产力之间普适性的二元变量相关关系,不如探究调节生态系统属性的多元变量之间的复杂网络联系(Adler et al.,2011),揭示二者表象相关性背后的驱动机理。Grace et al.(2016)采用结构方程模型(structural equation model)构建了包括气候、土壤、干扰、光照等因子在内的模型,分析了多元变量对物种多样性、生产力以及二者之间关系的作用机制。伴随着物种多样性——生产力关系的研究历程,生态学家越来越清晰地认识到仅仅用简单的二元变量关系刻化生态系统结构与功能之间的关系是远远不够的,探究生态系统多元变量之间的联系以及多功能网络之间的权衡关系是大势所趋。
实验技术、统计方法、模型与模拟分析等手段的不断发展促使生态系统多功能研究成为可能,生态学家们着眼于探寻生物多样性与系统多功能之间的内在联系。生物多样性对生态系统功能的有利作用经常用物种丰富度来量化(Spehn et al.,2005)。但是,除了丰富度之外,模拟物种之间和均匀度之间的相互作用,还可以提高我们对多样性驱动的生态系统功能改善的理解(Connolly et al.,2013;Finn et al.,2013;Wilsey et al.,2004)。然而,随着物种丰富度以及相互作用数量的增加,模拟物种相互作用变得越来越困难。近年来,模型分析在研究生态系统功能中得到热切关注,它可以测试生态系统功能如何受到物种类型、物种相互作用和均匀度以及丰富度的影响。然而,在一个物种丰富的生态系统中,可能有太多的物种进行相互作用,无法估计每个系数,如果所有的相互作用系数都是可估计的,它们可能没有任何合理的生物学意义,因此,其在使用中受到制约。Brophy et al.(2017)扩展了多样性——交互模型,用以描述多样性对生态系统功能的影响,并将固定系数和随机效应进行结合,用草地和微生物实验的数据说明了此种方法在测定固定系数中可以改进标准误差,其建模方法比较简约并能提供丰富的信息资源,能够大大减少模拟物种相互作用对生态系统功能的影响所需的系数,同时又不忽略生态系统功能变异性的潜在重要性,帮助我们更好地理解在物种丰富的生态系统中物种间的相互作用与促进生态系统功能复杂性的关系。
3 草地生态系统的管理
3.1 草地生态系统服务功能
生态系统服务功能是指生态系统及其生态过程所形成与维持的、人类赖以生存的环境条件和效用,是指通过生态系统的功能直接或间接得到的产品和服务。草地生态系统的服务功能包括了支撑人类生存环境、调节大气条件、维持生命系统、提供休闲旅游、文化传承等服务功能。草地生态系统可以为畜牧业发展提供优良牧草,具有防风、固沙、保土、涵养水源、调节气候等功能;草地是重要的生物多样性和珍稀动植物物种保护基地,也是重要的药材生产基地;草地生态系统作为重要的天然绿色屏障,具有净化空气、美化环境的作用;草地还具有重要的人文历史价值。
3.2 草地经营的发展简史与管理措施
自有人类以来,为了保证人类的繁衍与种族延续,人类的祖先就开始对草地进行利用。起初以游猎为主,后来随着自然环境的变化、人类认知能力的提高和生产工具的进步,产生了原始的种植业和畜牧业,这是人类社会的巨大进步,从此,人类开始了利用草场的历史。放牧是对草场利用的最原始、最主要的方式。19世纪中期,各国对草地的利用基本处于自然放牧状态,人们对草地只有利用却不进行建设,加上开垦破坏,导致草畜矛盾加剧、草地自然生态失调、自然灾害频发。20世纪初,人类才认识到科学管理与利用草地资源的重要性。20世纪以来,一些发达国家首先采用了草地科学管理和建设措施,如制定相关法律法规、实行轮牧、改良天然草地、建设人工草场、建立国家机构、加强科学研究,使草地利用迈入了现代科学经营的新阶段(李毓堂,2003)。中国对于草场的利用和放牧至少已有三四千年的历史,但是真正对草场进行经营管理的历史较短。中国草地经营科学起步于 20世纪40年代,40—50年代奠定了学科发展的基础;60年代初期学科发展有了新起点;80年代草地经营认识有了新高度;50—80年代为草业学科的形成期。从 2000年至今,草地经营管理的发展进入新的人为科学调控下的优化发展阶段(许庆方等,2005)。
人类为维持草地生态、实现草地的可持续发展也做出了许多努力,例如保护物种多样性(如设立自然保护区),在一定限期内实行轮牧、休牧和围封育种,进行人工草地建设,在景点建立风景名胜区等。
3.2.1 自然保护区
设立自然保护区仍然是保护生物多样性的核心举措(Cumming et al.,2015),是实现草地生物多样性及其生态系统服务的重要途径。近年来,关于自然保护区的系统保护和规划已具备坚实的理论基础,可以使生物多样性保护与其他社会经济目标之间的权衡达到最佳,并通过数学和模型分析的利用,力求对特定物种的保护水平达到理想目标(Kendall et al.,2015)。自然保护区的设置要充分衡量设定范围、空间配置、系统设计、物种名录和成本等因素,如果知道物种的确切位置,那么就可以应用空间优化技术根据保护目标将保护区的位置准确定位,同时尽量减少其他的保护成本。最近,Mackey et al.(2018)提出了一种新型保护区设计策略,将集合种群理论与空间保护的优先度相结合,对减少物种灭绝风险的保护方案进行估计,优化空间配置,通过考虑物种的特征和景观的连通性,使整个物种组合的集合种群持久性达到了最大化。但是,目前,在自然保护区规划和设计中,仍以生物多样性保护为主体,生态系统服务很少能得到充分考虑。因此,如何在保护资源有限的前提下兼顾二者的保护需求、实现生态保护效益的最大化成为热点和难点(房志等,2017)。另外,自然保护区的空间尺度问题仍然是需要探讨的问题。通过研究自然生态规律,为合理开发自然资源,建立自然保护区有利于对草地生态系统特有、珍稀和濒危生物资源进行保护,同时也可实现对特殊自然景观的保护。
3.2.2 人工草地
人工草地作为促进草地生态系统恢复的一种措施,是采用农业技术措施栽培而成的草地,目的是获得高产优质的牧草,以补充天然草地的不足,满足家畜的饲料需要。人工草地面积的多少,常是衡量一个地区或国家畜牧业发达程度高低的重要标志(李凌浩等,2016)。发达国家的畜牧业人工草地的面积通常占全部草地面积的10%~15%以上,西欧、北欧和新西兰已达 40%~70%或更多(胡自治,1995)。中国在人工草地建设上取得了丰硕的成果,尤其在内蒙古自治区和新疆维吾尔自治区建立了许多人工草地。近些年来,人工草地的生态系统服务功能受到了广泛重视,实践证明,进行人工草地建设是促进退化草场生态恢复的快速途径,是提高草地生产力和促进畜牧业发展的重要保障(Foley et al.,2011)。人工草地改变了草地的原有状态,其引起的植被等变化必将引起草地生态系统能量平衡的改变。
3.2.3 轮牧、休牧和围封
放牧改变了植物群落特征、养分有效性、物种多样性,进而影响生产力的形成过程。但是,由于人类超载放牧,引起了草地生态系统生态环境恶化,如草场退化、物种多样性降低、草地生产力下降,限制了畜牧业的发展(韩文祥,2013;Naeem et al.,1997;Tilman et al.,1994)。在发现这些问题之后,人类开始探究新的牧区管理方式,如轮牧、休牧和限期围封,以促进草场生态环境恢复,实现草地可持续发展。轮牧是经济有效利用草地的一种放牧方式,按季节依次轮回或循环放牧的一种放牧方式。休牧是中国主要放牧管理和草地保护措施之一,可以通过自然力的作用,使退化草地的植被与土壤得到恢复,维持多样性和较高的生物量。适度围封往往使植被层的高度、盖度和生物量都得到显著增加,使退化草地得到有效改善。但是,休牧和围封均有期限,否则将会限制草地生产力,不利于草地生态系统的保护和合理利用(Su et al.,2003)。围封作为退化草地恢复措施中常用的方法之一,具有成本低、见效快、操作简单等优势,在草地管理方面极为有效。
4 草地发展中存在的问题
相比于生态学理论上对生态系统多功能的认识和刻化,人们在畜牧业发展的实践中早就认识到草地生态系统多功能及功能之间的潜在联系。但是为了追逐经济效益的最大化,人们还是宁愿在超出草地承载力的条件下盲目发展畜牧业,使生产力功能最大化,以牺牲草地生态系统的其他功能为代价。
4.1 草地生态系统生态失衡
由于人类对草地生态系统的不合理利用,如无节制地开发生物资源、过度放牧、环境污染、外来物种入侵等,致使草地生产力下降、生物多样性锐减、动植物濒临灭绝、草场退化、水土流失严重、自然灾害频繁等生态环境问题加剧,因此草地生态系统失衡。
4.2 草地的多种生态服务功能没有得到开发
随着人类对草地放牧利用的实践以及生态学对草地生态系统结构和功能作用机理的认识的深入,人们越来越清晰地认识到草地的功能不能再盲目追逐高生产力的单一模式下发展了,除生产力功能外,草地生态系统同时还具有生态旅游、水土保持、美化环境、改善气候、维持环境与生命系统平衡等其他非农业用途(谢高地等,2003),评估草地价值不能只依靠经济输出(欧阳志云等,2002)。草地的重要性不仅表现在其经济功能上,而且还体现在生态功能上。我们不能再重蹈“先污染后治理”的老路。最近有关生态系统多功能研究(余兴光等,2006;马克明等,2001)的结果表明,盲目扩大生态系统某方面功能而引起另外功能失衡后,修复系统的代价远远高于提高了某一功能的所得。
4.3 草地生态系统管理滞后
尽管许多年来,草地经营管理取得了一系列成果,但是草地经营管理仍然面临许多严重问题,表现为草地基础建设薄弱、草地管理水平低、牲畜饲养管理水平落后、畜群结构不合理、草畜供求矛盾等。目前的草地管理模式很大程度上基于人类畜牧业发展过程中对草地利用的实践经验,理论发展的滞后严重地制约了草地管理的科学化和可调控性。
5 草地多样性保护、调控和优化草地多功能
草地生态系统作为农业经济发展的重要基地之一,在科学、技术、信息飞速发展的现阶段,迫切需要建立起系统全面、因地制宜、科学预警的管理发展模式,增强人为调控,不再依附靠天吃饭的局面。
5.1 生物多样性保护
草地蕴涵了丰富的生物多样性,然而人口的增长和经济的快速发展正日益对生物多样性造成严重威胁,物种灭绝、物种入侵等问题日益严重。保护生物多样性也即保护生态系统的多功能服务及其稳定性。草地生态系统本身的脆弱性使得草地生物多样性的保护成为维持生态系统多功能发挥的重要举措。
5.2 转变发展方式,实现可持续发展
加强宏观调控,调整产业结构是开发草地生态系统多功能和维持可持续发展的对策之一。实行退耕还林还草,经济发展不再局限于种植业和畜牧业,而应该在适度发展这些产业的同时,利用本地特有的资源发展其他辅助产业,如发展药材基地,通过发展生态、休闲旅游等带动草业、餐饮服务业等相关产业的发展,培植新的经济增长点。同时,要加强科学、文化建设,发挥草原的历史文化传承作用。
5.3 建立科学调控的管理模式
5.3.1 生物多样性信息学
信息技术的发展和开放性数据共享为生物多样性信息的产生、维持、传播和使用带来了革命性的发展,产生了生物多样性信息学(Causey et al.,2004;Bisby et al.,2002;Bisby,2000;Edwards et al.,2000),并且保持着相当乐观的发展前景。环境类型数据是研究生物多样性分布格局、多样性形成与维持机制、物种保育等很多重要生态学问题的基础(张凤麟等,2018)。目前的研究主要是通过生物多样性信息采集、整理、存储、处理和共享过程中的关键技术和手段,探讨生物多样性评价的方法并开发评价工具,研究制订生物多样性数据规范和标准,规划、设计并实施生物多样性信息数据库系统建设,并取得了一系列进展,主要成果是建立了多功能信息库、采集和预测系统。将该学科运用于草地生态系统中,可以通过对草地信息的采集,分类建立数据库,不仅有利于草地生物多样性保护措施的制定,而且可以更简便地预测和评估退化草地生态系统的恢复程度。
5.3.2 系统分析
系统分析方法从系统需求入手,从用户观点出发建立系统用户模型。用户模型从概念上全方位表达系统需求及系统与用户的相互关系。系统分析在用户模型的基础上,建立适应性强的独立与系统实现环境的逻辑结构。系统分析方法的具体步骤包括:限定问题、确定目标、调查研究收集数据、提出备选方案和评价标准、备选方案评估和提出最可行的方案。系统分析在草地生态中已得到应用,在限定草地生态问题和确定要达到的预期效果之后,根据草地生态系统的特性进行变量选择、反馈机制和确立平衡点,通过以上资料建立相应的模型,并对模型进行灵敏度分析。
5.3.3 网络分析
网络分析方法是一种系统、科学、有效的分析方法,它以网络结构为基础,实现对生态系统中物质和能量流动结构的模拟,有利于在时空分布结构上帮助我们对生态系统产生完整的认识。网络分析可以量化生态系统内部各组分之间的联系方式、作用关系,揭示生态系统的整体性和复杂性,已经成为研究生态系统结构、内部变化规律的有效工具(李中才等,2011)。网络分析已经被有效地应用到复合种群的多样性保护、保护区建立等领域。在利用此方法时,一般会涉及流动性、随机性、等级性和灵敏度的分析。目前,国外网络分析在自然保护区应用广泛,尤其是对濒危物种的保护发挥了积极作用,在实现物种保护的同时,也为具体的保护规划和精细评估提供及时、有效的信息。未来研究的重点可能涉及:网络结构的拓扑特性、网络的结构与功能关系、网络的优化算法与控制方法、分析软件功能的不断完善等。
5.4 草地管理应用研究
现今,草地管理应用仅仅依靠理论基础研究是不够的。人类与草地生态系统的关系不可分离,在保护草地资源的同时,其实对草地利用得更多,因此,研究草地生态系统时应该考虑人与草地的关系,应该使草地得到更加合理、充分、有效和长期的健康发展,真正实现生态、经济、社会效益最大化。
草地生态系统是动态平衡系统,这使得系统动力学、网络分析在草地资源的调控管理、草地生物多样性保护中具有广阔的发展前景。在解决草畜矛盾、人地矛盾、经济利益与生态维持等矛盾中,人类需采用动态而非静态的方法来解决这些问题。通过对不同草地生态系统建立相应的模型,在不断调节变量中权衡生态系统多元功能之间的关系,寻找最佳途径,提供未来发展变化趋势的有效预警,使草地系统在科学的人为调控下可持续发展。
ADLER P B, SEABLOOM E W, BORER E T, et al. 2011. Productivity is a poor predictor of plant species richness [J]. Science, 333(6050):1750-1753.
BACHMAN S, BAKER W J, BRUMMITT N, et al. 2004. Elevational gradients,area and tropical island diversity:an example from the palms of New Guinea [J]. Ecography, 27(3): 299-310.
BAILEY J K, SCHWEITZER J A, ÚBEDA F, et al. 2009. From genes to ecosystems:a synthesis of the effects of plant genetic factors across levels of organization [J]. Philosophical Transactions of the Royal Society B, 364(1523): 1607-1616.
BALVANERA P, PFISTERER A B, BUCHMANN N, et al. 2006.Quantifying the evidence for biodiversity effects on ecosystem functioning and services [J]. Ecology Letters, 9(10): 1146-1156.
BELL T, NEWMAN J A, SILVERMAN B W, et al. 2005. The contribution of species richness and composition to bacterial services [J]. Nature,436(7054): 1157-1160.
BISBY F A, SHIMURA J, RUGGIERO M, et al. 2002. Taxonomy, at the click of a mouse [J]. Nature, 418(6896): 367-367.
BISBY F A. 2000. The quiet revolution:biodiversity informatics and the internet [J]. Science, 289(5488): 2309-2312.
BOBBINK R, DURING H J, SCHREURS J, et al. 1987. Effects of selective clipping and mowing time on species diversity in chalk grassland [J].Folia Geobotanica et Phytotaxonomica, 22(4): 363-376.
BREGMAN T P, LEES A C, SEDDON N, et al. 2015. Species interactions regulate the collapse of biodiversity andecosystem function in tropical forest fragments [J]. Ecology, 96(10): 2692-2704.
BROPHY C, DOOLEY Á, KIRWAN L, et al. 2017. Biodiversity and ecosystem function: making sense of numerouss pecies interactions in multi-species communities [J]. Ecology, 98(7): 1771-1778.
BURTON J I, MLADENOFF D J, FORRESTER J A, et al. 2014.Experimentally linking disturbance,resources and productivity to diversity in forest ground-layer plant communities [J]. Journal of Ecology, 102: 1634-1648.
BYRNE M, MACDONALD B. 2000. Phylogeography and conservation of three oil mallee taxa, Eucalyptuskochii ssp.kochii, ssp. Plenissima and E. horistes [J]. Australian Journal of Botany, 48(3): 305-312.
CARDINALE B J, MATULICH K L, HOOPER D U, et al. 2011. The functional role of producer diversity in ecosystems [J]. American Journal of Botany, 98: 572-592.
CARDINALE B J, SRIVASTAVA D S, DUFFY J E, et al. 2006. Effects of biodiversity on the functioning of trophic groups and ecosystems [J].Nature, 443(7114): 989-992.
CARDINALE B J, WRIGHT J P, CADOTTE M W, et al. 2007. Impacts of plant diversity on biomass production increase through time because of species complementarity [J]. Proceedings of the National Academy of Sciences USA, 104(46): 18123-18128.
CARSON W P, PETERSON C J. 1990. The role of litter in an old-field community:impact of litter quantity in different seasons on plant species richness and abundance [J]. Oecologia, 85(1): 8-13.
CAUSEY D, JANZEN D H, PETERSON A T, et al. 2004. Museum collections and taxonomy [J]. Nature, 305(5687): 1106-1107.
CHAPIN F S, WALKER B H, HOBBS R J, et al. 1997. Biotic control over the functioning of ecosystems [J]. Science, 277(5325): 500-504.
CHEN Z Z, WANG S P. 2000. Typical grassland ecosystem of China [M].Beijing: Science Press.
CHESSON P. 2000. Mechanisms of maintenance of species diversity [J].Annual Review of Ecological Systems, 31(1): 343-366.
CLARK C M, TILMAN D. 2010. Recovery of plant diversity following N cessation: effects of recruitment, litter and elevated N cycling [J].Ecology, 91(12): 3620-3630.
CONNOLLY J, BELL T, BOLGER T, et al. 2013. An improved model to predict the effects of changing biodiversity levels on ecosystem function [J]. Journal of Ecology, 101(2): 344-355.
CUMMING G S, ALLEN C R, BAN N C, et al. 2015. Understanding protected area resilience: a multi-scale, social-ecological approach [J].Ecological Applications, 25(2): 299-319.
CURRIE D J. 1991. Energy and large-scale patterns of animal-and plant-species richness [J]. American Naturalist, 137(1): 27-49.
DIAZ S, CABIDO M. 2001. Vive la difference:plant functional diversity matters to ecosystem processes [J]. Trends in Ecology and Evolution,16(11): 646-655.
DUFFY J E. 2009. Why biodiversity is important to the functioning of real-world ecosystems [J]. Frontiers in Ecology and the Environment,7(8): 437-444.
EDWARDS J L, LANE M A, NIELSEN E S. 2000. Interoperability of biodiversity databases:biodiversity information on every desktop [J].Science, 289(5488): 2312-2314.
FACELLI J M, MONTERO C M, LEON R J C. 1988. Effect of different disturbance regimen on seminatural grasslands from the subhumid pampa [J]. Flora, German Democratic Republic, 180(3-4): 241-249.
FACELLI J M, PICKETT S T A. 1991. Plant litter: its dynamics and effects on plant community structure [J]. The Botanical Review, 57(1): 1-32.
FAY P A, PROBER S M, HARPOLE W S, et al. 2015. Grassland productivity limited by multiple nutrients [J]. Nature Plants, 1(7):15080.
FINN J A, KIRWAN L, CONNOLLY J E, et al. 2013. Ecosystem function enhanced by combining four functional types of plant species in intensively managed grassland mixtures: a 3-year continental-scale field experiment [J]. Journal of Applied Ecology, 50: 365-375.
FLYNN D F B, MIROTCHNICK N, JAIN M, et al. 2011. Functional and phylogenetic diversity as predictors of biodiversity-ecosystem-function relationships [J]. Ecology, 92(8): 1573-1581.
FOLEY J A, RAMANKUTTY N, BRAUMAN K A, et al. 2011. Solutions for a cultivated planet [J]. Nature, 478(7369): 337-342.
FRASER L H, JENTSCH A, STERNBERG M. 2014. What drives plant species diversity? A global distributed test of the unimodal relationship between herbaceous species richness and plant biomass [J]. Journal of Vegetation Science, 25(5): 1160-1166.
FRASER L H, PITHER J, JENTSCH A, et al. 2015. Worldwide evidence of a unimodal relationship between productivity and plant species richness [J]. Science, 349(6245): 302-305.
FRIDLEY J D, GRIME J P. 2010. Community and ecosystem effects of intraspecific genetic diversityin g rassland microcosms of varying species diversity [J]. Ecology, 91(8): 2272-2283.
GAMFELDT L, SNALL T, BAGCHI R, et al. 2013. Higher levels of multiple ecosystem services are found in forests with more tree species[J]. Nature Communications, 4: 1340.
GILLISON A N, BIGNELL D E, BREWER K R W, et al. 2013. Plant functional types and traits as biodiversity indicators for tropical forests:two biogeographically separated case studies including birds, mammals and termites [J]. Biodiversity and Conservation, 22(9): 1909-1930.
GRACE J B, ADLER P B, STANLEY H W, et al. 2014. Causal networks clarify productivity-richness interrelations, bivariate plots do not [J].Functional Ecology, 28: 787-798.
GRACE J B, ANDERSON T M, SEABLOOM E W, et al. 2016. Integrative modelling reveals mechanisms linking productivity and plant species richness [J]. Nature, 529(7586): 390-393.
GRIME J P. 1979. Plant Strategies and Vegetation Processes [M]. New York: Wiley Chichester.
HAUTIER Y, NIKLAUS P A, HECTOR A. 2009. Competition for light causes plant biodiversity loss after eutrophication [J]. Science,324(5927): 636-638.
HEILPERN S A, WEEKS B C, NAEEM S. 2018. Predicting ecosystem vulnerability to biodiversity loss from community composition [J].Ecology, 99(5): 1099-1107.
HILLEBRAND H, BENNETT D M, CADOTTE M W. 2008. Consequences of dominance: a review of evenness effects on local and regional ecosystem processes [J]. Ecology, 89(6): 1510-1520.
HOOPER D U, CHAPIN F S, EWEL J J, et al. 2005. Effects of biodiversity on ecosystem functioning:a consensus of current knowledge [J].Ecological Monographs, 75(1): 3-35.
HUGHES A R, INOUYE B D, JOHNSON M T J, et al. 2008. Ecological consequences of genetic diversity [J]. Ecology Letters, 11(6): 609-623.HUGHES A R, STACHOWICZ J J. 2009. Ecological impacts of genotypic diversity in the clonal seagrass Zostera marina [J]. Ecology, 90(5):1412-1419.
HUSTON M A. 1994. Biological Diversity, the Coexistence of Specieson Changing Landscapes [M]. Cambridge: UK: Cambridge University Press: 181-631.
ISBELL F V, CALCAGNO V, HECTOR A, et al. 2011. High plant diversity is needed to maintain ecosystem services [J]. Nature, 477(7363): 199-202.
KENDALL B E, KLEIN C J, POSSINGHAM H P. 2015. The role of scale in designing protected area systems to conserve poorly known species[J]. Ecosphere, 6(11): 1-17.
KREFT H, SOMMER J H, BARTHLOT W. 2006. The significance of geographic range size for spatial diversity patternsin Neotropical Palms[J]. Ecography, 29(1): 21-30.
LANDE R. 1988. Genetics and demography in biological conservation [J].Science, 241(4872): 1455-1460.
LOREAU M, NAEEM S, INCHAUSTI P, et al. 2001. Ecology-biodiversity and ecosystem functioning: current knowledge and future challenges[J]. Science, 294(5543): 804-808.
MACKEY M S, BRODIE J F. 2018. Reserve design to optimize the long-term persistence of multiple species [J]. Ecological Apllications,5(3): 1739.
MOUQUET N, MOORE J L, LOREAU M. 2002. Plant species richness and community productivity: why the mechanism that promotes coexistence matters [J]. Ecology Letters, 5(1): 56-65.
NAEEM S, LI S. 1997. Biodiversity enhances ecosystem reliability [J].Nature, 390(4): 507-509.
NORBERG R A. 1988. Theory of growth geometry of plants and self-thinning of plant populations: Geometric similarity, elastic similarity, and diffirent growth modes of plant parts [J]. American Naturalist, 131(2): 220-256.
OLLINGER S V. 2005. Biodiversity and Ecosystem Function in Forests [J].Ecology, 86(8): 2253-2254.
PENFOUND W T. 1964. Effects of denudation on the productivity of grassland [J]. Ecology, 45(4): 838-845.
QIAN H, RICKLEFS R E, WHITE P S. 2005. Beta Diversity of angiosperms in temperate floras of eastern Asia and eastern North America [J]. Ecology Letters, 8(1): 15-22.
ROSCHER C, SCHUMACHERB J, ANNETT L, et al. 2013. A functional trait-based approach to understand community assembly and diversity-productivity relationships over 7 years in experimental grasslands [J]. Perspectives in Plant Ecology, Evolution and Systematics, 15(3): 139-149.
SCHMITZ O J. 2008. Effects of predator hunting mode on grassland ecosystem function [J]. Science, 319(5865): 952-954.
SCHMITZ O J. 2009. Effects of predator functional diversity on grassland ecosystem function [J]. Ecology, 90(9): 2339-2345.
SMITH M D, WILCOX J C, KELLY T, et al. 2004. Dominance not richness determines invasibility of tallgrass prairie [J]. Oikos, 106: 253-262.
SPEHN E M, HECTOR A, JOSHI J. 2005. Ecosystem effects of biodiversity manipulations in European grasslands [J]. Ecological Monographs, 75(1): 37-63.
SRIVASTAVA D S, CARDINALE B J, DOWNING A, et al. 2009. Diversity has stronger top-down than bottom-up effectson decomposition [J].Ecology, 90(4): 1073-1083.
SU Y Z, ZHAO H L, ZHANG T H. 2003. Influences of Grazing and Exclosure on Carbon Sequestration in Degraded Sandy Grassland,Inner Mongolia, North China [J]. Environmental Science, 24(4): 23-28.TACK A J M, ROSLIN T. 2011. The relative importance of host-plant genetic diversityin structuring the associated herbivore community [J].Ecology, 92(8): 1594-1604.
TAYLOR D R, AARSSEN L W, LOEHLE C. 1990. On the relationship between r/K selection and environmental carrying capacity: a new habitat templet for plant life history strategies [J]. Oikos, 58: 239-250.TILMAN D, DOWNING J A. 1994. Biodiversity and stability in grassland[J]. Nature, 367(27): 363-365.
TILMAN D. 1987. On the meaning of competition and the mechanisms of competitive superiority [J]. Functional Ecology, 1(4): 304-315.
VANDERMEER J H.1972. Niche theory [J]. Annual Review of Ecology and Systematics, 3: 107-132.
VETAAS O R, GRYTNES J A. 2002. Distribution of Vascular Plant Species Richness and Endemic Richness Along the Himalayan Elevation Gradientin Nepal [J]. Global Ecology and Biogeography, 11(4):291-301.
VIOLLE C, NAVAS M L, VILE D, et al. 2007. Let the concept of trait be functional! [J]. Oikos, 116: 882-892.
WAIDE R B, WILLIG M R, STEINERET C F, et al. 1999. The relationship between productivity and species richness [J]. Annual review of Ecology and Systematics, 30: 257-300.
WHITHAM T G, BAILEY J K, SCHWEITZER J A, et al. 2006. A framework for community and ecosystem genetics: from genes to ecosystems [J]. Nature Reviews Genetics, 7(7): 510-523.
WILSEY B J, CHALCRAFT D R, BOWLES C, et al. 2005. Relationships among indices suggest that richness is an incomplete surrogate for grassland biodiversity [J]. Ecology, 86(5): 1178-1184.
WILSEY B J, POLLEY H W. 2004. Realistically low species evenness does not alter grassland species-richness-productivity relationships [J].Ecology, 85(10): 2693-2700.
ZAVALETA E S, PASARI J R, HULVEY K B, et al. 2010. Sustaining multiple ecosystem functions in grassland communities requires higher biodiversity [J]. Proceedings of the National Academy of Sciences USA, 107(4): 1443-1446.
ZHANG R Z, LIN Y L. 1985. The distribution direction of mammalin China and its neighborhood [J]. Acta Zoologica Sinica, 31(2): 187-197.
房志, 徐卫华, 张晶晶, 等. 2017. 基于生物多样性与生态系统服务功能的秦岭山系自然保护体系规划[J]. 生态学报, 37(16): 5334-5341.
韩文祥. 2013. 草原生态恢复与可持续发展存在的问题及对策分析[J].畜牧与饲料科学, 34(7-8): 74-75.
胡自治. 1995. 世界人工草地及其分类现状[J]. 草原与草坪, (2): 1-8.
雷羚洁, 孔德良, 李晓明, 等. 2016. 植物功能性状、功能多样性与生态系统功能:进展与展望[J]. 生物多样性, 24(8): 922-931.
李凌浩, 路鹏, 顾雪莹, 等. 2016. 人工草地建设原理与生产范式[J]. 科学通报, 61(2): 193-200.
李毓堂. 2003. 中国近现代草地管理开发科学技术的长期之后与发展对策[J]. 草业科学, 20(10): 1-7.
李中才, 徐俊艳, 吴昌友, 等. 2011. 生态网络分析方法研究综述[J]. 生态学报, 31(18): 5396-5405.
孟婷婷,倪健,王国宏.2007.植物功能性状与环境和生态系统功能[J].植物生态学报.31(1):150-165.
马克明,孔红梅,关文彬,等. .2001. 生态系统健康评价: 方法与方向[J]. 生态学报. 21(12), 2106-2116.
欧阳志云, 李文华. 2002. 生态系统服务功能内涵与研究进展[M]. 北京: 气象出版社.
谢高地, 鲁春霞, 肖玉, 等. 2003. 青藏高原高寒草地生态系统服务价值评估[J]. 山地学报, 21(1): 50-55.
许庆方, 韩建国, 董宽虎. 2005. 我国草业发展简要回顾[J]. 草业科学,22(4): 1-4.
余兴光,郑森林,卢昌义. 2006. 厦门海湾生态系统退化的影响因素及生态修复意义[J]. 生态学杂志. 25(8): 974-977.
张凤麟, 王昕, 张健. 2018. 生物多样性信息资源. II. 环境类型数据[J].生物多样性, 26(1): 53-65.
猜你喜欢
四川蚕业(2022年2期)2022-11-19
幼儿100(2020年31期)2020-11-18
世界科学技术-中医药现代化(2020年2期)2020-07-25
疯狂英语·初中版(2019年4期)2019-09-10
科学大众(中学)(2019年3期)2019-05-17
汽车观察(2018年10期)2018-11-06
小太阳画报(2018年6期)2018-05-14
临床医药文献杂志(电子版)(2017年11期)2017-05-17
科技知识动漫(2017年1期)2017-02-06
少儿科学周刊·少年版(2015年1期)2015-07-07
