
产气荚膜梭菌α毒素C末端与中和抗原表位的串联表达及免疫保护性分析
2018-09-06 11:21杜吉革彭小兵张秀坤李启红印春生姚文生陈小云
中国兽药杂志 2018年8期
杜吉革,薛 麒,朱 真,彭小兵,张秀坤,李启红,印春生,姚文生,康 凯,陈小云
(中国兽医药品监察所,北京100081)
产气荚膜梭菌是一种重要的人畜共患菌,能引起人类以及多种家畜家禽发病[1],不仅威胁着人类的健康,而且对畜牧业造成了巨大的经济损失[2]。产气荚膜梭菌主要的致病因素是其分泌的外毒素,并通过其产生的毒素发挥致病作用[3]。根据产生4种主要致死性外毒素α(CPA)、β(CPB)、ε(ETX)和ι(CPI)的种类,可将该菌分为A、B、C、D、E五个毒素型[3]。其中,对养牛业危害最大的是A、C和D型。在国外,牛的产气荚膜梭菌病以C、D型菌导致牛的坏死性肠炎或肠原性毒血症为主,而在国内,牛的产气荚膜梭菌病则以A型菌导致的牛“猝死症”为主[4]。病牛往往无任何前驱症状,而突然发作死亡[4-5]。因此,疫苗免疫是防制该病的有效手段。然而,国内还没有商品化的疫苗来预防牛的A型产气荚膜梭菌病。在预防羊的产气荚膜梭菌病方面,虽然产气荚膜梭菌灭活疫苗,起到了一定的效果,但该类疫苗是通过灭活细菌培养物上清而制备的天然类毒素,制备过程冗杂,抗原成分复杂,有效抗原量较低。如大剂量的肌肉注射会破坏牛肉的品质。因此,提供安全、纯净、有效的抗原对未来预防该菌感染具有重要意义。
CPA是由产气荚膜梭菌染色体基因plc编码[6],该基因存在于五个毒素型的菌株中,但在A型毒株中表达水平最高[7]。CPA分为N-末端(1-246,简称CPAN)和C-末端(247-370)两个结构域。其中,CPAN是CPA发挥酶活性的主要区域,而CPAC是毒素与细胞结合的主要区域[8]。研究表明,重组CPA仍存在一定的毒力[9],而通过甲醛灭活后的天然CPA抗原性会明显降低[10-11]。为此,无毒力CPA的研制具有重要的意义。已有的研究证实,CPAC能够对天然CPA起到一定的免疫保护作用[12-14]。此外,Alison等[15]筛选出一株在体内和体外实验中对CPA均具有中和保护作用的单抗,进一步的研究证实,该单抗针对的线性表位NE的氨基酸序列是ARGFAK,位于CPA的第193~198位。由于CPAC蛋白分子量较小,研究者通常选择将CPAC与GST等大分子量标签蛋白融合表达,从而引入了过多无关的抗原成分[13-14]。为了更好发挥CPAC和NE的免疫保护作用,试验根据我国现行A型产气荚膜梭菌标准株(C57-1株)CPAC和NE的编码序列进行密码子优化设计,并将上述两种基因序列分别进行3次重复后串联,经原核系统表达、纯化和鉴定,并对重组蛋白的免疫保护性进行研究。
1 材料与方法
1.1 菌株、质粒、实验动物和试剂 A型产气荚膜梭菌C57-1株、A型产气荚膜梭菌C57-1株天然毒素、A型产气荚膜梭菌毒素抗血清及pET30a(+)(以下简称pET)表达载体均为中国兽医药品监察所基础保障处微生物实验室保存;1.5~2.0 kg普通级健康日本大耳白兔和16~18 g ICR小鼠购自北京维通利华实验动物技术有限公司;感受态细胞Top10和BL21(DE3)购自北京全式金生物技术有限公司;佐剂Montanide ISA 201购自法国seepic公司;蛋白Marker(M1)、Western blot Marker(M2)、Ni-IDA亲和层析介质试剂盒,购自金斯瑞生物科技有限公司;高保真PCR酶KOD购自东洋坊;premix taq version 2.0,购自takara;T4 DNA连接酶、DNA凝胶回收试剂盒购自Promaga公司;限制性内切酶BamHⅠ和XhoⅠ购自NEB公司;抗His标签单抗、Bradford蛋白浓度测定试剂盒,购自碧云天生物技术有限公司;LB培养基购自北京中海生物科技有限公司。
1.2 基因合成及密码子优化 以A型产气荚膜梭菌C57-1 的CPAC和NE3编码基因序列为模版进行优化设计,将二者的3拷贝序列串联后,在串联基因的3'端添加6*His标签蛋白的编码序列,由中美泰和公司合成基因片段GCPAC3NE3。
1.3 原核表达载体的构建 以人工合成的片段为模板,采用引物对1F/1R进行PCR扩增。其中上游引物1F序列为:5'-GGCGGATCCGTTGGTAAGAAC-3',其5'端引入限制性内切酶BamHⅠ位点(下划线部分)及保护性碱基;下游引物1R序列为:5'-GGCCTCGAGTTAGTGGTGATGGT及保护性碱基,其5'端引入限制性内切酶XhoⅠ位点(下划线部分)。PCR体系为50 μL。PCR反应条件为:94 ℃预变性4 min;98 ℃变性10 s,56 ℃退火30 s,68 ℃延伸2 min,共33个循环;最后68 ℃延伸7 min。
回收目的条带,采用BamHⅠ和XhoⅠ进行双酶切,与经过相同酶切处理的pET连接。将连接好的质粒转化Top10感受态细胞,挑取单克隆,提取质粒,进行PCR和双酶切鉴定,将鉴定结果为阳性的质粒送中美太和公司测序,将测序正确的质粒命名为pCPAC3NE3。
1.4 重组蛋白的表达与纯化 将pCPAC3NE3以及pET转化至BL21感受态细胞中,分别在15 ℃和37 ℃条件下用IPTG诱导表达,采用SDS-PAGE检测重组蛋白的表达情况及其可溶性,并采用Western blot方法,以抗His标签蛋白抗体为一抗,对重组蛋白做进一步的鉴定。对以包涵体形式表达的蛋白进行进一步的纯化和复性,具体方法见文献[2]。
1.5 rCPAC3NE3与A型产气荚膜梭菌毒素抗血清的反应 rCPAC3NE3经SDS-PAGE电泳后转印至硝酸纤维素膜(NC膜)上,以A型产气荚膜梭菌毒素抗血清为一抗,HRP标记的山羊抗兔IgG为二抗进行孵育,按照底物显色试剂盒说明书进行显色。
1.6 抗原性分析
1.6.1 免疫程序 选用对A型产气荚膜梭菌毒素的中和效价为0的家兔,进行重组蛋白的免疫,具体免疫方法和程序见文献[2]。
1.6.2 血清中和效价测定 采用血清中和的方法测定兔血清的毒素中和效价,具体操作方法见文献[2]。
1.6.3 攻毒试验 二免后21d,用1个MLD的A型产气荚膜梭菌天然毒素进行攻毒,具体的免疫程序见文献[2],判定试验疫苗的免疫保护效力。
2 结 果
2.1 CPAC3NE3串联基因的原核表达载体的成功构建
采用引物1F/1R进行PCR扩增,扩增片段经酶切后克隆至pET载体上。获得重组质粒的双酶切电泳结果如图1所示,酶切后出现大小约5 kb的载体DNA片段,以及大小约1203 bp的目的基因片段,与预期相符。测序结果表明,插入的外源基因序列正确,将此质粒命名为pCPAC3NE3。
2.2 CPAC3NE3串联基因的原核表达与纯化 将pCPAC3NE3及pET分别转化BL21(DE3)并诱导表达。SDS-PAGE和Western blot结果显示,在15 ℃诱导16 h和37 ℃诱导4 h 两种条件下表达的重组蛋白均以包涵体的形式表达,分子量约为55 kD,且均能与抗His标签抗体发生反应,与预期相符(图2)。综合考虑蛋白表达量和诱导时间,选择37 ℃诱导4 h的条件进行后续的诱导。如图3所示,按照Ni-IDA琼脂糖凝胶说明书对重组蛋白进行纯化,收集纯度较高的Lane11-12洗脱液进行透析和复性,最终获得的rCPAC3NE3蛋白浓度为0.671 mg/mL。

a:重组蛋白表达的SDS-PAGE鉴定;b:重组蛋白与抗His单抗反应的Western blotM1: 蛋白Marker;M2: Western blot Marker;PC1: BSA (1 μg);PC2: BSA (2 μg);pET: pET 37 ℃、4 h诱导的细胞裂解物;1: pCPAC3NE3,15 ℃、16 h诱导的细胞裂解物;2: pCPAC3NE3,37 ℃、4 h诱导的细胞裂解物;pET1: pET 37 ℃ 4 h诱导的细胞裂解上清;pET2: pET 37 ℃ 4 h诱导的细胞裂解沉淀;3: pCPAC3NE3 15℃ 16 h诱导的细胞裂解上清;4: pCPAC3NE3 15 ℃ 16 h诱导的细胞裂解沉淀;5: pCPAC3NE3 37 ℃ 4 h诱导的细胞裂解上清;6: pCPAC3NE3 37 ℃ 4 h诱导的细胞裂解沉淀a.The identification of recombinant protein expression by SDS-PAGE; b: The identification of recombinant protein with anti-His monoclonal antibody by Western blot M1: protein Marker;M2: Western blot Marker;PC1: BSA (1 μg);PC2: BSA (2 μg);pET: The cell lysates of pET induced with IPTG for 4 h at 37 ℃; 1: The cell lysates of pCPAC3NE3 induced with IPTG for 16 h at 15 ℃; 2: The cell lysates of pCPAC3NE3 induced with IPTG for 4 h at 37 ℃;pET1:The supernatant of cell lysates of pET induced with IPTG for 4 h at 37 ℃;pET2:The precipitation of cell lysates of pET induced with IPTG for 4 h at 37 ℃; 3:The supernatant of cell lysates of pCPAC3NE3 induced with IPTG for 16 h at 15 ℃; 4: The precipitation of cell lysates of pCPAC3NE3 induced with IPTG for 16 h at 15 ℃; 5: The supernatant of cell lysates of pCPAC3NE3 induced with IPTG for 4 h at 37 ℃;6: The precipitation of cell lysates of pCPAC3NE3 induced with IPTG for 4 h at 37 ℃图2 rCPAC3NE3的原核表达与鉴定Fig 2 Prokaryotic expression and identification of rCPAC3NE3
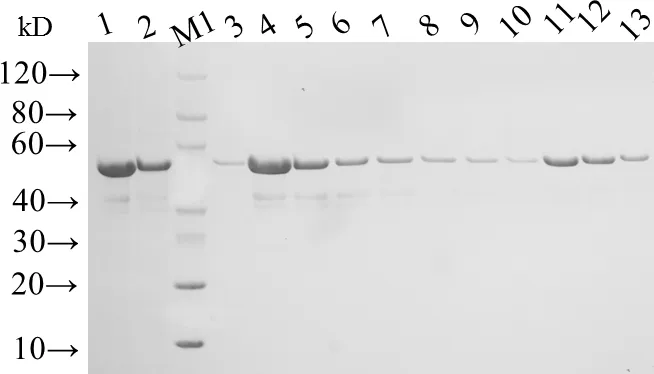
M:蛋白Marker; 1:包涵体溶解离心后上清;2:上清同Ni-IDA孵育后流出液;3-10:50 mmol/L Imidazole的洗脱液;11-12:100 mmol/L Imidazole的洗脱液;13:300 mmol/L Imidazole的洗脱液M: protein Marker; 1: The supernatant of dissolved inclusion bodies; 2: The flow-through from Ni-IDA resin after incubated with supernatant;3-10: The elution from Ni-IDA resin washed with elution buffer (contain 50 mmol/L Imidazole); 11-12: The elution from Ni-IDA resin washed with elution buffer (contain 100 mmol/L Imidazole);13: The elution from Ni-IDA resin washed with elution buffer contain 300 mmol/L Imidazole)图3 rCPAC3NE3的纯化Fig 3 Purification of CPAC3NE3
2.3 rCPAC3NE3与A型产气荚膜梭菌毒素抗血清的鉴定 rCPAC3NE3经SDS-PAGE后转移到NC膜上进行Western blot,结果如图4所示,rCPAC3NE3与A型产气荚膜梭菌毒素抗血清发生明显反应。
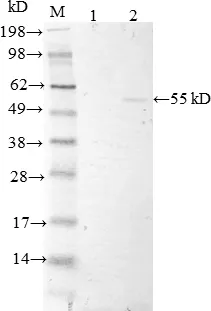
M: 蛋白Marker;1. pET 37 ℃ 4 h诱导的细胞裂解物;2: 纯化后的rCPAC3NE3M: Protein Marker; 1: The cell lysates of pET induced with IPTG for 4 h at 37 ℃;2: rCPAC3NE3 after purification图4 rCPAC3NE3与A型产气荚膜梭菌抗毒素血清的反应Fig 4 Interaction of rCPAC3NE3 with antitoxin serum of Clostridium perfringens type A
2.4 rCPAC3NE3的免疫原性分析
2.4.1 抗rCPAC3NE3兔血清的毒素中和抗体效价测定
经血清中和法测定,以rCPAC3NE3免疫家兔后,每毫升的一免抗血清可中和40个MLD的A 型产气荚膜梭菌毒素;每毫升的二免抗血清可中和80个MLD的A 型产气荚膜梭菌毒素,而佐剂对照组的兔血清对毒素无中和作用。
2.4.2 rCPAC3NE3免疫对兔的免疫保护结果 在二免后21 d,对所有rCPAC3NE3免疫组和对照组的家兔,经耳缘静脉注射1个MLD的A型产气荚膜梭菌天然毒素,结果对照组家兔在攻毒后5 d内全部死亡,rCPAC3NE3免疫组家兔全部健活,未见任何不良反应,保护率达100%。
3 讨 论
随着研究的深入,与产气荚膜梭菌密切相关的主要致死性外毒素的结构和致病机理越来越清晰,致死性外毒素的部分无毒区域(CPA、CPB以及CPI的C末端)或者无毒突变体(ETX 和θ毒素(PFO)[2])作为亚单位疫苗抗原已经被证实能够有效地中和相应的毒血症。作为A型产气荚膜梭菌的主要致死性毒素,CPA的减毒乃至无毒研究工作成为了众多研究者的方向。虽然对CPA发挥功能的关键氨基酸位点进行单点突变能够实现减毒的目的[16-18],但这些突变体的免疫原性还没有得到充分的研究。此外,单个氨基酸突变的CPA在未来基因工程疫苗大规模生产中存在一定的生物安全隐患。对CPA的两个区域免疫保护性分析的结果表明,单独的CPAC具有一定的免疫保护作用,而单独的CPAN却无免疫保护作用。然而,针对CPA第193~198位氨基酸的单抗已被证实具有良好的中和保护作用,而该表位则处于CPAN内[15]。这可能由于单独的CPAN分子量过小,用与GST融合的蛋白免疫动物,实际的抗原量不足。为此,试验将CPAC和线性表位NE分别进行3次重复后串联进行融合表达,不仅增加了有效抗原量,也增加抗原蛋白整体的分子量,更有利于增强抗原蛋白的抗原性。
对非可溶形式表达的重组蛋白进行纯化和复性后获得rCPAC3NE3,进一步的研究发现,rCPAC3NE3具有较好的免疫原性。在实际生产中,以包涵体形式表达的重组蛋白在纯化中可显著降低产物中内毒素的含量,这是rCPAC3NE3作为制备疫苗用抗原的优势之一。然而,在前期的研究中我们发现可溶性表达的重组ETX免疫原性明显高于非可溶性表达的重组蛋白。为此,如何提高rCPAC3NE3的可溶性表达量并验证其免疫原性,将是后续研究的重要方向。
猜你喜欢
当代水产(2022年2期)2022-04-26
猪业科学(2022年1期)2022-03-24
动物医学进展(2022年1期)2022-02-12
环境卫生工程(2021年5期)2021-11-20
中国饲料(2021年17期)2021-11-02
当代水产(2021年2期)2021-03-29
兽医导刊(2021年15期)2021-01-10
天津建设科技(2020年5期)2020-11-12
Defence Technology(2019年6期)2020-01-07
中国当代医药(2019年36期)2019-02-11
