
光照、温度及盐度对卤虫(Artemia salina)耗氧率与排氨率的影响
2019-01-16 12:26丁红霞景素琴范佩丽刘少贞
盐科学与化工 2019年1期
丁红霞,景素琴,范佩丽,许 涛,魏 伟,刘少贞,宋 晶*
(1. 南风化工集团股份有限公司技术中心,山西 运城 044000;2. 山西农业大学动物科技学院,山西 太谷 030801)
呼吸和排泄是代谢的两个重要组成部分,研究各种环境因子对耗氧和排氨的影响,有利于了解水生动物代谢活动的变化规律, 是生态能量学和营养生理学研究的主要内容,也是机体生理代谢和生长状况的重要指标。耗氧率的大小及变化在很大程度上反映其代谢水平的高低及变化规律。排氨率则反映水生动物的代谢特征、动物自身的生理状况和营养状况以及对外界环境条件的适应能力。
卤虫是一种生活于高盐度水体中的甲壳纲(Crustacea)动物,隶属于鳃足亚纲(Branchiopoda),无甲目(Anostraca)卤虫科(Artemidae),在世界沿海和内陆盐湖中广泛分布。自1933 年Seale最先将卤虫无节幼体作为鱼苗的开口饵料以来,卤虫卵广泛应用于海水鱼、对虾和河蟹育苗中,其年需求量也从刚开始的几吨到目前的上万吨[1]。卤虫成虫也是一种重要的水产饵料,其蛋白质含量达60%以上,富含多种必需氨基酸、类胡萝卜素、天然诱食剂等[2]。但近30年来,围海造陆、盐湖面积缩减、环境恶化、气候剧变、盐化工的发展,均使得产量原本就极不稳定的卤虫和虫卵资源更显稀缺[3]。自20世纪80年代末,研究人员就一直在探索卤虫的养殖技术,并取得了很多重要成果。养殖替代捕捞,粗养向精养转变是卤虫养殖行业发展的必然趋势。
卤虫和其他其他甲壳动物一样,均为“排氨型动物”,其呼吸生理研究一直受到国内外研究者们的重视。目前,主要的研究工作多围绕具有生态指示功能或具较高经济价值的甲壳类展开。之前的研究结果表明,甲壳动物耗氧率和排氨率主要受到温度[4-5]、光照[6]、基础氧量[7-8]、盐度[9-11]、纬度[12]、金属离子[13-14]等环境因子的影响;不同的发育时期[15-17]、蜕壳[18]、选择性穴居[19]等因子也会影响甲壳动物耗氧率和排氨率。但对卤虫(Artemia salina)的此类研究尚属空白,卤虫呼吸代谢的特性亟待确定。在生产周期中,水体溶解氧的饱和度随水温升高而降低,而水体中的化学耗氧量、养殖动物和其它生物的耗氧量则随温度升高而增加。水中氧气消耗,代谢产物积累诱发的水质恶化,可能会使孤雌生殖的卤虫向两性生殖转变,这将对以收获鲜卤虫为主的养殖生产造成不利影响。故该研究对主要环境因子对卤虫呼吸代谢的影响进行初探,旨在掌握卤虫的相关生理特征,为推动卤虫的集约化养殖积累原始资料。
1 材料和方法
1.1 试验材料
实验动物用卤虫均为成体,于2017-07取自南风化工集团卤虫养殖基地,选择三种不同盐度(14.0%、16.0%、19.4%)养殖水体中的卤虫进行耗氧率和排氨率测定。窒息点、温度、光照对耗氧率和排氨率的影响,均在盐度14.0%测定。
1.2 窒息点的测定
卤虫窒息点的测定以125 mL具塞磨口试剂瓶为呼吸瓶。选用的卤虫均为成体,并在不同温度(20 ℃、25 ℃、30 ℃)间进行比对。每隔30 min观察一次卤虫的行为和存活情况,当半数受试动物窒息时,立即连同受试动物一起固定,以碘量法测定溶氧值[20],作为卤虫的窒息点。
1.3 耗氧率、排氨率的测定方法
耗氧率、排氨率的测定采用容积1 100 mL的褐色广口瓶作为呼吸室,每个实验单元,设置2个对照、3个平行。先将瓶中注满采自养殖区、并经过机械增氧的高氧盐水(≥6 mg/L),加入一定湿重的卤虫,加盖排空气体,水下密封实验,1 h后,以隔膜电极法[20]测定溶氧(台湾衡欣,AZ-8403),以改良次溴酸盐氧化法[21]测定氨氮,分别用于计算耗氧率和排氨率。
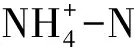
1.4 数据统计
实验数据采用平均值±标准差表示,利用Excel 2010和SPSS 19.0软件进行数据统计分析。各组数据在进行方差分析之前,均先通过正态性和方差齐性检验,组间差异进行单因素方差分析(one-way ANOVA),采用Duncan’s多重比较,显著水平为P<0.05,极显著水平为P<0.01。温度对耗氧率和排氨率的影响,采用二项式回归分析。
2 结果
2.1 不同温度下卤虫的窒息点变化
在测定卤虫窒息点的过程中,其活动规律与在自然条件下差异明显(见表1)。但在试验过程中,与死亡状态差异不明显,故需要细心观察。窒息点试验结果表明,卤虫在20 ℃、25 ℃和30 ℃的窒息点分别为1.15 mg/L、1.32 mg/L和1.69 mg/L。卤虫的窒息点与温度高低密切相关,且窒息点随着温度的升高而升高,符合生物代谢规律(见图1)。

表1 试验过程中卤虫的游泳行为变化Tab.1 The changes of swimming behavior in A. salinaduring the course of the experiment

图1 三种不同温度下卤虫的窒息点Fig.1 The asphyxiant point of A. salina at 3 different temperatures
2.2 光照对卤虫耗氧率和排氨率的影响
选取三组卤虫,在相同温度(25.0 ℃)、盐度(14.0%)但不同光照条件(0 lx、200 lx、1 000 lx)下,测定其耗氧率和排氨率,同时设置两个对照实验。实验结果显示:卤虫的耗氧率在0 lx、200 lx、1 000 lx条件下的数值分别是1.134 mg/g·h±0.153 mg/g·h、1.480 mg/g·h±0.168 mg/g·h、1.552 mg/g·h±0.152 mg/g·h,显然,卤虫的耗氧率随着光照的增强而提高,有光照和无光照之间的耗氧率差异显著,不同光照强度之间的耗氧率差异不显著;排氨率在0 lx、200 lx、1 000 lx条件下的数值分别是155 μg/g·h±16.6 μg/g·h、176 μg/g·h±17.5 μg/g·h、198 μg/g·h±23.4 μg/g·h,可以看出卤虫的排氨率与光照强度成正相关关系,各组之间的排氨率差异显著。3种盐度下卤虫的耗氧率和排氨率见图2。

图2 3种盐度下卤虫的耗氧率和排氨率Fig.2 The oxygen consumption rate and ammonia excretion rate in A. salina at 3 different salinities
2.3 温度对卤虫耗氧率和排氨率的影响
结果表明,在20 ℃~40 ℃范围内,卤虫的耗氧率先上升,后下降,约在30 ℃~35 ℃达到峰值(见图3A);在20 ℃~40 ℃范围内,卤虫的排氨率持续上升(见图3B)。对其变化规律进行多种回归分析,都有较好程度的拟合,但是二项式能够获得最高的回归系数(R2)值,分别得出耗氧率和排氨率回归方程为:OCR=-0.133 1x2+0.964 5x-0.190 4,R2=0.918 5;AER=-4.892 9x2+63.007x+26.3,R2=0.995 8。
2.4 盐度对卤虫耗氧率和排氨率的影响
选取3种不同盐度水体的卤虫,分别于常温、常光下测定其耗氧率排氨率,结果表明,高盐度组(19.4%)的耗氧率和排氨率在数值上均大于中(16.0%)、低(14.0%)度组,但差异不显著(见图4)。可能与卤虫具有良好的盐度长期适应性和极强的稳态调节能力相关。相比盐度驯化,盐度的骤变往往更能引起甲壳类的呼吸、排泄变化。

图3 温度对OCR和AER的影响Fig.3 Effect of temperature on OCR and AER

图4 盐度对OCR和AER的影响Fig.4 Effect of salinity on OCR and AER
3 讨论
3.1 不同温度下卤虫的窒息点变化
甲壳动物的窒息点研究,多服务于经济甲壳类苗种运输,南美白对虾(Litopenaeusvannamei)仔虾、罗氏沼虾(Macrobrachiumrosenbergii)仔虾和中华绒螯蟹(Eriocheirsinensis)各期幼体的窒息点均有研究。由于天然卤虫或较为粗放的卤虫养殖多是浅池养殖,养殖区域都在空旷的海滩、盐湖、池沼,风力增氧作用非常强,不易发生整体缺氧,但精养池水中氧气消耗过大,代谢产物积累诱发的水质恶化,可能会使孤雌生殖的卤虫向两性生殖转变,这将对以收获鲜卤虫为主的养殖生产造成不利影响。研究表明,卤虫在20 ℃、25 ℃和30 ℃的窒息点分别为1.15 mg/L、1.32 mg/L和1.69 mg/L,窒息点与温度呈现正相关,而且不同温度组间差异显著,显示出与之前大多研究结果相同的规律,即:在生理适温范围内,窒息点与温度正相关。可以推论,在卤虫养殖过程中,若遇高温,需防低氧;在卤虫运输过程中,低温运输能够通过降低代谢而提高存活率,要安排好运输密度和运输时间。与其他相似体型的甲壳动物相比,卤虫代谢高,自主运动能力差,没有其他辅助的呼吸方式,更容易被风吹到池塘一侧,造成短时间密度过高,局部缺氧,迫近窒息点而大量死亡。
卤虫对低氧有一定的抵抗力,和大多数甲壳类一样,其窒息点随温度的升高而增长[15-16],但卤虫属于呼吸调节型生物还是呼吸适应型生物需要进一步研究。
3.2 光照对卤虫耗氧率和排氨率的影响
光照对水生生物的耗氧率和排氨率也有一定影响。在对海蜇(Rhopilemaesculenta)窒息点的研究中发现,受到“水呼吸”的影响,光照组比无光组耗氧更快[21]。光照对中国对虾(Fenneropenaeuschinensis)稚虾的消化生理研究中发现,光强比光色和光周期等参数影响更大[22]。光周期对卤虫的生长无影响,但弱光(6 h/d)更有利于卤虫的存活[23]。
3.3 温度对卤虫耗氧率和排氨率的影响
口虾蛄(Oratosquillaoratoria)的耗氧率随季节温度变化[24]。在12 ℃以下可低温麻醉日本对虾(Penaeusjaponicus),即使操作处理也不会引起耗氧率升高[10]。在适宜的温度范围内,卤虫的耗氧率、排氨率也随温度升高而上升(见图3)。
作为外温动物,卤虫的特殊动力作用较小(special dynamic action,SDA)。耗氧率、排氨率都反应的是卤虫的总代谢情况,主要包括基础代谢和活动代谢两部分。在适温范围内,基础代谢和活动代谢提高,会导致耗氧率、排氨率提高;但接近生理上限的高温,会降低活动代谢并破坏基础代谢。氨氮的毒性效应,会在低溶氧、高温时表现更为明显,所以,在卤虫养殖过程中,要注意监控水质和密度,注意适时采收。由于变温动物的代谢水平随温度的升高而提高,故其耗氧多少与水温高低呈正相关。另外,在实际生产中,水体溶解氧的饱和度随水温升高而降低,而水体中的化学耗氧量、养殖动物和其它生物的生物耗氧量则随温度升高而增加。因此,在高温季节应经常清理饲养环境、适当降低放养密度;在运输卤虫活体时,应尽量避开高温天,控制数量,保障存活率。同类型研究采用的回归方法各有不同,理论上讲,耗氧率、排氨率与温度的关系符合指数回归,但选择回归系数更大的二项式回归能更加精准地反映变化趋势,分别得出耗氧率和排氨率回归方程为:OCR=-0.133 1x2+0.964 5x-0.190 4,R2=0.918 5;AER=-4.892 9x2+63.007x+26.3,R2=0.995 8。
3.4 盐度对卤虫耗氧率和排氨率的影响
渗透压的变化对广盐性生物的呼吸生理有着重要影响。V. Aarset等对极地的一种钩虾(Gammaruswilkitzkii)的代谢进行研究发现,当盐度低时,需要增加能量供应以进行渗透压调节[9]。盐度过高或过低会导致刀额新对虾(Metapenaeusmonoceros)能耗增加,而在盐度快速变化时这种趋势尤为明显[25]。日本对虾(Penaeusjaponicus)温度和盐度对代谢交互作用时,发现随着盐度从15‰上升到25‰,耗氧率降低,继续上升到35‰,则差异不显著;在各个温度组,盐度从15‰上升25‰,再到35‰,排氨率都有所减少[10]。可见广盐性物种的耗氧率和排氨率与其调节体内稳态的程度密切相关,越是接近其常规生态环境的,耗能越小,反之亦然。除了盐度的变化范围,其变化速度也有一定影响,骤然变化,则会快速反馈,长期驯化,可能变化不大。研究中选择的3种盐度的水,都来自于不同的卤虫养殖池,统计结果指出组间差异不显著可能与其已经适应其所在的盐度有关,而且卤虫适应盐度的能力较强,在设置组别时,应该加大组距。尽管如此,在19.4%的高盐度组,其耗氧率和排氨率的均值还是要高于其他两组,根据结果推断,卤虫养殖在盐度较低的组别可能会获得更高的产量。
实验结果表明,选取3种不同盐度水体的卤虫,分别于常温、常光下测定其耗氧率排氨率,高盐度组(19.4%)的耗氧率和排氨率在数值上均大于中(16.0%)、低(14.0%)盐度组,但差异不显著,可能与卤虫具有良好的盐度长期适应性和极强的稳态调节能力相关。对于有一定盐度适应性的生物,要正确把握耗氧率与盐度变化的关系就必须在实验前给予动物足够的驯化时间,并了解动物移入呼吸器后耗氧率的时间变化。相比盐度驯化,盐度的骤变可能更能引起甲壳类的呼吸、排泄变化。
猜你喜欢
黑龙江科学(2021年22期)2021-01-13
天津科技大学学报(2018年5期)2018-10-23
水产科学(2018年3期)2018-06-04
中国盐业(2017年4期)2017-07-19
中成药(2016年4期)2016-05-17
河北渔业(2015年7期)2015-07-21
中国新技术新产品(2015年3期)2015-07-19
中国洗涤用品工业(2015年5期)2015-02-28
河北渔业(2015年1期)2015-01-19
河北渔业(2014年3期)2014-07-08
