
新疆野苹果次生代谢产物对虫害胁迫的响应
2019-07-18 00:56艾沙江买买提朱燕飞韩立群王继勋
新疆农业科学 2019年4期
梅 闯,闫 鹏,艾沙江·买买提,朱燕飞,马 凯,韩立群,王继勋
(新疆农业科学院园艺作物研究所/农业部新疆地区果树科学观测试验站,乌鲁木齐 830091)
0 引 言
【研究意义】新疆野苹果(Malussieversii),分布于天山山脉,同源性高且最原始[1],其花、果实、营养成分、生长习性具有丰富的多样性[2],遗传基因丰富可促进苹果品种的选育[3]。苹果小吉丁虫(AgrilusmaliMats.),属鞘翅目,是苹果毁灭性钻蛀害虫,其幼虫隐藏于树干或枝干皮层内盘旋蛀食,致使木质部和韧皮部分离,造成枝干枯死,为害严重时造成果树整株死亡[4]。中国多省份零星分布为害,1995年在新疆发现,并入侵至天山野果林,快速繁殖并最终暴发成灾[5],被我国定为高度危险有害生物,该虫害枯死野苹果达到600多公顷[6]。栽培苹果是世界上最重要的水果之一,具有自交不亲和,无性繁殖的特点[7],这些特征导致新疆野苹果重要基因消亡后,将不可再生。研究新疆野苹果次生代谢物对虫害胁迫的响应,对保护利用新疆野苹果资源有重要意义。【前人研究进展】昆虫取食寄生植物后,会诱导植物产生系统性的抗虫响应,表现为受损植物细胞信号输入,随后进行信号转导(钙离子通量,磷酸化级联和激素串扰)和信号处理(转录组,蛋白质组和代谢组的重构),随后发生信号响应(防御化合物,蛋白质和次级代谢产物的产生)[8, 9]。次生代谢物质是复杂的分支代谢途径产物,不直接参与维持植物的生长发育和生殖有关的基础生化活动,但可作为信号分子,或直接参与化学防御,包括酚类化合物、萜类化合物、含氮有机碱类化合物、硫代葡萄糖苷等[10]。同时可以利用植物自身的防御代谢物质合成植物源杀虫剂消灭害虫[11]。拟南芥中研究表明,硫代葡萄糖苷构成了植物-昆虫互作主要的防御性状,作为对昆虫取食的响应,MYR催化硫代葡萄糖苷的氰基化以释放毒性化合物,例如,腈,异硫氰酸酯和硫氰酸甲酯[12]。柳枝稷在受到蚜虫侵染后,其体内的ROS和多种次级代谢产物发生显著变化,并伴随着NAC,WRKY和MYB转录因子的上调表达[13]。研究发现,不同品系杨树咖啡酸、肉桂酸、丁香酸对青杨脊虎天牛(Xylotrechusrusticus)的驱避效果明显[14]。另外黄酮类物质也对昆虫有毒性,可以影响昆虫的行为和代谢,或破坏昆虫的正常代谢过程,对其生长、发育和繁殖均有明显的抑制作用[15]。【本研究切入点】研究新疆野苹果受到虫害诱导前后某些次生代谢物质的变化情况及其产生规律。【拟解决的关键问题】以经过前期鉴定稳定遗传的抗虫株系为材料,利用液相色谱-质谱(LC-MS)联用技术,定性定量测定次生代谢物质含量变化趋势,研究新疆野苹果(Malussieversii)受到虫害胁迫诱导后相关次生代谢物质的变化及其产生规律,为保护利用新疆野苹果资源提供理论依据。
1 材料与方法
1.1 材 料
2011~2016年在实地调查新疆新源县苹果小吉丁虫疫区新疆野苹果资源,总计调查面积2 500余hm2,新疆野苹果受苹果小吉丁虫危害严重(干枯死枝率达到90%以上),同海拔同一地点有5株新疆野苹果受害较轻(干枯死枝率低于10%,且枝干生长旺盛)[16]从表型性状分析该类单株可能具备一定的抗虫能力,选定一株为害等级最低植株定为抗虫株系[6]。在具有抗虫株系新疆野苹果周边选取1株具有代表性的受侵害单株作为对照株系,树龄在45~60 a。基于广泛靶向代谢组技术平台的代谢分析,检测分析16个抗虫代谢物,对代谢物的定量和差异分析,揭示新疆野苹果在不同处理条件下抗虫代谢物的代谢机制。表1
表1 检测代谢物种类及数量
Table 1 Detecting the type and quantity of metabolites

ID英文名称English name中文名称Chinese NameMas01Theophylline茶碱Mas02Syringic acid丁香酸Mas03Chlorogenic acid绿原酸Mas04Vanillic acid香草酸Mas05Ferulic acid阿魏酸Mas06Caffeic acid咖啡酸Mas07p-Coumaric acid4-香豆酸Mas08Benzoic acid苯甲酸Mas09Quercetin3-O-a-L-rhamnoside (Quercitrin)槲皮素 3-O-鼠李糖苷 (槲皮苷)Mas10Quercetin槲皮素Mas11aloperine苦豆碱Mas12Theobromine咖啡碱Mas13Gramine芦竹碱Mas14Salicylic acid (SA)水杨酸Mas15trans-Cinnamic acid反式肉桂酸Mas16Rutin芸香甙
1.2 方 法
1.2.1 侵染处理
根据昆虫危害习性,苹果小吉丁虫寄生于苹果枝条韧皮部,虫卵于上一年产于韧皮部,次年5月幼虫开始为害,至7月为高峰,持续到9月。选择经过多年鉴定,抗虫表现稳定的新疆野苹果单株。根据苹果小吉丁虫生育期,分别对两种处理进行幼虫侵染(模拟幼虫孔道,接入成龄幼虫),采集样品,分别是侵染后10 d(机械伤害愈合且接入幼虫取食距伤口>3 cm处),采集抗虫株系与对照株系的健康部位和侵染部位韧皮部50 g,立即在液氮中冷冻并储存在-80℃冰箱中,每个株系3个生物学重复,共计12个样品。
1.2.2 样品提取
取出超低温冷冻保存的生物材料样本,利用研磨仪(MM 400, Retsch)在30Hz条件下研磨1 min,称取100 mg的粉末,利用80%甲醇1.0 mL于4℃提取过夜,期间涡旋3次,使提取更为充分。提取后,12 000 g离心15 min,吸取上清,35℃加热模式下氮气吹干,用100 μL 30%甲醇水复溶样品,涡旋使目标物质充分溶解,12 000 g离心15 min,吸取上清,并保存在进样瓶中用于LC-MS分析。
1.2.3 色谱质谱采集条件
参照Pan、Francescato和Wojakowska[17-19]的方法,采用液相色谱串联质谱(LC-MS)分析方法测定次生代谢物含量。所有的化学试剂都是分析纯或色谱纯的制剂。化学标准品从BioBioPha公司(http://www.biobiopha.com/)与美国Sigma-Aldrich公司购买(http://www.sigmaaldrich.com/united-states.html)。分析条件主要包括:(1)色谱柱:WatersACQUITY UPLC HSS T3 C18 1.8 μm,2.1 mm ×100 mm。(2)流动相:水相为超纯水(加入0.1%的甲酸),有机相为乙腈(加入0.1%的甲酸);(3)洗脱梯度,水:乙腈,0 min为95∶5 V/V,11.0 min为5∶95 V/V,12.0 min为5∶95 V/V,12.1 min为95∶5 V/V,15.0 min为95∶5 V/V;(4)流速为0.4 mL/min;(5)柱温为40℃;(6)进样量为5 μL。样品经过色谱分离后进入到质谱进行分析。
配置不同浓度的标准品溶液,进行质谱分析,其中,标准品浓度包括0.1、0.2、0.5、1.0、2.0、5.0和10.0 μg/mL等。
1.3 数据处理
采用 Microsoft Excel 2010软件对数据进行整理,使用 SPSS 17.0 软件进行方差分析(P<0.05)及标准误的计算。绘图用 origin 8.5 软件。
2 结果与分析
2.1 新疆野苹果自然条件下次生代谢组分差异
研究表明,此次靶向代谢检测分为四个大类,分别是羟基肉桂酰衍生物(图1-A)、生物碱(图1-B)、黄酮醇(图1-C)和其它物质(图1-D)。
羟基肉桂酰衍生物中对照株系香豆酸含量显著高于抗虫株系,含量分别为2.2×104和0.5×104μg/mL,差异倍数达到4.4。丁香酸含量高于抗虫株系42.3%,未达到显著差异。其中反式肉桂酸、咖啡酸、香草酸、阿魏酸含量变化不大;生物碱含量中,抗虫株系普遍低于对照株系,其中苦豆碱含量最低,分别是3.9×103和3.1×103μg/mL,咖啡碱含量最高,达到1.1×106和9.5×105μg/mL,对照株系高于抗虫株系16.9%。芦竹碱和茶碱含量差异均不显著。
黄酮醇含量测定中,各组分含量呈现不同差异变化趋势,其中抗虫株系的芸香苷含量显著高于对照株系,含量分别为9.8×106和4.4×106μg/mL,差异倍数为2.3。抗虫株系的榭皮苷含量高于对照株系,分别达到2.5×107和2.1×107μg/mL;在检测的其它组分中,差异均不显著,其中绿原酸含量最高,苯甲酸含量最低。图1
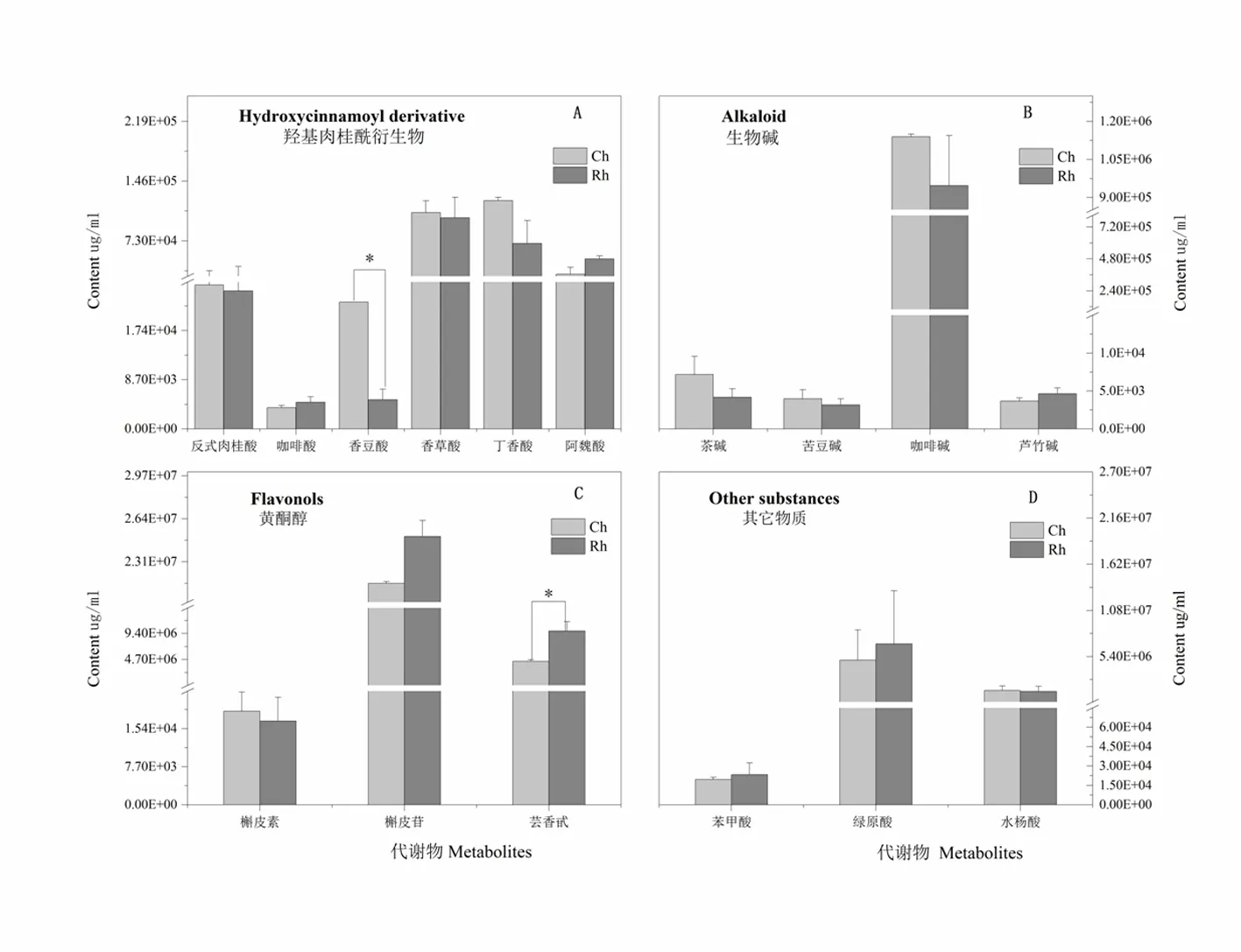
注:Ch:对照株系健康部位。Rh:抗虫株系健康部位
Note:Ch: Healthy parts of control strains. Rh: Healthy parts of insect-resistant strains
图1 新疆野苹果自然条件下次生代谢组分差异
Fig. 1 Differences of the secondary metabolites in Xinjiang wild apple under normal conditions
2.2 新疆野苹果虫害侵染条件下次生代谢组分对虫害胁迫的响应
研究表明,抗虫株系与对照株系在被小吉丁虫侵染处理后,羟基肉桂酰衍生物中的反式肉桂酸含量上升明显,达到极显著水平,含量分别为3.3×104和2.2×106μg/mL,差异倍数为6.8。同时抗虫株系的咖啡酸含量极显著高于对照株系,提升倍数为3.8。此外,阿魏酸含量也显著上升,含量分别为4.5×104和5.3×104μg/mL(图2-A)。由此可见,变化明显的大多属于羟基肉桂酰衍生物一大类,说明抗虫植株在受到虫害胁迫后,激发植物体自身调节变化,提高羟基肉桂酰衍生物次生代谢含量以抵御侵害;生物碱组分含量测定中,虫害侵染处理后,各组分均没有达到显著差异水平,其中咖啡碱含量最高,对照株系高于抗虫株系,含量为9.7×105和7.1×105μg/mL,差异倍数为1.4,苦豆碱含量最低(图2-B)。
黄酮醇类组分中,对照株系榭皮素含量显著高于抗虫株系,达到44.3%,榭皮苷与芸香苷含量在虫害侵染前后变化差异不明显,但榭皮苷含量在黄酮醇类组分中最高,分别为2.4×107和2.3×107μg/mL;在其它组分含量测定中,苯甲酸、绿原酸、水杨酸对虫害胁迫响应明显,均有不同程度的提高,其中水杨酸上升达到显著水平。图2
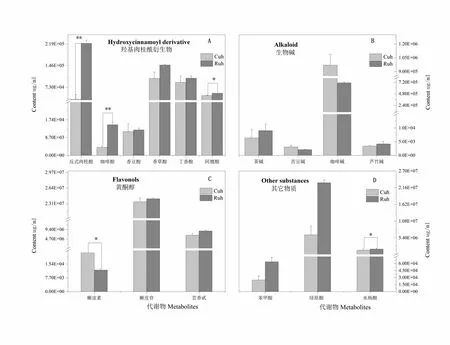
注:Cuh:对照株系侵染部位。Ruh:抗虫株系侵染部位
Note:Cuh: Infection site of control strain. Ruh: Infection site of insect-resistant strains
图2 新疆野苹果虫害侵染后次生代谢组分的响应
Fig. 2 Response of secondary metabolites after pest infestation in Xinjiang wild apple
3 讨 论
植物的次生代谢产物非常丰富,根据成分,主要分为萜类、生物碱、苯丙烷类和相关苯基化合物四大类[20],植物受到虫害侵染后,会产生诱导抗性,从而产生相关萜类化合物,组织防御体系[21],研究认为,在自然条件下,测定健康部位羟基肉桂酰衍生物、生物碱、黄酮醇及其它代谢物质变化差异不明显。在虫害胁迫条件下反式肉桂酸、咖啡酸、阿魏酸、绿原酸、水杨酸含量均显著上升,说明这些代谢物对虫害胁迫产生积极响应。此外,酚类物质、单宁、生物碱与蛋白质及酶作用,形成不易消化的复合物,影响昆虫消化率及摄食量[22]。苯丙烷类途径可生成反式肉桂酸、香豆酸、阿魏酸、芥子酸等中间产物,这些中间产物可进一步转化为香豆素、绿原酸,也可形成反式香豆酰辅酶A酯,再通过多条途径进一步转化为木质素、黄酮、异黄酮、生物碱、苯甲酸酯糖苷等次生代谢产物,这些产物对昆虫具有一定的毒害与防御作用[23]。兴安落叶松受损后其叶内单宁含量呈显著增加的趋势,北美云杉(Piceasitchensis) 和多叶羽扇豆(Lupinuspolyphyllus) 在受到伤害后,创伤树脂和生物碱的含量分别增加[24, 25]。与此同时许多酚酸对害虫威慑非常有效,是许多具有传递抗性的复合分子前体[26]。通过代谢组与转录组联合分析二化螟对水稻的响应机制认为,莽草酸、糖代谢、氨基酸、有机酸和萜类代谢等途径共同参与了水稻抗虫性调控过程[27]。这与研究结果相似,即新疆野苹果受虫害胁迫逆境条件下,通过调节自身免疫系统,造成植物“自身抗虫”的表型。
植物在受到昆虫危害后的防御反应是多维的动态过程,包含的层次与功能相对复杂[25],植物与昆虫互作机理研究通常会使用高通量组学技术(转录组学、基因组学、代谢组学等)揭示其抗性产生机理[28]。通过转录组分析并鉴定候选基因,了解植物在生物胁迫和非生物胁迫过程中的应激反应[29-31]。苹果小吉丁虫是新疆野苹果这一“起源种”的毁灭性虫害,进一步通过转录组、代谢组和生理水平研究新疆野苹果抗苹果小吉丁虫的生理生化响应机理、转录调控机制以及如何增强抗苹果小吉丁虫功能是接下来重点拟解决的关键科学问题。为有效保护、利用新疆野苹果这一珍贵资源提供理论基础。
4 结 论
新疆野苹果在被苹果小吉丁虫侵染后反式肉桂酸含量极显著提高,含量分别为3.3×104和2.2×106μg/mL,差异倍数为6.8。咖啡酸、绿原酸、槲皮素、阿魏酸组分含量显著提高,且上升明显的次生代谢物大多属羟基肉桂酰衍生物一大类,抗虫植株在受到虫害胁迫后,此类代谢物质产生积极响应,激发植物体自身调节变化,提高羟基肉桂酰衍生物等次生代谢含量。
猜你喜欢
当代水产(2022年1期)2022-04-26
中国农业科学(2021年23期)2022-01-14
江西农业学报(2021年8期)2021-09-08
昆虫学报(2021年3期)2021-05-11
林业科技(2020年3期)2021-01-21
上海师范大学学报·自然科学版(2020年5期)2020-12-18
中国瓜菜(2020年8期)2020-09-26
植物研究(2020年6期)2020-03-05
上海农业科技(2019年3期)2019-06-25
江苏农业科学(2016年10期)2017-02-05
