
稻虾共作模式氮和磷循环特征及平衡状况*
2019-08-31 02:48佀国涵袁家富彭成林夏贤格程建平徐祥玉贾平安谢媛圆周剑雄
中国生态农业学报(中英文) 2019年9期
佀国涵, 袁家富**, 彭成林, 夏贤格, 程建平, 徐祥玉, 贾平安, 谢媛圆, 周剑雄
稻虾共作模式氮和磷循环特征及平衡状况*
佀国涵1,2, 袁家富1,2**, 彭成林1,2, 夏贤格2, 程建平3, 徐祥玉1,2, 贾平安4, 谢媛圆1, 周剑雄1
(1. 湖北省农业科学院植保土肥研究所 武汉 430064; 2. 农业农村部废弃物肥料化利用重点实验室 武汉 430064; 3. 湖北省农业科学院粮食作物研究所 武汉 430064; 4. 潜江市农技推广中心 潜江 433199)
稻虾共作模式是我国长江中下游地区一种新兴的稻田复合种养生态模式, 研究稻虾共作系统中氮(N)和磷(P)的循环特征及盈亏状况, 对合理调控稻虾共作系统养分循环和平衡, 指导系统N和P优化管理具有重要的意义。本文通过田间试验, 采用投入产出法, 以中稻单作模式作为对照, 研究了稻虾共作模式下N和P的循环特征及平衡状况。结果表明: 克氏原螯虾子系统N和P的输出/输入比分别为0.62和0.44, 且子系统中N和P以饲料输入最大, 占总输入的92.9%和96.4%, 以成虾N和P输出最大, 分别占总输出的53.3%和59.5%; 在目前投入水平下, 两种模式土壤子系统中N和P表观平衡均出现盈余, 且稻虾共作模式土壤子系统N的盈余量高于中稻单作模式, 而P的盈余量则低于中稻单作模式; 稻虾共作模式和中稻单作模式的N和P输出/输入比均小于1, 且稻虾共作模式的N和P输出/输入比均小于中稻单作模式; 稻虾共作模式增加了土壤截存的N和P量, 其土壤截存的N和P量较中稻单作模式分别提高49.2 kg∙hm-2和9.1 kg∙hm-2; 稻虾共作模式提高了系统N和P表观损失量, 其系统N和P表观损失量较中稻单作模式分别提高10.2 kg∙hm-2和1.0 kg∙hm-2。可见稻虾共作模式降低了N和P输出/输入比, 促进了土壤中N和P的累积, 但增加了系统N和P表观损失量。
稻虾共作模式; 养分循环; 养分平衡; 氮磷
农业生态系统是人工生态系统, 与无限自然延续的自然生态系统相比, 农业生态系统能够持续不断地发展主要靠人为因素的补给和控制, 因此人为控制下的养分循环是建立持续可发展农业的物质基础[1-2]。生态系统中营养物质的循环是系统生产力和持久性的决定因素, 对生物圈化学环境有重大影响[3]。氮(N)和磷(P)是植物生长所必需的营养物质, 也是土壤养分的主要限制因子, 其循环特征和平衡状况对提高生态系统生产力起着重要的作用。稻虾[水稻()-克氏原螯虾()]共作模式是我国长江中下游地区一种新兴的稻田种养复合生态模式, 稻田生态系统中引入克氏原螯虾不仅能够大幅提高水、肥和能量的利用率, 增强稻田生态系统的稳定性及抗外界冲击的能力[4-5], 而且促进了系统中物质循环, 阻止了稻田物质流的外溢, 使稻田生态系统从结构与功能上得到改善和提高[6]。目前关于稻田复合种养生态系统中养分循环方面开展了大量研究, 稻鸭生态系统中碳(C)循环[7]、N循环[8-10]以及P循环[10-11]研究表明, 稻鸭生态系统C、N和P的输出/输入比均小于常规稻作模式; 稻鱼生态系统N平衡[12-14]研究表明, N的输入大于输出, 且系统中水稻吸收的31.8%的N来自于未被鱼食用的残饵; 稻蟹生态系统中农田养分平衡[15]研究表明, 稻蟹模式的农田中N和P均为盈余, 且其盈余量均高于单作稻田。目前关于稻虾共作生态系统中N和P循环特征、平衡状况等方面的研究鲜有报道, 为此本文通过设置田间试验, 采用投入产出法研究稻虾共作模式中N和P的循环特征及平衡状况, 明确农田生态系统中N和P的盈亏状况, 这为合理调控稻虾共作系统养分循环和平衡, 指导系统N和P优化管理提供了数据支撑。
1 材料与方法
1.1 试验区概况
试验区位于湖北省潜江市后湖农场移民村(30°22′41.2″N、112°41′32.5″E), 该区域属江汉平原低湖区, 冬季静态地下水位40~60 cm, 北亚热带季风湿润气候, 年均气温16.1 ℃, 无霜期246 d, 年均降雨量1 100 mm。种植制度为“中稻-冬闲”, 土壤类型为湖积物发育而成的潮土性水稻土。试验前0~20 cm耕层土壤基本理化性状为: pH 7.5, 容重0.97 g∙cm-3, 有机质 26.4 g∙kg-1, 全氮2.36 g∙kg-1, 全磷0.47 g∙kg-1, 全钾20.6 g∙kg-1, 阳离子交换量 28.1 cmol∙kg-1, 碱解氮129.5 mg∙kg-1, 速效磷7.53 mg∙kg-1, 速效钾157.7 mg∙kg-1。
1.2 试验设计
田间试验始于2014年10月, 该研究于2015年10月—2016年10月进行, 田间试验布局见图1。试验设2个处理: ①稻虾共作模式(CR): 中稻收获后灌水养虾, 稻草全量还田; ②中稻单作模式(MR): 水稻收获后冬干, 不养虾, 稻草全量还田。每个处理设置3次重复, 每小区面积100 m2, 小区周围做宽60 cm、高40 cm的埂, 并用地膜包裹。为了防止串水、串虾, 处理间设宽0.4 m、深1.0 m的沟; 在稻虾共作模式小区一侧开挖宽3.0 m、深1.0 m的虾沟, 且四周设尼龙拦虾网, 尼龙网埋入地下约1.0 m, 地上露出约0.3 m, 并用小竹棍支撑。稻草还田方式为稻草覆盖还田, 供试水稻为中稻品种‘鉴真2号’, 供试虾为克氏原螯虾。

图1 稻虾共作模式和中稻单作模式的田间试验布局图
1.3 田间管理
于每年6月中下旬进行整地、水稻移栽, 栽插的原则是宽行窄株、沟边密植, 株行距为16.7 cm×26.6 cm, 10月中下旬收割; 稻虾共作模式和中稻单作模式均施用等量等比例化肥, 施用N、P2O5、K2O分别为180 kg∙hm-2、90 kg∙hm-2、144 kg∙hm-2, 氮肥施用比例为基肥∶分蘖肥∶穗肥=5.8∶1.8∶2.4, 磷肥一次性基肥施入, 钾肥施用比例为基肥∶穗肥=5∶5。插秧前施入基肥, 插秧后15 d施入分蘖肥, 插秧后50 d施入穗肥。每公顷补充武汉高飞农业有限公司生产的大粒锌(Zn≥25%)6 kg和大粒硅(SiO2≥20%)60 kg。
在稻虾共作模式中, 虾苗于2014年10月按照重量为(5±2) g的幼虾9.0万只∙hm-2的标准进行投放, 虾苗在稻田中自行繁殖, 以后每年此时根据实际情况补投适量亲虾, 在每年的3—5月进行虾饲料投放, 投入饲料量平均为1 800 kg∙hm-2, 于每年6月上旬捕捞成虾完毕, 而未成熟的幼虾随水迁移至虾沟中, 待整田、插秧、晒田控蘖及复水后再次进入稻田生活, 在中稻收获前收获第2季成熟虾, 10月底中稻收割完毕灌水后进行下一季克氏原螯虾养殖, 如此循环轮替(图2)。稻田灌溉水来自附近的沟渠, 水稻季各处理田面水均维持在10 cm, 当田面水降至5 cm时进行稻田灌水, 水稻齐穗期以后稻田停止灌溉; 克氏原螯虾养殖季稻田水面维持在50 cm, 并于水稻种植前将田间养殖废水排干后进行整地、施肥及插秧等农事操作。稻虾共作模式和中稻单作模式农田系统边界及循环模式见图3。
1.4 样品的采集与测定方法
1.4.1 土壤样品测定
在2015年10月15日和2016年10月17日, 依据S形5点采样法于每个小区采集0~20 cm土层土样, 采集土样去除植物残根和石块并混匀, 风干过筛后备用; 采用环刀法测定耕层土壤容重。土壤全氮采用半微量开氏法测定, 全磷采用浓硫酸-高氯酸消煮钼锑抗比色法测定[16]。
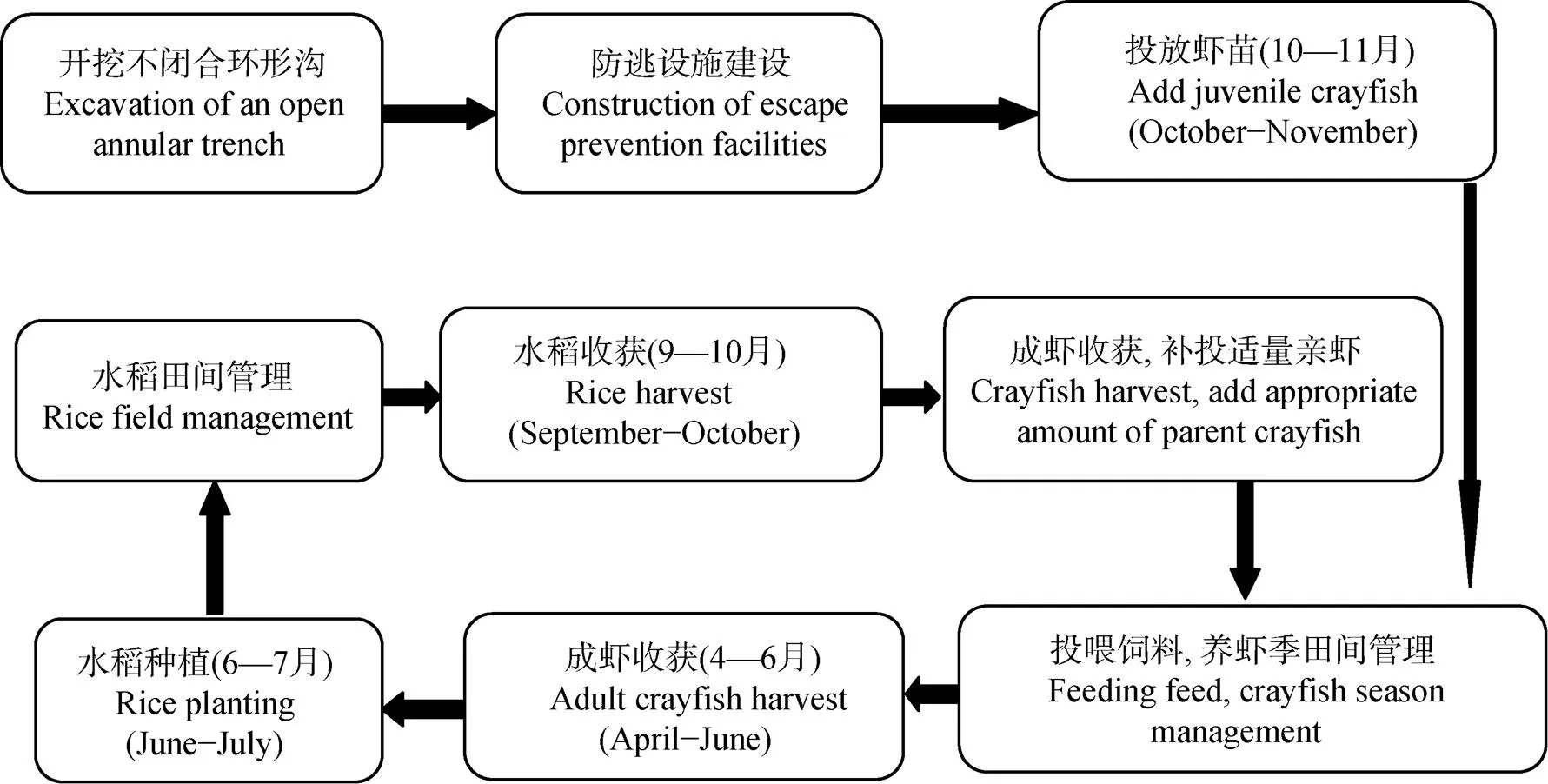
图2 稻虾共作模式示意图
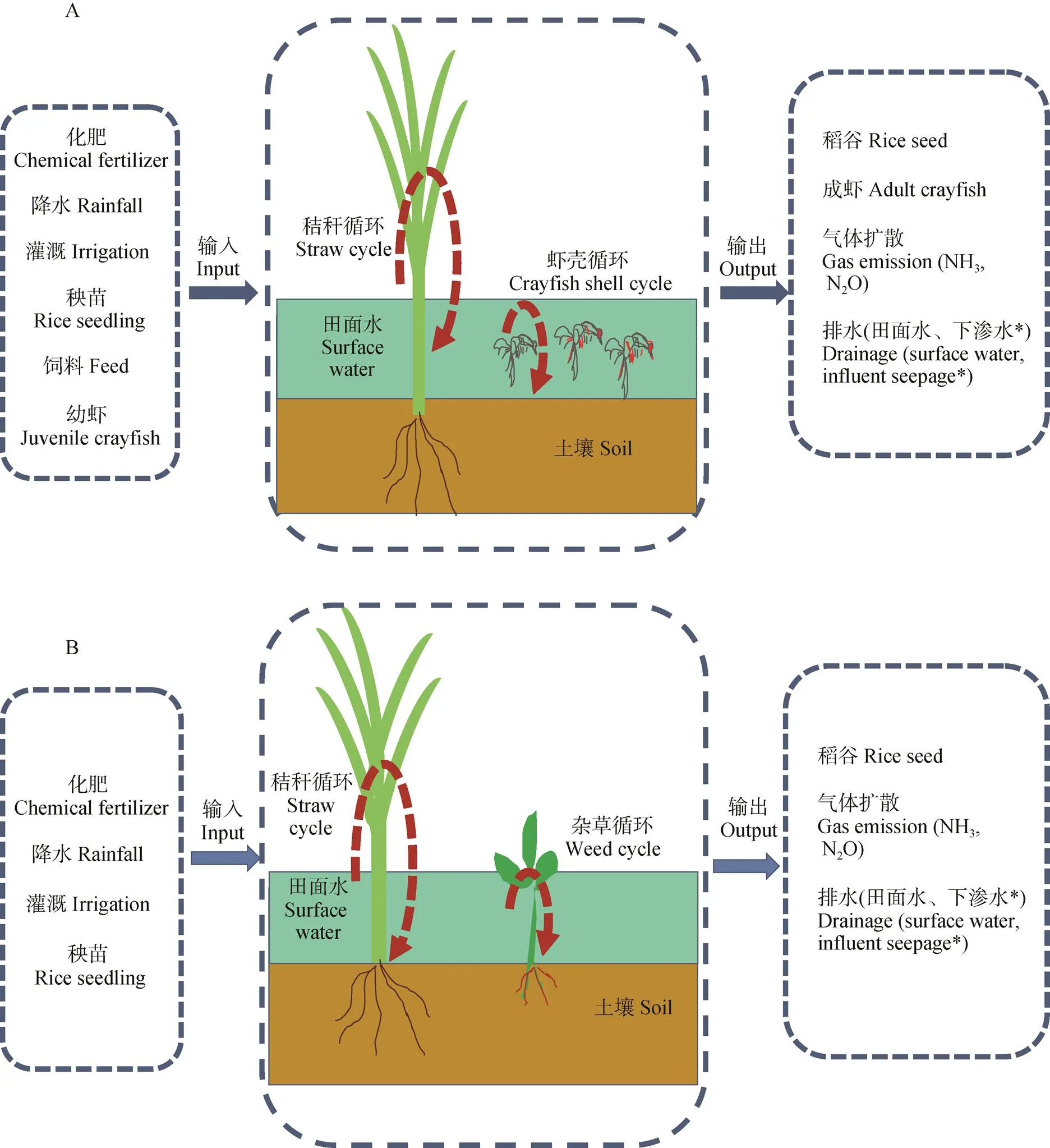
图3 稻虾共作模式(A)和中稻单作模式(B)农田系统边界及循环模式
1.4.2 水稻植物样品测定
在水稻苗期, 每小区随即取5个1 m2样方秧苗测定植株的生物量; 在水稻收获期, 每小区随机取5个1 m2样方测定水稻籽粒和秸秆生物量, 根系生物量按照根冠比0.1计算[17]。将烘干植物样品磨碎过筛后, 经浓硫酸-过氧化氢高温消化, 半微量蒸馏法测全氮含量, 钼锑抗比色法测定其全磷含量[16]。
1.4.3 杂草植物样品测定
相对于冬季泡水养虾的稻虾共作模式, 中稻单作模式为冬闲, 稻田中会生长大量杂草, 其种类主要为菵草()、棒头草()和丁香蓼()。在中稻单作模式下于2016年5月底稻田整地、施肥前, 每小区随机取3个1 m2样方测定杂草生物量, 同时进行杂草取样, 测定全氮和全磷含量, 测定方法同水稻植物样品分析方法。
1.4.4 克氏原螯虾饲料及克氏原螯虾样品测定
每天详细记录成虾出售重量, 计算整年度成虾产出总量, 详细记载幼虾投入量。于2016年4月1日、5月10日和5月25日分别取幼虾、饲料和成虾进行含水量、全氮和全磷含量分析, 全氮和全磷的测定方法同水稻植物样品分析方法。
1.4.5 虾壳样品测定
幼虾(约5 g)至性成熟一般需要经过5~6次的蜕壳[18]。在克氏原螯虾养殖季2016年3月20日投入至5月20日, 每隔10 d对模拟养殖箱中克氏原螯虾进行称重, 共计6次, 按照测定的虾壳占虾重平均比值12.4%计算, 根据输出成虾的总重量, 估算出克氏原螯虾蜕壳总量; 另取完整虾壳进行全氮和全磷含量分析, 其测定方法同水稻植物样品分析方法。
1.4.6 灌溉水样品测定
记载水稻整个生育期间和养虾季灌水量以及养殖结束时养殖废水的排放量, 从稻田进水口处于2015年10月—2016年10月每10 d采集1次水样, 同时在养殖废水排放前采集水样; 采集水样的全磷含量采用过硫酸钾氧化-钼蓝比色法测定, 全氮含量采用过硫酸钾氧化-紫外分光光度法测定[19]。
1.4.7 降雨样品测定
于2015年10月—2016年10月利用SDM6型雨量器, 连续不间断地收集雨水样品; 每次降雨结束后, 于次日9:00左右先记录降雨量, 然后采集混合均匀的雨水样品, 冷冻保存分析备用, 同时将储水瓶洗净放回圆筒内。降雨中全氮和全磷的测定方法同灌溉水样分析。氮和磷湿沉降量用实际收集的降水的N、P浓度与相应降雨量的乘积累加求得[20]。
1.4.8 N2O采集测定及计算
N2O气体采用静态箱采集, Aglient 7980A气相色谱仪分析测定[21]。在水稻秧苗移栽后10 d开始直至水稻收获, 每7~10 d采样1次, 在非水稻季10~15 d采集1次, 每个处理设置3次重复, 采样时间为每日上午8:00—10:00。采气时预先在底座水槽中注入水, 然后盖上采气箱, 采样箱大小为42 cm×42 cm×50 cm, 在盖箱后0 min、5 min、10 min、15 min、20 min用30 mL医用注射器采集1个气体样品, 24 h内带回实验室进行分析测定完毕。通过插值法进行累积排放量计算, 即以相邻两次监测日的平均通量乘以此期间天数作为该段时间累计排放量, 以此类推, 然后将各个时间段的累积排放量进行相加, 作为整个监测期累积排放量。
N2O气体通量计算公式为:
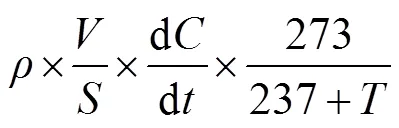
式中:为气体通量,为标准状态下N2O的密度(kg∙m-3),为密闭箱有效体积(m3),为底座面积(m2), d/d表示单位时间内密闭箱内N2O的浓度变化量,为密闭箱内平均气温。
1.4.9 NH3挥发测定及计算
稻田土壤NH3挥发采用田间原位通气法测定[22]。分别于基肥、分蘖肥和穗肥施用后的第1周每天采样1次, 第2周每2天采样1次, 之后根据测到的挥发氨量, 每7 d取样1次, 直至监测不到NH3挥发。每个小区均匀放置2个收集装置, 收集的海绵带回实验室, 采用300 mL 1.0 mol∙L-1的KCl溶液进行浸提, 在25 ℃恒温转速180 r∙min-1条件下震荡1 h后过滤, 采用AA3连续流动分析仪测定滤液中的铵态氮含量。土壤NH3挥发速率和累积量计算方法参考文献[22]。
1.4.10 数据统计分析
测定数据利用Excel 2007进行初处理, 采用SPSS 21.0软件进行统计方差分析, 最小显著性检验(LSD)法比较不同处理间的差异显著性, 以不同小写字母表示同年度不同模式间差异达<0.05显著水平。
2 结果与分析
2.1 克氏原螯虾子系统N、P循环与平衡状况
稻虾共作模式由于克氏原螯虾的引入使得整个系统具有次级生产, 克氏原螯虾觅食、蜕壳等活动使系统更加复杂, 系统的N和P投入与产出也随之发生改变。由表1可知, 克氏原螯虾子系统N和P输入均大于输出; 克氏原螯虾系统输入饲料为1 800.0 kg∙hm-2, 饲料N和P含量分别为46.6 g∙kg-1和11.0 g∙kg-1, 按照5.8%含水量计算, 饲料带入的N和P分别为79.0 kg∙hm-2和18.6 kg∙hm-2, 分别占总输入的92.9%和96.4%。稻虾共作模式中投入的幼虾经过6次蜕壳后, 累积的虾壳重量达201.3 kg∙hm-2, 蜕壳中N和P的含量分别为51.8 g∙kg-1和8.9 g∙kg-1, 可为克氏原螯虾子系统输出N和P分别为10.4 kg∙hm-2和1.8 kg∙hm-2; 成虾产量为2 250 kg∙hm-2, 其N和P含量分别为56.2 g∙kg-1和10.0 g∙kg-1, 按照77.9%含水量计算, 成虾输出的N和P分别为27.9 kg∙hm-2和5.0 kg∙hm-2, 分别占总输出的53.3%和59.5%。Kristiansen等[23]研究表明在20 ℃且喂养饲料的情况下, 成虾每年通过排泄物输出的N和P量分别为24.9 g∙kg-1和2.8 g∙kg-1, 克氏原螯虾体重的增长呈典型S型曲线, 幼虾在条件适宜下90 d即可达到上市标准[24], 据此粗略估算克氏原螯虾排泄物输出的N和P量分别为14.0 kg∙hm-2和1.6 kg∙hm-2。克氏原螯虾子系统中N和P的输出/输入比值分别为0.62和0.44, 这表明在克氏原螯虾子系统中部分饲料N和P滞留在稻虾共作系统中。
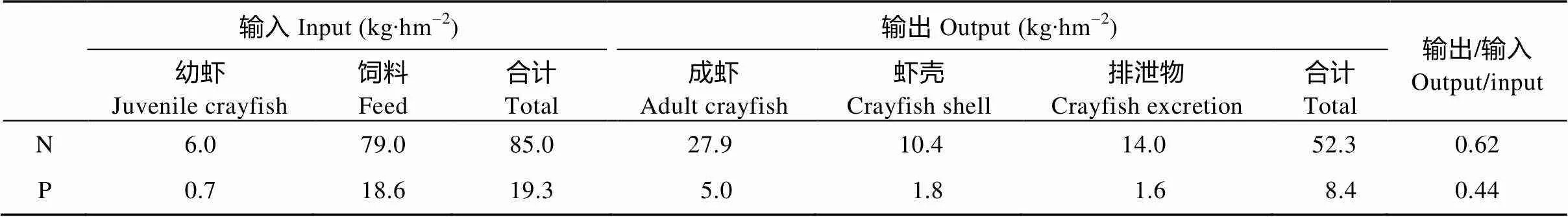
表1 稻虾共作模式中克氏原螯虾子系统N和P循环与平衡状况
2.2 土壤子系统N和P循环与平衡状况
稻虾共作模式和中稻单作模式土壤子系统N和P循环见表2。稻虾共作模式的土壤子系统N和P的输入均高于中稻单作模式。由不同稻作模式下水稻植株N、P吸收量可知(表3), 稻虾共作模式水稻籽粒、秸秆和根系吸收的N和P较中稻单作模式呈增加趋势, 其中稻虾共作模式的水稻根系吸收的P显著高于中稻单作模式(<0.05), 较中稻单作模式提高33.3%。中稻单作模式下冬闲季节杂草的生物量为1 388.0 kg∙hm-2, 其输出的N和P分别为9.8 kg∙hm-2和2.7 kg∙hm-2。
在水稻种植季稻田长期保持10 cm水层, 两种模式的灌溉水体积均为12 000 m3∙hm-2, 灌溉水N和P平均浓度分别为1.9 mg∙L-1和0.2 mg∙L-1, 故水稻生长季灌溉水N和P的输入分别为23.3 kg∙hm-2和2.8 kg∙hm-2; 克氏原螯虾养殖季稻田长期维持50 cm水层, 克氏原螯虾养殖季灌水和养殖季结束排出的养殖废水体积分别为9 000 m3∙hm-2和5 000 m3∙hm-2, 根据克氏原螯虾养殖季灌溉水N和P平均浓度分别为1.97 mg∙L-1和0.25 mg∙L-1, 养殖废水中N和P平均浓度分别为2.21 mg∙L-1和0.50 mg∙L-1计算, 克氏原螯虾养殖季灌溉水输入的N和P分别为6.7 kg∙hm-2和-0.3 kg∙hm-2, 因此稻虾共作模式下在水稻种植季和克氏原螯虾养殖季由灌水输入的N和P总量分别为30.0 kg∙hm-2和2.5 kg∙hm-2。稻虾共作模式和中稻单作模式在整个水稻生育期间由N2O排放的气态N输出分别为0.9 kg∙hm-2和0.7 kg∙hm-2, 而在非水稻季由N2O排放输出的气态N分别为0.4 kg∙hm-2和0.6 kg∙hm-2, 因此在整年监测周期间, 稻虾共作模式和中稻单作模式由N2O排放输出的气态N均为1.3 kg∙hm-2; 稻虾共作模式和中稻单作模式由NH3挥发输出的气态N分别为10.7 kg∙hm-2和11.7 kg∙hm-2。可见在不考虑水稻秸秆、根系及虾壳等归还的情况下, 两种模式土壤子系统中N和P表观平衡均出现盈余, 且稻虾共作模式土壤子系统的N盈余量高于中稻单作模式, 而P盈余量则低于中稻单作模式。
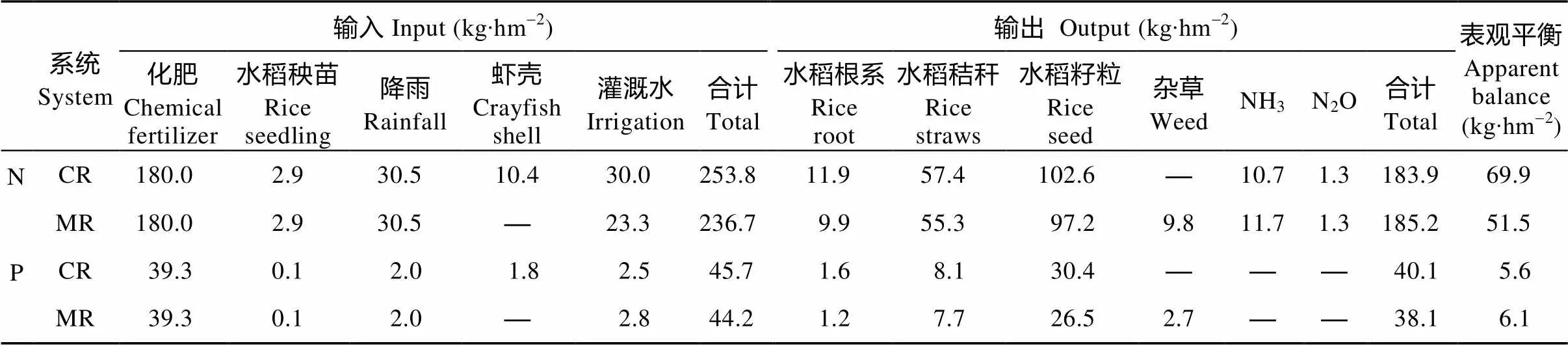
表2 稻虾共作模式和中稻单作模式的土壤子系统N和P循环
CR: 稻虾共作模式; MR: 中稻单作模式。CR: integrated rice-crayfish system; MR: rice monoculture system.
2.3 稻虾共作模式N循环特征及平衡状况
由表4可知, 中稻单作模式中N输入主要是化学肥料、灌溉水及中稻秧苗带入, 其中化肥N占总输入的76.0%。中稻单作模式只有初级生产, 所以N输出主要是收获的水稻籽粒, 输出N为97.2 kg∙hm-2; 水稻栽培系统循环的N主要是水稻秸秆N、水稻根系N和杂草N, 其分别占循环总量的73.7%、13.2%和13.1%。稻虾共作模式因有次级生产——克氏原螯虾的引入, N的输入、输出以及循环量均高于中稻单作模式。稻虾共作模式N输入, 化肥N占54.8%, 饲料N占24.1%; 稻虾共作模式N输出为142.5 kg∙hm-2, 其中水稻籽粒N占72.0%, 而成虾N占19.6%; 系统循环的N主要为水稻秸秆N、水稻根系N和虾壳N, 其分别占循环总量的72.0%、14.9%和13.1%。两种模式的N输出/输入均小于1, 且稻虾共作模式N输出/输入比均低于中稻单作模式, 这主要与系统中加入次级生产后, N输入量大幅提高有关。稻虾共作模式和中稻单作模式的土壤截存的N分别为166.7 kg∙hm-2和117.5 kg∙hm-2, 根据物质循环与平衡的原理推算出系统N表观损失量(N输入-N输出-土壤N截留量), 稻虾共作模式和中稻单作模式的N表观损失量分别为19.2 kg∙hm-2和9.0 kg∙hm-2, 可见稻虾共作模式有利于土壤中N素累积, 但增加了系统N表观损失量。
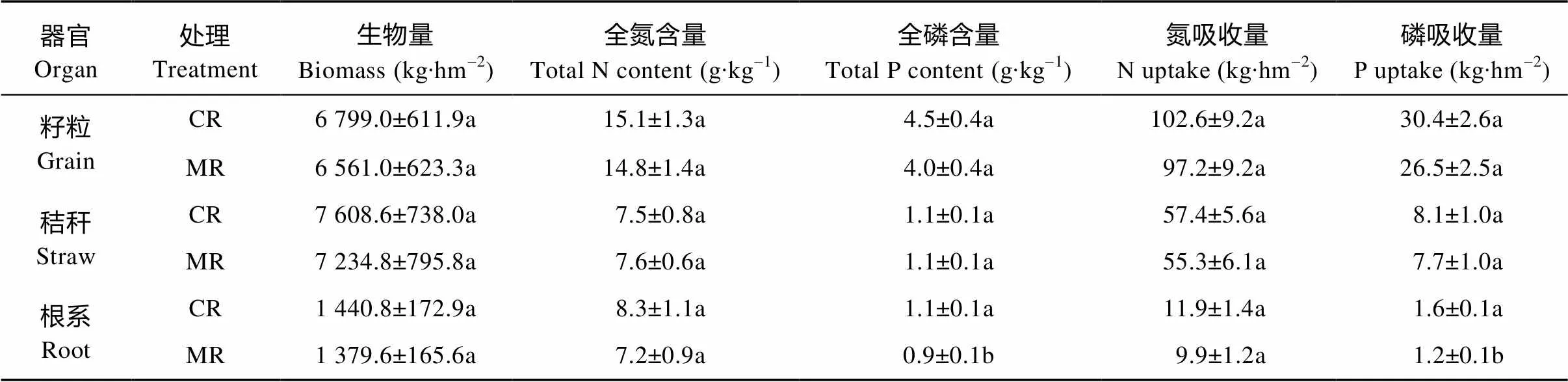
表3 稻虾共作模式和中稻单作模式中水稻植株的氮、磷吸收量
CR: 稻虾共作模式; MR: 中稻单作模式。同列数据同一器官不同字母表示两种模式间差异达5%显著水平。CR: integrated rice-crayfish system; MR: rice monoculture system. Values of the same organ in a column followed by different letters are significantly different at 5% level.
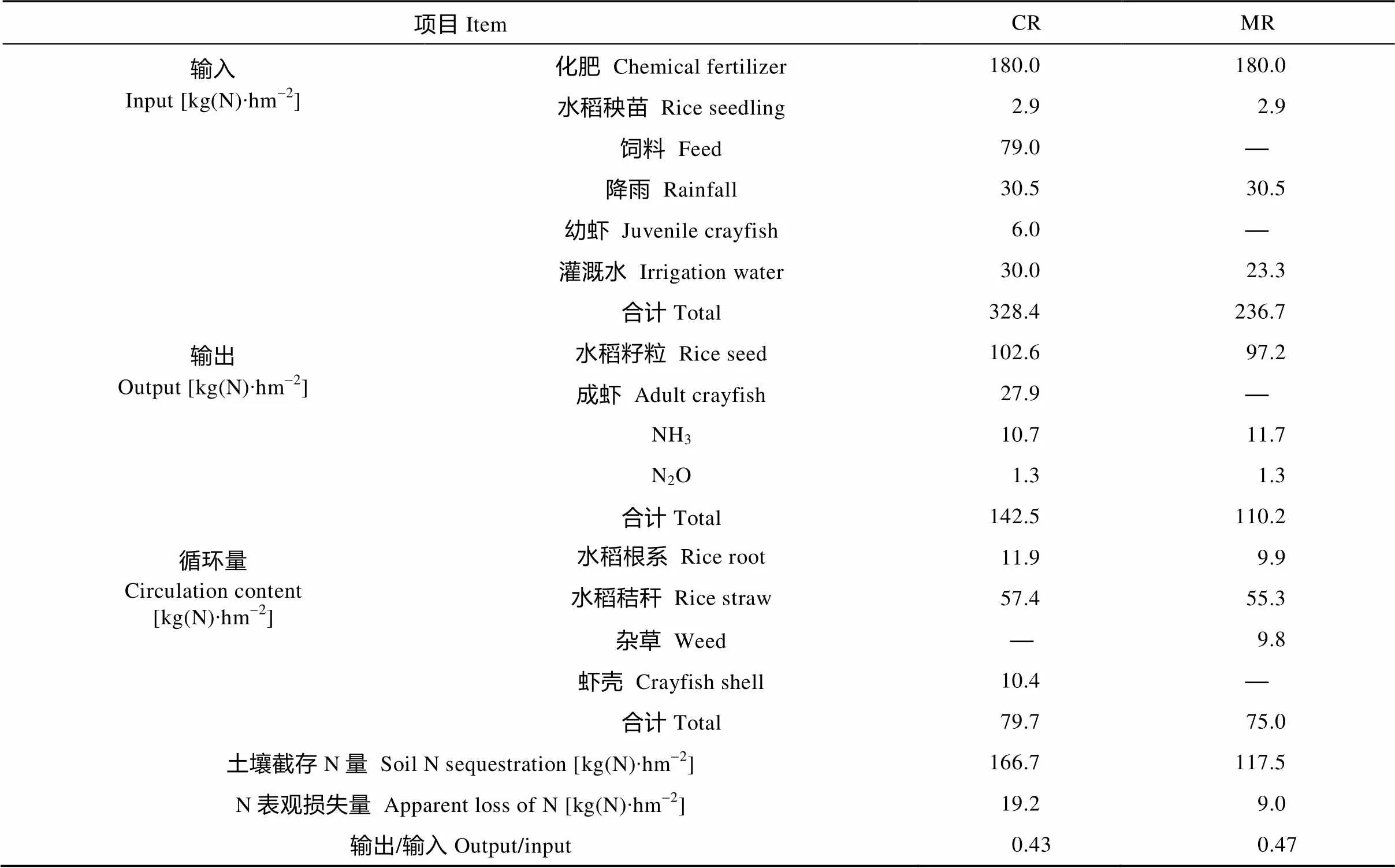
表4 稻虾共作模式和中稻单作模式的N循环与平衡状况
CR: 稻虾共作模式; MR: 中稻单作模式。CR: integrated rice-crayfish system; MR: rice monoculture system.
2.4 稻虾共作模式P循环特征及平衡状况
由表5可知, 中稻单作模式中P输入主要是化学肥料、灌溉水及中稻秧苗带入, 其中化肥P占总输入的88.9%。中稻单作模式中只有初级生产, 所以P输出主要是收获的水稻籽粒, 其P输出量为26.5 kg∙hm-2; 系统循环的P主要是水稻秸秆P、水稻根系P和杂草P, 其分别占循环量的66.4%、10.3%和23.3%。稻虾共作模式因有次级生产——克氏原螯虾的引入, P的输入和输出均高于中稻单作模式, 稻虾共作模式P输入中, 化肥P占62.2%, 饲料P占29.4%; 稻虾共作模式P输出为35.4 kg∙hm-2, 其中水稻籽粒P占85.9%, 而成虾P占14.1%; 系统循环的P为水稻秸秆P、水稻根系P和虾壳P, 其分别占总循环量的70.4%、13.9%和15.7%。两种模式的P输出/输入比均小于1, 且稻虾共作模式P输出/输入比均低于中稻单作模式, 这表明稻虾共作模式中P盈余量大于中稻单作模式。由表5可知, 稻虾共作模式和中稻单作模式土壤截存的P分别为26.2 kg∙hm-2和17.1 kg∙hm-2, 根据物质循环与平衡的原理推算出系统P表观损失量(P输入-P输出-土壤P截留量), 稻虾共作模式和中稻单作模式的P表观损失量分别为1.6 kg∙hm-2和0.6 kg∙hm-2, 可见稻虾共作模式有利于土壤中P的累积, 但增加了系统P表观损失量。

表5 稻虾共作模式和中稻单作模式的P循环与平衡状况
CR: 稻虾共作模式; MR: 中稻单作模式。CR: integrated rice-crayfish system; MR: rice monoculture system.
3 讨论
稻虾共作模式是运用生态位、食物链以及种群间互利共生等理论构建出的一种立体高效、绿色安全、生态环保的复合种养农业新模式, 已经成为我国长江中下游稻作区重要的生态农业模式[25]。稻虾之间存在着明显的互利共生关系, 克氏原螯虾蜕壳及残饵等为水稻生长提供了丰富的营养, 另外克氏原螯虾掘穴等生物扰动改善了稻田土壤的通透性, 协同了土壤水、肥、气、热, 从而有利于水稻的生长[26-27]。本研究表明, 稻虾共作模式水稻籽粒、秸秆和根系吸收的N和P较中稻单作模式呈增加趋势, 其中稻虾共作模式的水稻根系吸收的P显著高于中稻单作模式(<0.05)。土壤子系统N和P的盈亏状况不仅关系到土壤肥力的高低, 而且影响着系统的养分平衡。在目前的投入水平下, 两种模式土壤子系统中N和P均出现盈余, 且稻虾共作模式土壤子系统的N盈余量高于中稻单作模式, 而P的盈余量则低于中稻单作模式。这主要是由于中稻单作模式冬闲季节稻田生长的杂草, 增加了其土壤子系统中N和P的输出, 但是由于稻虾共作模式中水稻根系P的输出量显著高于中稻单作模式, 从而使得其盈余量低于中稻单作模式, 因此稻虾共作模式中应增加磷肥投入, 增加土壤中磷和氮交互作用, 从而提高水稻的产量。
克氏原螯虾为杂食性动物, 主要摄食绿色幼嫩植物、动植物碎屑以及浮游生物等[28]。在稻虾共作模式中为了提高克氏原螯虾的品质和产量, 往往投入一定量的饲料从而达到短时间内实现经济效益的目的。李聪等[29]研究表明, 克氏原螯虾的食物来源中饲料贡献率为38.0%~44.2%, 而浮游动物和有机碎屑的贡献率分别为11.2%~14.4%和21.2%~23.4%, 其他的食物贡献率主要来自于幼嫩植物、藻类等。本研究表明克氏原螯虾子系统输入的N和P主要以饲料带入, 且其N和P的输出/输入比分别为0.62和0.44, 这表明在克氏原螯虾子系统中一部分饲料的N和P滞留在系统中。克氏原螯虾的排泄量主要与温度、食物以及体重等有关, 且其排泄物主要以水溶性排泄物为主[30], 因此在土壤子系统中未考虑其输入量。
稻虾共作模式中克氏原螯虾通过摄食系统中的浮游动物、有机碎屑以及幼嫩植物等, 将中稻单作模式中流失的物质和能量截留利用起来, 从而提高了系统输出量。本研究表明, 稻虾共作模式N和P的输出均高于中稻单作模式, 其中成虾输出的N和P分别占总输出的19.6%和14.1%。本研究表明, 相对于中稻单作模式, 稻虾共作模式在水稻季增加了N2O排放, 但在非水稻季则降低了N2O排放, 这与孙自川[31]的研究一致。稻虾共作模式下由于克氏原螯虾的掘穴活动, 提高了土壤的通透性, 在水稻季晒田等农事操作提高了土壤的硝化作用, 促进了N2O排放; 而在非稻季, 由于稻田长期淹水养虾, 土壤处于强还原状态, 硝化作用受到抑制, 从而降低了N2O排放。本研究表明, 稻虾共作模式的NH3挥发累积量低于中稻单作模式, 且两种模式的NH3挥发累积量占施氮量的比例为6.0%~6.5%, 这与李诗豪等[32]报道的5.4%~13.0%的研究结论较为接近; 另外在稻虾共作模式下, 由于克氏原螯虾的扰动, 改善了土壤结构, 促进了水稻对N的吸收, 在一定程度上降低了稻田的NH3挥发损失量。
由于养分在农田生态系统中存在各种损失, 因此养分输入与支出间的平衡状况是表征农田养分管理是否可持续的重要指标, 可反映土壤养分水平的变化状况[33]。本研究表明, 稻虾共作模式中N和P的输出/输入比均低于中稻单作模式, 且均小于1, 稻虾共作模式土壤截存的N和P量较中稻单作模式分别提高49.2 kg∙hm-2和9.1 kg∙hm-2, 这表明稻虾共作模式N和P盈余量均高于中稻单作模式, 而且盈余的N和P进入了土壤黑箱中, 促进了稻虾共作模式土壤中N和P的累积。
4 结论
稻虾共作模式中克氏原螯虾子系统N和P主要以饲料输入和成虾输出比例最大, 其中N和P的输出/输入比值分别为0.62和0.44; 当前投入水平下, 两种模式下土壤子系统中N和P表观平衡均出现盈余, 且稻虾共作模式土壤子系统的N盈余量高于中稻单作模式, 而P盈余量则低于中稻单作模式; 稻虾共作模式和中稻单作模式中N和P输出/输入比值均小于1, 且稻虾共作模式的N和P输出/输入比小于中稻单作模式, 且其土壤截存的N和P量较中稻单作模式分别提高了49.2 kg∙hm-2和9.1 kg∙hm-2, 促进了土壤中N和P的累积; 稻虾共作模式的N和P表观损失量较中稻单作模式分别提高了10.2 kg∙hm-2和1.0 kg∙hm-2, 增加了系统N和P表观损失量。
[1] 鲁如坤, 刘鸿翔, 闻大中, 等. 我国典型地区农业生态系统养分循环和平衡研究Ⅰ. 农田养分支出参数[J]. 土壤通报, 1996, 27(4): 145–150 LU R K, LIU H X, WEN D Z, et al. Study on nutrient cycling and balance of China’s typical agri-ecosystem Ⅰ. Output parameter of farmland nutrient[J]. Chinese Journal of Soil Science, 1996, 27(4): 145–150
[2] 时正元, 鲁如坤. 农田养分再循环研究Ⅰ. 作物秸秆养分的利用率[J]. 土壤, 1993, 25(6): 281–285 SHI Z Y, LU R K. Study on nutrient recycling of farmland. Ⅰ. Nutrient use efficiency of crop straw[J]. Soils, 1993, 25(6): 281–285
[3] 曹凑贵, 张光远, 王运华. 农业生态系统养分循环研究概况[J]. 生态学杂志, 1998, 17(4): 26–32 CAO C G, ZHANG G Y, WANG Y H. Situation of research on nutrient cycling in agroecosystem[J]. Chinese Journal of Ecology, 1998, 17(4): 26–32
[4] 程慧俊. 克氏原螯虾稻田养殖生态学的初步研究[D]. 武汉: 湖北大学, 2014: 7–10 CHENG H J. A preliminary study on the ecology aspects culture of the crayfish () in rice fields[D]. Wuhan: Hubei University, 2014: 7–10
[5] 曹凑贵, 江洋, 汪金平, 等. 稻虾共作模式的“双刃性”及可持续发展策略[J]. 中国生态农业学报, 2017, 25(9): 1245–1253 CAO C G, JIANG Y, WANG J P, et al. “Dual character” of rice-crayfish culture and strategies for its sustainable development[J]. Chinese Journal of Eco-Agriculture, 2017, 25(9): 1245–1253
[6] 奚业文, 周洵. 稻虾连作共作稻田生态系统中物质循环和效益初步研究[J]. 中国水产, 2016, (3): 78–82 XI Y W, ZHOU X. A preliminary study on the material cycle and benefit of rice-crayfish continuous cropping and co-cropping system in paddy fields[J]. China Fisheries, 2016, (3): 78–82
[7] 张帆, 高旺盛, 隋鹏, 等. “双季稻-鸭”共生生态系统C循环[J]. 生态学报, 2012, 32(10): 3198–3208 ZHANG F, GAO W S, SUI P, et al. Carbon cycling from rice-duck mutual ecosystem during double cropping rice growth season[J]. Acta Ecologica Sinica, 2012, 32(10): 3198–3208
[8] 张帆, 陈源泉, 隋鹏, 等. “双季稻-鸭”共生生态系统稻作季节氮循环[J]. 应用生态学报, 2012, 23(1): 178–184 ZHANG F, CHEN Y Q, SUI P, et al. Nitrogen cycling in rice-duck mutual ecosystem during double cropping rice growth season[J]. Chinese Journal of Applied Ecology, 2012, 23(1): 178–184
[9] LI C F, CAO C G, WANG J P, et al. Nitrogen losses from integrated rice-duck and rice-fish ecosystems in southern China[J]. Plant and Soil, 2008, 307(1/2): 207–217
[10] 张帆, 隋鹏, 陈源泉, 等. “稻鸭共生”生态系统稻季N、P循环[J]. 生态学报, 2011, 31(4): 1093–1100 ZHANG F, SUI P, CHEN Y Q, et al. Nitrogen and phosphorus cycling from rice-duck mutual ecosystem during late rice growth season[J]. Acta Ecologica Sinica, 2011, 31(4): 1093–1100
[11] 张帆, 陈源泉, 高旺盛. “双季稻-鸭”共生生态系统稻季磷循环[J]. 生态学杂志, 2012, 31(6): 1383–1389 ZHANG F, CHEN Y Q, GAO W S. Phosphorus cycling in rice-duck mutual ecosystem in double cropping rice growth seasons[J]. Chinese Journal of Ecology, 2012, 31(6): 1383–1389
[12] 谢坚. 农田物种间相互作用的生态系统功能——以全球重要农业文化遗产“稻鱼系统”为研究范例[D]. 杭州: 浙江大学, 2011: 13–19 XIE J. Ecosystem functioning of species interactions in farming system: A case study on globally important agricultural heritage system[D]. Hangzhou: Zhejiang University, 2011: 13–19
[13] 吴雪. 稻鱼系统养分循环利用研究[D]. 杭州: 浙江大学, 2012: 17–33WU X. The utilization of nutrients in traditional rice-fish co-culture system[D]. Hangzhou: Zhejiang University, 2012: 17–33
[14] OEHME M, FREI M, RAZZAK M A, et al. Studies on nitrogen cycling under different nitrogen inputs in integrated rice-fish culture in Bangladesh[J]. Nutrient Cycling in Agroecosystems, 2007, 79(2): 181–191
[15] 陈飞星, 张增杰. 稻田养蟹模式的生态经济分析[J]. 应用生态学报, 2002, 13(3): 323–326 CHEN F X, ZHANG Z J. Ecological economic analysis of a rice-crab model[J]. Chinese Journal of Applied Ecology, 2002, 13(3): 323–326
[16] 鲍士旦. 土壤农化分析[M]. 3版. 北京: 中国农业大学出版社, 2000: 257–282 BAO S D. Soil and Agricultural Chemistry Analysis[M]. 3rd ed. Beijing: China Agriculture Press, 2000: 257–282
[17] 展茗. 不同稻作模式稻田碳固定、碳排放和土壤有机碳变化机制研究[D]. 武汉: 华中农业大学, 2009: 41–43 ZHAN M. Studies on mechanisms of carbon sequestration, carbon emissions and soil organic carbon dynamics in different farming paddy fields[D]. Wuhan: Huazhong Agricultural University, 2009: 41–43
[18] CASTAÑON-CERVANTES O, LUGO C, AGUILAR M, et al. Photoperiodic induction on the growth rate and gonads maturation in the crayfishduring ontogeny[J]. Comparative Biochemistry and Physiology Part A: Physiology, 1995, 110(2): 139–146
[19] 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2000: 12–226 LU R K. Analytical Method of Soil and Agricultural Chemistry[M]. Beijing: China Agricultural Science and Technology Press, 2000: 12–226
[20] 刘冬碧, 张小勇, 巴瑞先, 等. 鄂西北丹江口库区大气氮沉降[J]. 生态学报, 2015, 35(10): 3419–3427 LIU D B, ZHANG X Y, BA R X, et al. Atmospheric nitrogen deposition in Danjiangkou Reservoir area of Northwest Hubei[J]. Acta Ecologica Sinica, 2015, 35(10): 3419–3427
[21] 徐祥玉, 张敏敏, 彭成林, 等. 稻虾共作对秸秆还田后稻田温室气体排放的影响[J]. 中国生态农业学报, 2017, 25(11): 1591–1603 XU X Y, ZHANG M M, PENG C L, et al. Effect of rice-crayfish co-culture on greenhouse gases emission in straw-puddled paddy fields[J]. Chinese Journal of Eco-Agriculture, 2017, 25(11): 1591–1603
[22] LI P F, LU J W, WANG Y, et al. Nitrogen losses, use efficiency, and productivity of early rice under controlled-release urea[J]. Agriculture, Ecosystems & Environment, 2018, 251: 78–87
[23] KRISTIANSEN G, HESSEN D O. Nitrogen and phosphorus excretion from the noble crayfish,L., in relation to food type and temperature[J]. Aquaculture, 1992, 102(3): 245–264
[24] 韩光明, 张家宏, 王守红, 等. 克氏原螯虾生长规律及大规格生态养殖的关键技术和效益分析[J]. 江西农业学报, 2015, 27(2): 91–94 HAN G M, ZHANG J H, WANG S H, et al. Growth regularity of, key technology and benefit analysis of its large-size ecological rearing[J]. Acta Agriculturae Jiangxi, 2015, 27(2): 91–94
[25] SI G H, YUAN J F, XU X Y, et al. Effects of an integrated rice-crayfish farming system on soil organic carbon, enzyme activity, and microbial diversity in waterlogged paddy soil[J]. Acta Ecologica Sinica, 2018, 38(1): 29–35
[26] SI G H, PENG C L, YUAN J F, et al. Changes in soil microbial community composition and organic carbon fractions in an integrated rice-crayfish farming system in subtropical China[J]. Scientific Reports, 2017, 7(1): 2856
[27] 佀国涵, 彭成林, 徐祥玉, 等. 稻虾共作模式对涝渍稻田土壤理化性状的影响[J]. 中国生态农业学报, 2017, 25(1): 61–68 SI G H, PENG C L, XU X Y, et al. Effect of integrated rice-crayfish farming system on soil physico-chemical properties in waterlogged paddy soils[J]. Chinese Journal of Eco-Agriculture, 2017, 25(1): 61–68
[28] COVICH A P. How do crayfish respond to plants and mollusca as alternate food resources[J]. Freshwater Crayfish, 1977, 3: 165–179
[29] 李聪, 成永旭, 管勤壮, 等. 用稳定性同位素技术分析稻-虾系统中不同“碳/氮”投喂方式对克氏原螯虾食性的影响[J]. 水产学报, 2018, 42(11): 1778–1786 LI C, CHENG Y X, GUAN Q Z, et al. Using stable isotopes to estimate the effects of different carbon/nitrogen feeds on feeding habit ofin the rice-crayfish co-culture system[J]. Journal of Fisheries of China, 2018, 42(11): 1778–1786
[30] 温小波, 库夭梅, 罗静波. 温度、体重及摄食状态对克氏原螯虾代谢的影响[J]. 华中农业大学学报, 2003, 22(2): 152–156 WEN X B, KU Y M, LUO J B. Effects of temperature, body weight and feeding on metabolism of[J]. Journal of Huazhong Agricultural University, 2003, 22(2): 152–156
[31] 孙自川. 稻虾共作下秸秆还田和投食对温室气体排放的影响[D]. 武汉: 华中农业大学, 2018: 16–35SUN Z C. Effects of straw returning and feeding on greenhouse gas emissions in rice-crayfish co-culture ecosystem[D]. Wuhan: Huazhong Agricultural University, 2018: 16–35
[32] 李诗豪, 刘天奇, 马玉华, 等. 耕作方式与氮肥类型对稻田氨挥发、氮肥利用率和水稻产量的影响[J]. 农业资源与环境学报, 2018, 35(5): 447–454 LI S H, LIU T Q, MA Y H, et al. Effects of tillage practices and nitrogen sources on NH3volatilization, nitrogen use efficiency and yield in paddy fields in central China[J]. Journal of Agricultural Resources and Environment, 2018, 35(5): 447–454
[33] VAN DER HOEK K W, BOUWMAN A F. Upscaling of nutrient budgets from agroecological niche to global scale[M]/SMALING E M A, OENEMA O, FRESCO L O. Nutrient Disequilibria in Agroecosystems: Concepts and Case Studies. Wallingford: CBA International, 1999: 57–73
Nitrogen and phosphorus cycling characteristics and balance of the integrated rice-crayfish system*
SI Guohan1,2, YUAN Jiafu1,2**, PENG Chenglin1,2, XIA Xiange2, CHENG Jianping3, XU Xiangyu1,2,JIA Ping’an4, XIE Yuanyuan1, ZHOU Jianxiong1
(1. Plant Protection and Soil Fertilizer Institute, Hubei Academy of Agricultural Sciences, Wuhan 430064, China; 2. Key Laboratory of Fertilizer Resource Utilization in Wastes, Ministry of Agriculture and Rural Affairs, Wuhan 430064, China; 3.Food Crops Institute, Hubei Academy of Agricultural Sciences, Wuhan 430064, China; 4. Qianjiang Agro-Technology Extension Center, Qianjiang 433199, China)
The integrated rice-crayfish system is an emerging complex ecological system of planting and breeding in the middle and lower reaches of the Yangtze River in China. The study of N and P cycling characteristics and the profit and loss status of N and P in the integrated rice-crayfish system is of great significance to rationally regulate the nutrient cycling and balancing of the integrated rice-crayfish system and guide the optimal management of the N and P. In this study, a field experiment was conducted to study the cycling characteristics and apparent balance of N and P under the integrated rice-crayfish system using an input-output method, with the rice monoculture system as the control. The results showed that the output/input ratios of N and P in the crayfish subsystem were 0.62 and 0.44, respectively. Inside the crayfish subsystem, the N and P were the largest in the feed input, accounting for 92.9% and 96.4% of the total input, respectively; the output of adult crayfish N and P was the largest, accounting for 53.3% and 59.5% of the total output, respectively. Under the current input levels, the apparent balance of both N and P in the soil subsystems of the two systems were in surplus, and the surplus of N in the soil subsystem of the integrated rice-crayfish system was higher than that of the rice monoculture system, while the surplus of P was lower than that of the rice monoculture system. The N and P output/input ratios of the integrated rice-crayfish system and the rice monoculture system were both less than 1, and the N and P output/input ratio of the integrated rice-crayfish system were less than that of the rice monoculture system. The integrated rice-crayfish system increased the amount of N and P in sequestered soil, and the N and P levels in sequestered soil of the integrated rice-crayfish system were higher than those of the rice monoculture system by 49.2 kg∙hm-2and 9.1 kg∙hm-2, respectively. The integrated rice-crayfish system increased the apparent loss of N and P in the system, and the apparent losses of N and P of the integrated rice-crayfish system were higher than those of the rice monoculture system by 10.2 kg∙hm-2and 1.0 kg∙hm-2, respectively. It can be seen that the integrated rice-crayfish system reduces the output/input ratio of N and P and promotes the accumulation of N and P in the soil but increases the apparent loss of N and P in the system.
Integrated rice-crayfish system; Nutrient cycling; Nutrient balance; Nitrogen and phosphorus
, E-mail: fu1682@sina.com
Jan. 31, 2019;
Mar. 27, 2019
S181
2096-6237(2019)09-1309-10
10.13930/j.cnki.cjea.190088
袁家富, 主要从事植物营养与土壤保育方面研究。E-mail: fu1682@sina.com 佀国涵, 主要从事土壤肥力与养分循环方面的研究。E-mail: siguoh@qq.com
2019-01-31
2019-03-27
* This work was supported by the National Key Research and Development Project of China (2018YFD0301305, 2017YFD0301400, 2016YFD0200807) and the Scientific and Technological Achievements Cultivation Project of Hubei Academy of Agricultural Sciences of China (2017CGPY01).
*表示未考虑其输出N。* means that nitrogen was not included.
* 国家重点研发计划项目(2018YFD0301305, 2017YFD0301400, 2016YFD0200807)和湖北省农业科学院重大研发成果培育项目(2017CGPY01) 资助
佀国涵, 袁家富, 彭成林, 夏贤格, 程建平, 徐祥玉, 贾平安, 谢媛圆, 周剑雄. 稻虾共作模式氮和磷循环特征及平衡状况[J]. 中国生态农业学报(中英文), 2019, 27(9): 1309-1318
SI G H, YUAN J F, PENG C L, XIA X G, CHENG J P, XU X Y, JIA P A, XIE Y Y, ZHOU J X.Nitrogen and phosphorus cycling characteristics and balance of the integrated rice-crayfish system[J]. Chinese Journal of Eco-Agriculture, 2019, 27(9): 1309-1318
猜你喜欢
磷肥与复肥(2022年6期)2022-08-02
当代水产(2022年6期)2022-06-29
广西林业科学(2022年2期)2022-05-09
农业科技与信息(2021年14期)2021-12-05
当代水产(2021年9期)2021-12-02
山东农业科学(2019年11期)2019-12-24
农民致富之友(2019年32期)2019-11-23
当代水产(2019年7期)2019-09-03
江苏农业科学(2019年11期)2019-07-22
福建农业科技(2018年2期)2018-09-10
