
P2X7受体胞外段蛋白的克隆、表达、纯化及鉴定①
2019-10-22 09:31贾丽娜彭效祥李明萱罗淑萍张云芳赵荣兰
中国免疫学杂志 2019年19期
贾丽娜 彭效祥 李明萱 罗淑萍 张云芳 赵荣兰
(潍坊医学院医学检验学系纳米医学技术研究所,潍坊医学院临床检验诊断学山东省“十二五”高校重点实验室,潍坊 261053)
近年来嘌呤能离子通道型受体(Purinergic ligand-gcotedion channel 7 receptor,P2X7R)已成为众多学者广泛关注的热点,由起初在神经系统的初步研究发展到在多个系统炎症性疾病的广泛研究。P2X7R是以ATP为天然配体的非选择性阳离子通道,在组织损伤、感染、肿瘤微环境等伴随炎症发生时,受损细胞和死亡细胞释放ATP使离子通道打开,细胞内外的阳离子浓度以及流动方向发生改变,细胞膜发生去极化使膜的通透性发生改变,引起炎症物质如NO、IL-1β、IL-18、TNF-α等释放进而参与多个系统炎症反应。 P2X7R胞外环上含有ATP结合位点,研究显示抗P2X7R胞外结构域抗体可竞争性结合ATP结合位点从而阻断该受体的激活,如抗P2X7R胞外结构域抗体与P2X7R胞外结构域结合抑制巨噬细胞的死亡;在破骨细胞向多核巨噬细胞的融合过程中也可被抗P2X7R胞外结构域抗体阻断[1,2]。除此之外,利用抗P2X7R胞外段结构域抗体,通过流式细胞技术、免疫组织化学技术及蛋白印迹技术等可检测多种免疫细胞及组织中P2X7R表达情况[3],为研究P2X7R在多种炎症性及免疫性疾病中的作用及机制提供了重要帮助。鉴于上述抗P2X7R胞外段结构域抗体的广泛作用,许多研究者尝试制备抗P2X7R单克隆抗体,但其结合活性不是很理想[4],为此本研究尝试制备高亲和性的抗P2X7R胞外段ATP结合域抗体,为进一步制备抗人P2X7R胞外段单克隆抗体奠定基础。
1 材料与方法
1.1材料
1.1.1主要仪器与试剂 DNA合成仪(Bioauto-maion,中国);ABI 9700 PCR 仪(ABI,美国);核酸电泳仪(北京君意东方电泳设备有限公司,中国);XO-1200D 超声细胞破碎仪(宁波新芝生物科技有限公司,中国);GL21M 高速冷冻离心机(宁波新芝生物科技有限公司,中国);台式高速离心机(Eppendorf,美国),DYY-6C电泳系统(南京大学普阳分析仪器厂,中国);BE-1100 四维旋转混合仪(海门市其林贝尔仪器制造有限公司,中国);QHZ-98A台式全温摇床(太仓市华美生化仪器厂,中国);甲叉双丙烯酰胺、丙烯酰胺、四甲基乙二胺、过硫酸铵(Sigma,美国);二硫苏糖醇(DTT)、苯甲基磺酰氟(PMSF)、还原型谷胱甘肽(GSH)、(氧化型)谷胱甘肽(GSSG)、三羟甲基氨基甲烷(Tris)、十二烷基硫酸钠(SDS)、乙二胺四乙酸(EDTA)、氯化钠(NaCl)、甘油(Glycerol)、咪唑(Imidazole)(BBI,美国);质粒提取试剂盒、DNA 回收试剂盒(天根生化科技有限公司,中国);T4 DNA 连接酶、高保真 DNA 聚合酶[生物工程(大连)有限公司,中国];Bradford 蛋白浓度测定试剂盒(上海碧云天生物技术有限公司,中国);Ni-IDA琼脂糖凝胶、ECL 试剂盒、SDS-PAGE 和 WB Marker[德泰生物科技(南京)有限公司,中国]。
1.2方法
1.2.1P2X7R胞外段蛋白编码序列的扩增 根据GenBank收录的人P2X7R基因(GenBank 登记号:NP_002553.3)序列,确定其胞外段包含ATP结合位点的编码序列区域,采用德泰生物的密码子优化软件 MaxCodonTMOptimization Program (V13)对锁定的P2X7R胞外段蛋白氨基酸编码序列进行优化,共设计14条引物,引物序列见表1,在第1条和第14条引物设计时分别加入NdeⅠ和HindⅢ酶切位点,使用高保真DNA聚合酶进行重叠PCR扩增。第一轮全长PCR扩增:反应体系中加入全部14条引物,扩增参数为95℃预变性3 min;95℃变性25 s,60℃退火20 s,72℃延伸40 s,25个循环;72℃延伸 1 min。第二轮全长PCR扩增:上述第一轮PCR产物为模板,以第1和第14条引物为上下游引物,扩增参数同第一轮PCR扩增参数。琼脂糖凝胶电泳后回收第二轮PCR产物,用于后续重组表达质粒的构建。
表1 引物序列
Tab.1 Primers used for overlapping PCR
2.2P2X7R胞外段蛋白重组表达质粒构建及鉴定 用NdeⅠ和HindⅢ双酶切P2X7R胞外段蛋白(P2X7R蛋白)编码序列的PCR产物和表达载体pET30a(+),琼脂糖凝胶电泳后,回收纯化双酶切后的PCR产物及线性化载体pET30a(+),T4 DNA连接酶4℃ 连接过夜,连接产物转化至TOP10感受态细胞。将转化后产物接种至含50 μg/ml卡那霉素的LB琼脂平板上,37℃培养16 h,挑取单个菌落,使用T7/T7 terminator 引物进行PCR鉴定,阳性菌落过夜摇菌后提取质粒,酶切鉴定及测序分析,成功构建pET30a-P2X7R表达质粒。
2.3P2X7R蛋白诱导表达 将测序正确的pET30a-P2X7R重组质粒转化至E.coli BL21(DE3+)表达菌株中,挑取阳性单克隆于含50 μg/ml的卡那霉素LB 培养基中。 37℃培养过夜,次日按1∶100接种至4 ml新鲜 LB培养基培养至 OD600 为 0.6,向试管培养液中加入不同终浓度IPTG,筛选出最佳诱导剂的浓度为0.2 mmol/L IPTG及最佳诱导时间为16 h。在新鲜接种的菌液OD值为0.6时,加入0.2 mmol/L IPTG,分别在15℃和37℃诱导16 h,分别取等量不同温度诱导的菌液,SDS-PAGE分析鉴定蛋白的诱导表达。
2.4P2X7R蛋白产物纯化 收集大量培养后的菌体,加入20倍体积(v/m)的缓冲液[20 mmol/L PB(pH7.2)、300 mmol/L NaCl、20 mmol/L Imidazole含1% Triton X-100、1 mmol/L DTT、1 mmol/L PMSF]重悬菌体。置冰浴中超声碎菌后,12 500 r/min 4℃离心15 min,离心后分别收集上清和沉淀。碎菌后的沉淀(包涵体)缓冲液(20 mmol/L pH7.2 PB、300 mmol/L NaCl含1% Triton X-100、5 mmol/L DTT)洗涤后,使用包涵体溶解液(20 mmol/L pH7.2的PB、300 mmol/L NaCl、8 mol/L Urea、20 mmol/L 咪唑、1 mmol/L DTT)溶解包涵体后,12 500 r/min 离心收集上清。将上述两步中收集到的上清分别以0.45 μm滤膜过滤后;分别加入平衡后的Ni-IDA柱中,旋转混合仪4℃孵育80 min;将带有His标签的重组蛋白以 50、100、300 mmol/L的咪唑缓冲液洗脱,分别取不同浓度咪唑洗脱的重组P2X7R蛋白进行SDS-PAGE,检测纯化后的蛋白。
2.4Western blot鉴定表达产物 重组P2X7R蛋白样品进行SDS-PAGE后,再湿转至硝酸纤维素膜上,5%的脱脂奶粉室温封闭2 h,1∶5 000稀释鼠源性的抗His标签单克隆抗体,4℃孵育过夜;TBST洗涤3次后,加入1∶6 000稀释的HRP标记的山羊抗鼠的二抗,室温摇床孵育2 h,TBST洗3次后,进行化学发光检测。
2 结果
2.1P2X7R胞外段蛋白编码序列的扩增结果 重叠PCR扩增产物的琼脂糖凝胶电泳出现约900 bp左右的条带,与预期扩增的P2X7R胞外段蛋白编码序列片段长度相符(图1)。
2.2P2X7R重组表达质粒构建及鉴定 P2X7R胞外段蛋白编码序列与pET30a(+)质粒分别经NdeⅠ和HindⅢ双酶切,T4 DNA连接酶连接过夜后,转化至Top10克隆菌株。挑取单个阳性菌落培养后PCR鉴定(图2A)及提质粒后酶切鉴定(图2B),琼脂糖凝胶电泳,出现约900 bp的插入片段,与预期结果一致。

图1 P2X7R胞外段蛋白编码序列重叠PCR扩增产物琼脂糖凝胶电泳鉴定结果Fig.1 Agarose gel electrophoresis showing overlap PCR amplification of P2X7R extracellular segment protein coding sequenceNote: Lanes 1 and 2.PCR amplification product;Lane 3.DL5000.
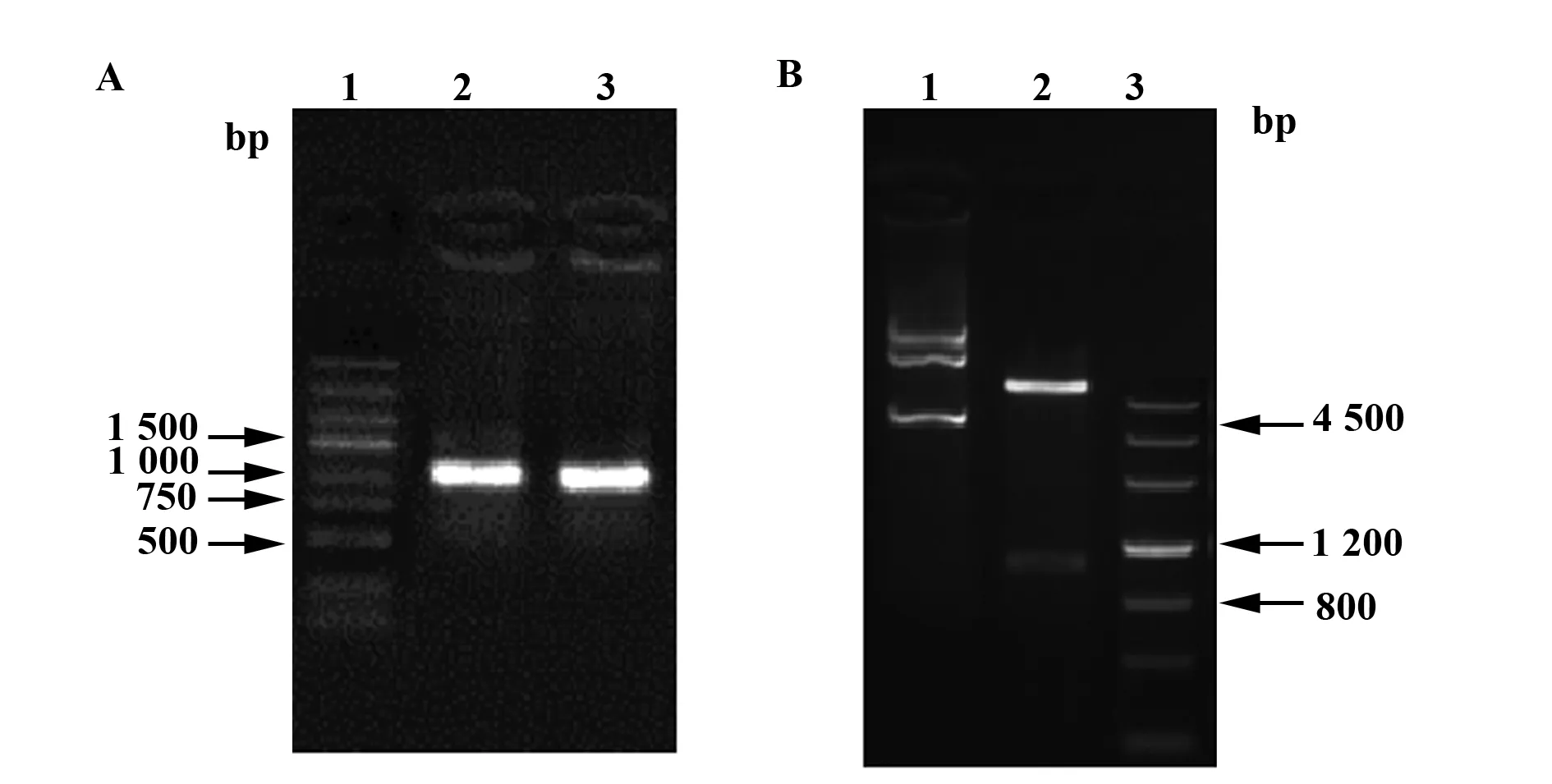
图2 P2X7R重组表达质粒PCR鉴定和酶切鉴定Fig.2 Agarose gel electrophoresis for confirmation of clones by PCR and restriction enzyme digestionNote: A.PCR identification of pET30a-P2X7R recombinant plasmid.Lane 1.DL5000;Lanes 2 and 3.Amplified products.B.Restriction enzyme digestion identification of pET30a-P2X7R recombinant plasmid.Lane 1.pET30a-P2X7R recombinant plasmid;Lane 2.pET30a-P2X7R recombinant plasmid digested by NdeⅠ and HindⅢ;Lane 3.DL4500.
2.3重组P2X7R蛋白的诱导表达 将pET30a-P2X7R重组质粒转化至E.coli BL21(DE3+)后,经0.2 mmol/L IPTG分别于15℃和37℃诱导表达16 h,分别取1 ml菌液离心取沉淀,PBS重悬后加入SDS-PAGE 上样缓冲液后,于 100℃下加热样品10 min,离心取上清电泳。SDS-PAGE结果显示,诱导表达的重组蛋白分子量为35 kD左右(图3),与预期结果相符。

图3 SDS-PAGE分析重组P2X7R蛋白的诱导温度Fig.3 SDS-PAGE analysis of expression of recombinant P2X7R protein with induction temperatureNote: Lane 1.Protein marker;Lane 2.Empty bacteria of BL21(DE3+);Lane 3 and 4.Expression of recombinant P2X7R protein in 16 hours at 15℃ and 37℃.

图4 SDS-PAGE 分析超声碎菌上清中重组P2X7R蛋白纯化结果Fig.4 SDS-PAGE analysis of purification of recombinant P2X7R protein in supernatant of broken bacteriaNote: Lane 1.Supernatant of broken reconstitution bacterial;Lane 2.Effluent of supernatant incubated on Ni-IDA affinity chromatographic column;Lane 3.Protein marker;Lane 4.Recombinant P2X7R protein eluted by 50 mmol/L imidazole buffer;Lanes 5-7.Recombinant P2X7R protein eluted by 100 mmol/L imidazole buffer;Lanes 8-10.Recombinant P2X7R protein eluted by 300 mmol/L imidazole buffer.
2.4重组P2X7R蛋白纯化 超声碎菌离心后,分别收集碎菌后的上清和沉淀(包涵体);包涵体溶解液溶解后的包涵体与碎菌后收集的上清,分别经0.45 μm滤膜过滤,经Ni-IDA亲和层析柱纯化,最后分别以 50、 100、 300 mmol/L咪唑缓冲液洗脱,经超滤获得纯化的重组P2X7R蛋白。SDS-PAGE结果显示,100 mmol/L咪唑缓冲液洗脱效果最好,重组蛋白主要存在于包涵体中(图4、5)。
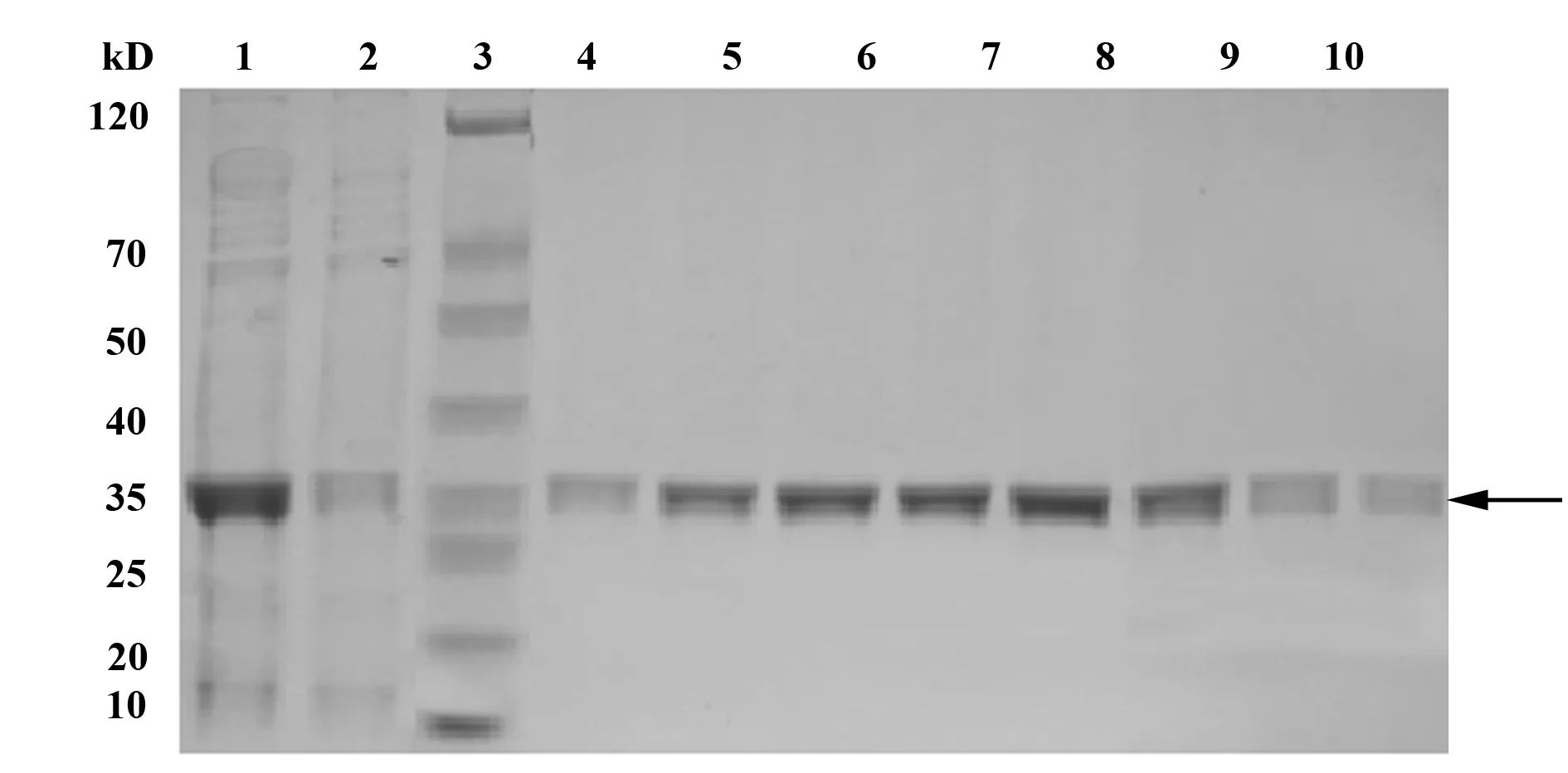
图5 SDS-PAGE 分析超声碎菌后包涵体中重组P2X7R蛋白纯化结果Fig.5 SDS-PAGE analysis of purification of recombinant P2X7R protein in inclusion body of broken bacteriaNote: Lane 1.Supernatant after dissolution of inclusion bodies;Lane 2.Effluent of supernatant incubated on Ni-IDA affinity chromatographic column;Lane 3.Protein marker;Lane 4.Recombinant P2X7R protein eluted by 50 mmol/L imidazole buffer;Lanes 5-7.Recombinant P2X7R protein eluted by 100 mmol/L imidazole buffer;Lanes 8-10.Recombinant P2X7R protein eluted by 300 mmol/L imidazole buffer.

图6 重组P2X7R蛋白的Western blot鉴定分析Fig.6 Western blot analysis of recombinant P2X7R proteinNote: Lane 1.Protein marker;Lane 2.Recombinant P2X7R protein eluted by 100 mmol/L imidazole buffer.
2.5Western blot鉴定重组P2X7R蛋白 将100 mmol/L 咪唑缓冲液洗脱的重组P2X7R蛋白经SDS-PAGE分离后,转至硝酸纤维素膜上,Western blot结果显示,抗His标签单克隆抗体能特异性识别35 kD左右的条带(图6)。
3 讨论
P2X7R广泛分布于T淋巴细胞、巨噬细胞、树突状细胞、肥大细胞等体内多种免疫细胞上,对多个系统的炎症性疾病起调控作用。P2X7R是以配体ATP为内源天然性激活剂的非选择性阳离子通道。生理状态下,胞外ATP处于较低的pmol/L~nmol/L水平;病理状态时,胞内ATP释放至胞外,当其浓度大于100 μmol/L时,部分通道开放,K+外流、Ca2+、Na+内流,膜通透性增加释放炎性因子介导炎症反应,当在高浓度ATP持续刺激时,形成非选择性质膜孔道(Pannexinal,Panx1),允许大分子量的物质进入胞内引起细胞发生凋亡[5,6]。
P2X7R调控炎性疾病的主要信号通路是发生在P2X7R激活后,如由K+外流引起的胞内低K+可激活IL-1β转化酶,P2X7R激活可活化NLRP3使半胱氨酸蛋白酶-1(Caspase-1)激活,而IL-1β转化酶和Caspase-1都可促使炎介质IL-1β和IL-18转变成具有生物学功能的成熟体[7,8]。研究发现,将单核和巨噬细胞用LPS做预处理,在激活P2X7R后可使MAPK、NF-κB等信号通路开放,引起Caspase-1活化使IL-1β和IL-18分化成熟,P2X7R激活可使巨噬细胞生成活性氧(ROS),在核苷酸受体介导的p38和JNK信号通路激活中ROS也发挥重要作用[9]。
大量研究显示,P2X7R与多系统炎症性疾病的发生发展密切相关。①呼吸系统疾病:在小鼠急慢性过敏性气道炎症、哮喘以及香烟烟雾诱导的慢性阻塞性肺气肿和肺部炎症等呼吸系统疾病中,P2X7R表达明显上调[10-12]。②消化系统疾病:在腹膜注射雨蛙肽诱导的小鼠慢性胰腺炎模型中,发现P2X7R拮抗剂氧化ATP和亮蓝-G组与正常对照组相比,胰腺组织中P2X7R、NLRP3和Caspase-1在基因和蛋白水平表达均显著下降,P2X7R拮抗剂组胰腺慢性炎症和纤维化显著减弱,结果提示阻断P2X7R信号通路可能成为治疗慢性胰腺炎及其纤维化的新策略[13]。Panx1存在于许多类型细胞中,此通道激活后释放的ATP可激活 P2X7R诱导炎症性肠病的发生[14]。③心血管系统疾病:动脉粥样硬化是一种慢性炎症,同型半胱氨酸(Homocysteine,Hcy)是心血管疾病的危险因素,病理水平的Hcy可诱导巨噬细胞高表达P2X7R,通过ATP/NLRP3信号通路使IL-1β分化成熟并参与动脉粥样硬化的发展[15]。在冠状动脉粥样硬化性心脏病引起的心肌梗死(Myocardial infraction,MI)灶周围,发现P2X7R表达增加并激活NF-κB信号通路,导致MI后室性心律失常易感性增加[16]。在血管紧张素Ⅱ依赖性高血压时,P2X7R可引起肾小球血管阻力增加,尿钠排泄障碍[17]。④神经系统疾病:P2X7R在脑外伤、阿尔茨海默症、脊髓外伤和神经性疼痛等疾病中发挥重要作用[18-20]。除此之外,P2X7R还与一些肿瘤疾病的发展密切相关,P2X7R激活可提高线粒体的氧化磷酸化效率,同时增加乳酸含量,上调几乎所有参与糖酵解的酶和转运体,从而促进肿瘤细胞的增殖和存活;P2X7R激活可增强PC9肺癌和 T47D乳腺癌细胞的迁移,可使前列腺癌细胞的迁移性和侵袭性增加,也可通过JNK途径促进肝癌细胞的增殖,使用P2X7R抑制剂和基因沉默P2X7R都可抑制肿瘤细胞的增殖,另外PI3K/Akt通路也参与P2X7R依赖性肿瘤细胞的生长、侵袭性、转移性扩散和血管生成[21]。P2X7R在乳腺癌细胞系和组织中过度表达,P2X7R-shRNA表达载体有效抑制人乳腺癌细胞MCF-7中P2X7R的表达,并诱导MCF-7凋亡和增殖水平降低[22]。在淋巴瘤的研究中,通过基因沉默小鼠淋巴样瘤细胞P388D1中P2X7R可使T复合多肽1(Tailless complex polypeptide 1,TCP-1)下降,而下调TCP-1可以抑制体内淋巴瘤细胞的转移[23]。血清饥饿可提高神经母细胞瘤细胞中P2X7R的表达,有助于维持癌细胞即使在没有营养支持的情况下也处于增殖状态,而PI3K/Akt途径抑制可阻止血清饥饿后神经母细胞瘤细胞中P2X7R的表达[24]。P2X7R在人胰腺导管腺癌(Pancreatic ductal adenocarcinoma,PDAC)细胞高表达,可促进人PDAC癌细胞的增殖和迁移,P2X7R抑制剂AZ10606120可降低胰腺癌侵袭性[25]。
鉴于P2X7R在多系统多疾病中发挥重要作用,许多研究者尝试利用抗P2X7R抗体去阻断该通道的打开或者检测组织细胞中该受体的表达[1-3,26],借以研究P2X7R在不同疾病中的作用,因此抗P2X7R 抗体成为研究P2X7R与多种炎症疾病关系的重要工具。现有抗P2X7R抗体除结合活性受限外,其售价较高,直接限制了该抗体在体内外实验研究中的应用,因此本研究致力于制备高亲和性及低成本的抗P2X7R抗体,而本研究中P2X7R胞外段蛋白的成功克隆表达,为今后制备抗人P2X7R胞外段单克隆抗体奠定了坚实的基础,也为进一步深入研究P2X7R相关疾病的诊断和治疗提供了重要的工具和思路。
猜你喜欢
广东药科大学学报(2022年3期)2023-01-04
生物学通报(2022年1期)2022-11-22
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
成都医学院学报(2022年4期)2022-08-19
中国农学通报(2022年12期)2022-06-01
临床肺科杂志(2021年4期)2021-12-23
中国种业(2021年11期)2021-11-25
现代临床医学(2021年4期)2021-07-31
江西农业学报(2021年4期)2021-04-20
北京大学学报(医学版)(2020年6期)2020-12-14
