
组蛋白乙酰化对灵芝生长、灵芝多糖和灵芝酸生物合成的影响
2020-02-25 08:25张宗源蒋咏梅章文贤
中国农业科学 2020年3期
张宗源,蒋咏梅,章文贤
组蛋白乙酰化对灵芝生长、灵芝多糖和灵芝酸生物合成的影响
张宗源1,2,蒋咏梅1,章文贤1
(1福建师范大学生命科学学院,福州 350108;2河南省科学院生物研究所有限责任公司,郑州 450008)
【目的】组蛋白乙酰化修饰对真菌生长发育和次级代谢合成具有重要的调控作用。研究组蛋白乙酰化对静置培养的灵芝生长发育和主要代谢物合成的影响,为表观遗传手段提高灵芝多糖和灵芝酸生物合成提供理论依据。【方法】采用液体振荡-静置两阶段法培养灵芝。在静置培养阶段添加不同浓度(0、0.6、60、120和180 μmol·L-1)辛二酰苯胺异羟肟酸(SAHA),采用常规方法测定或观察灵芝生物量、糖消耗、上层菌丝膜形成、菌丝体形态、灵芝孢子生成以及灵芝酸和灵芝多糖生物合成,利用蛋白免疫印迹技术测定灵芝组蛋白乙酰化水平;利用qRT-PCR技术对灵芝多糖(和)、灵芝酸(、、和)生物合成关键基因以及灵芝全局调控因子相关基因(和)进行表达分析。【结果】SAHA处理使灵芝组蛋白H4乙酰化水平提高,最高为对照组的1.6倍;抑制了灵芝菌丝体生长和色素产生,改变了菌丝体的形态。灵芝孢子的形成也受到抑制,且SAHA浓度越大,抑制程度越明显。SAHA处理显著增加了灵芝多糖的产量,最高增加50%;灵芝酸生物合成受到抑制,与对照相比降低13%—27%;qRT-PCR分析结果表明SAHA处理下灵芝多糖与灵芝酸合成关键酶基因表达均有不同程度上调,其中灵芝多糖合成关键酶基因提升1.5—3.5倍,灵芝酸合成关键酶基因提升1.8—12.1倍;灵芝全局性调控因子和的表达被抑制,仅为对照组的11.30%—62.4%。【结论】组蛋白乙酰化可通过灵芝全局调控因子调控灵芝生长发育进而影响灵芝酸生物合成,同时组蛋白乙酰化对灵芝多糖生物合也有影响。
灵芝;组蛋白乙酰化;灵芝酸;灵芝多糖;SAHA
0 引言
【研究意义】灵芝()是我国中医药宝库中的珍品,具有重要的药用价值和保健功能。研究表明灵芝酸(GA)[1]与灵芝多糖(胞外多糖EPS、胞内多糖IPS)[2]是其主要的药用成分,其中灵芝酸因具有抗癌、抗HIV等功效,灵芝多糖具有抗氧化、免疫调节等作用而受到越来越多研究者的关注[3-4]。近期研究表明,灵芝多糖还作为植物免疫诱抗剂增加番茄植株对灰霉病抗性[5]和小麦对纹枯病的抗性[6]。FANG[7]等发现液体振荡-静置两阶段培养法与传统的液体发酵和固体培养灵芝子实体等方法相比,能显著提高灵芝活性物质的产量。近年来,随着对灵芝酸和灵芝多糖的应用需求,进一步提高其产量很有必要。真菌表观遗传研究发现,组蛋白乙酰化不仅能够影响菌落建成[8]、光响应[9-10],而且还调控真菌的有(无)性发育[11-12]、次级代谢产物合成[13-15],并在维持细胞周期[16]、环境适应[17]等方面也起着至关重要的作用。化学表观遗传抑制剂的使用为真菌生长发育和代谢的表观遗传研究提供了新的策略[18-19],同时也可成为增加真菌代谢产物合成的强有力手段。【前人研究进展】据邓茂常[20]的文献记载,我国于20世纪60年代末已经开展灵芝液体深层发酵技术的研究。在接下来的10多年里,研究的侧重点主要为发酵过程中灵芝菌丝体形态的变化和如何获得更多的灵芝生物量[21]。90年代至今,在提高灵芝酸和灵芝多糖生产方面已有大量的文献报道,主要致力于灵芝深层培养的最佳条件、最佳培养基配方、接种量以及反应器的选择等,以提升灵芝多糖和灵芝酸的产量[22-24]。2002年,FANG等[7]首次报道了一种液体振荡发酵与液体静置培养两阶段培养法,使灵芝酸生物合成大大提高,随后有大量研究在此基础上展开,包括在静置培养中添加P450诱导剂苯巴比妥[25]、添加钙离子和镁离子或锰离子[26]以及静置培养过程中氮源限制[27]和调节气相氧浓度[28]对灵芝酸合成都有提高作用。刘高强等[4]以14C标记的葡萄糖作为碳源初步推断出灵芝胞外多糖IPS-1合成基本途径;JI等[29]通过基因工程手段高表达灵芝同源基因,提高了灵芝多糖产量;刘柯等[30]通过异源表达透明颤菌血红蛋白基因也获得了高产灵芝多糖菌株。组蛋白乙酰化修饰是真菌表观遗传研究的主要内容之一。组蛋白乙酰化平衡主要依靠组蛋白乙酰转移酶(histone acetyltransferases,HATs)和组蛋白去乙酰化酶(histone deacetylase,HDAC)维持。SAHA是一种化学表观遗传抑制剂,可抑制HDAC活性,打破组蛋白乙酰化水平,改变基因的转录从而影响真菌的生长和代谢。WILLIAMS等[31]利用组蛋白去乙酰化抑制剂和DNA甲基化抑制剂作用12种真菌,其中一些真菌的次级代谢物产量提高而且还分离得到结构新颖的次级代谢物。在黑曲霉研究中,SAHA添加改变了次级代谢物合成基因的表达水平[32]。BASIMIA等[18]通过SAHA处理,严重抑制了黄曲霉毒素生物合和孢子的产生。【本研究切入点】目前灵芝的发酵研究主要侧重于发酵条件优化、添加激发子、信号转导工程以及基因工程菌构造等策略,以促进灵芝生长,提高代谢物合成,而组蛋白乙酰化的表观遗传有待进一步研究。【拟解决的关键问题】本研究通过SAHA处理,研究组蛋白乙酰化对灵芝生长、菌丝形态、孢子分化以及灵芝酸和灵芝多糖代谢合成的影响,并进一步在分子水平探讨SAHA处理后,组蛋白乙酰化对灵芝发育及灵芝酸和灵芝多糖合成的作用机理,为解析灵芝表观遗传调控其代谢的分子作用奠定基础,为提高灵芝酸和灵芝多糖产量提供新思路。
1 材料与方法
试验于2015年9月至2016年12月在福建师范大学国家教育部工业微生物研究工程中心进行。
1.1 供试菌株及处理
灵芝[(Leyss. ex Fr. ) Karst]由福建师范大学国家教育部工业微生物研究工程中心提供。液体振荡-静置两阶段培养灵芝,具体培养方法与步骤见参考文献[7],将SAHA(上海蓝木化工有限公司)溶解在DMSO(北京鼎国昌盛有限公司)中,配置为200 mmol∙L-1的母液,用MillporeExpress膜针头式过滤器过滤除菌。静置培养阶段,在发酵液中加入不同体积的SAHA母液,使其终浓度分别为0(空白对照组)、0.6、60、120和180mmol∙L-1,并设置溶剂对照组(0.09% DMSO)。试验组与对照组各设置3个重复。
1.2 灵芝菌丝表型特征分析
观察记录不同培养时间、不同处理的灵芝白色菌丝膜形成过程,通过显微镜观察灵芝菌丝体形态及灵芝孢子。按照ZHANG等[28]的方法收集灵芝孢子,使用血球计数法测定孢子数目。
1.3 生物量及残糖量测定
收取不同培养时间的灵芝白色菌丝膜细胞,于50℃烘箱中烘干至恒重,称量生物量,同时收集发酵液12 000×条件下离心15 min,取上清液采用DNS法测定残糖量。灵芝细胞平均生长速率(g∙L-1∙d-1)=最大生物量(g∙L-1)/达到最大生物量的天数(d)。
1.4 灵芝酸、灵芝多糖产量及灵芝组蛋白乙酰化水平测定
参照TANG等[33]的方法测定灵芝胞外多糖产量和胞内多糖含量。参照蓝丽雯[34]的方法提取并测定总灵芝酸含量。按照胡彬彬等[35]的方法提取灵芝菌丝体蛋白,使用蛋白质免疫印迹法测定灵芝组蛋白H3、H4乙酰化水平,一抗:Histone H3ac (pan-acetyl) antibody,Histone H4ac (pan-acetyl) antibody(active motif),内参抗体:Histone H3 antibody(ABclonal);二抗:Hrp Goat anti-Rabbit IgG(H-L)(ABclonal),样品送厦门泰京生物有限公司进行测定。
1.5 灵芝多糖、灵芝酸生物合成关键酶及全局调控因子相关基因表达水平
按照北京鼎国昌盛生物技术有限公司TRigol试剂的说明书提取灵芝总RNA,测定其OD260/280值,取1 μL进行1%琼脂糖凝胶电泳分析提取的RNA纯度及检测其完整性。以此RNA为模板,用北京全式金生物技术有限公司的TransScript One-Step gDNA Removal and cDNA Synthesis SuperMix反转录试剂盒合成灵芝cDNA。灵芝酸生物合成关键酶基因、灵芝全局调控因子基因及内参基因参考参考文献[34]和ZHANG等[36];灵芝多糖生物合成关键酶基因荧光定量PCR引物及内参基因参考JI等[29](表1)。使用美国ABI公司的Prism 7300定量PCR仪和DBI® Bioscience公司的Bestar® SybrGreen qPCR Mastermix 定量PCR试剂盒进行基因表达测定。荧光定量PCR的程序参考蓝丽雯[34]的方法,表达量的计算分析采用LIVAK等[37]的方法。

表1 qRT-PCR引物
1.6 统计学分析
使用SPSS软件进行差异显著性分析。不同小写字母表示差异显著(<0.05)。
2 结果
2.1 SAHA对灵芝细胞生长、葡萄糖消耗和组蛋白乙酰化水平的影响
图1显示了不同浓度SAHA 处理与溶剂对照中灵芝细胞生长和葡萄糖消耗的动态过程。由图可知,随着发酵的开始,各试验组的灵芝细胞生物量均先升高而后降低。SAHA处理后细胞最高生物量(干重,DW)在20.0—20.8 g∙L-1,均低于对照组(如无特殊说明均指空白对照组,下同)((22.3±0.30)g∙L-1,6 d)和DMSO组((21.2±0.30)g∙L-1,6 d);处理组的平均生长速率在3.42—2.22 g∙L-1∙d-1,低于对照组((3.71±0.04)g∙L-1∙d-1)和DMSO组((3.54±0.04)g∙L-1∙d-1)3.3%—39.6%。灵芝细胞对葡萄糖消耗与灵芝细胞生长情况相吻合(图1),随着SAHA浓度的升高,灵芝细胞对葡萄糖的消耗越来越慢,进而影响了灵芝细胞生物量。对照组灵芝细胞的葡萄糖消耗在第3—6天时最快,第9天时葡萄糖消耗殆尽;处理组中灵芝细胞糖耗速率均低于对照组,其中180 μmol·L-1SAHA处理组在整个培养过程中,葡萄糖消耗速率较为平缓。图2比较了灵芝细胞的对照组和SAHA处理组(60 μmol·L-1和180 μmol·L-1)在静置培养1 d和12 d的生长情况,由图可见,培养1 d可在平板表面观察到一层白色菌丝体膜(图2),对照组的白色菌丝体膜更早出现且更致密,培养12 d的对照组已有明显的色素出现,而处理组尚不明显。可见,SAHA处理对培养前期白色菌丝体膜的形成和培养后期灵芝细胞色素的产生有一定抑制作用,且浓度越高效果越明显。
通过蛋白免疫印迹(western blotting,WB)分析培养9 d时0、60和180 μmol·L-1SAHA处理下灵芝细胞组蛋白乙酰化水平的差异,结果如图3-A所示。SAHA添加导致组蛋白H4乙酰化水平的提高,利用Image J软件对免疫印迹条带灰度分析,结果如图3-B所示,60和180 μmol·L-1SAHA处理组组蛋白H4乙酰化水平分别是对照组的1.4、1.6倍,而组蛋白H3乙酰化水平没有差异。可见,组蛋白乙酰化修饰对灵芝细胞生长以及细胞色素的生产起重要作用。
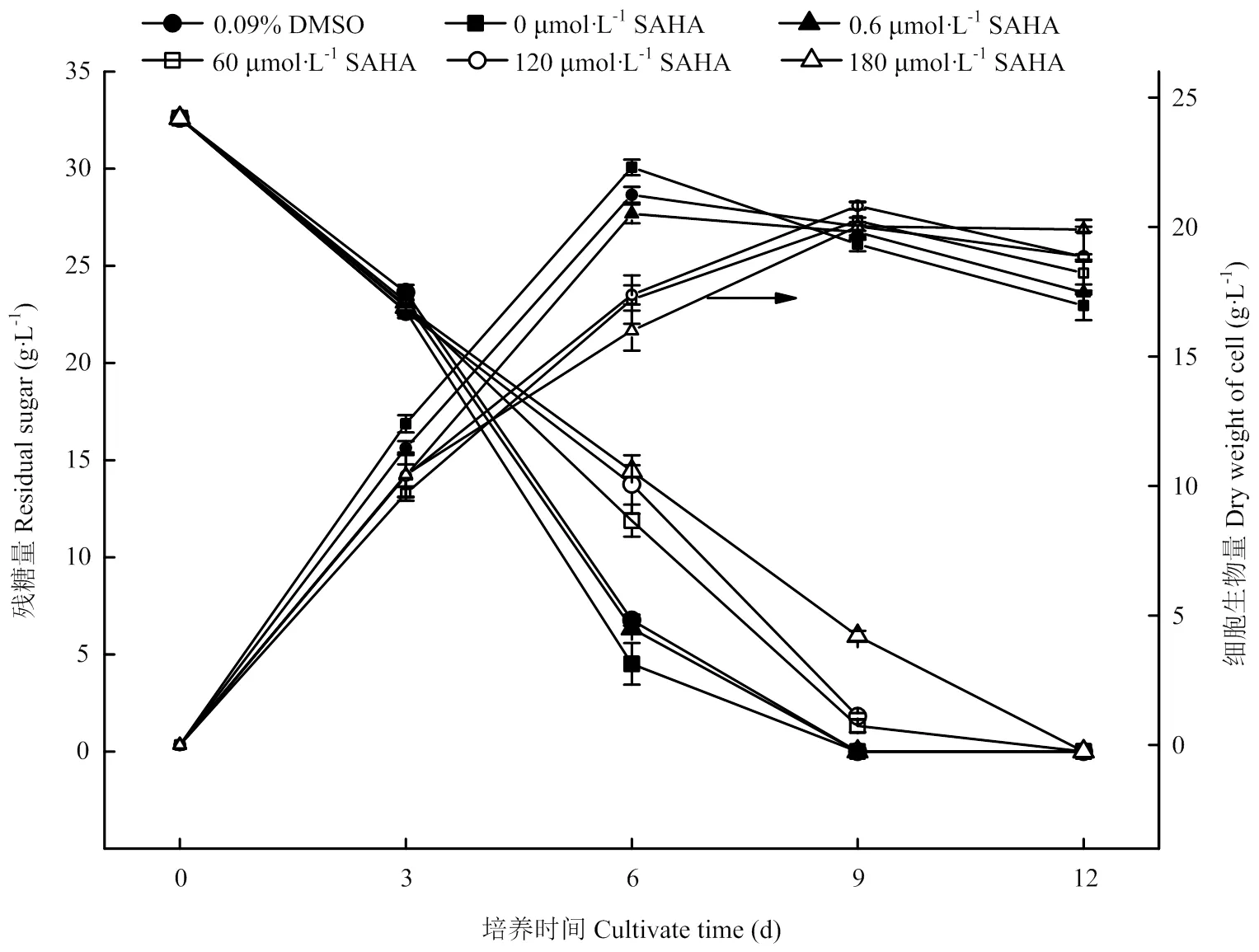
图1 SAHA对灵芝生物量及糖消耗的影响

图2 SAHA对灵芝白色菌丝膜生长的影响
2.2 SAHA对灵芝菌丝和孢子数的影响
对不同SAHA浓度处理的灵芝细胞的气生菌丝进行显微镜观察,结果见图4。SAHA处理后,菌丝体形态发生改变,如图4-A(Ⅰ—Ⅲ)所示:对照组灵芝菌丝体整体呈直线分支状,而在SAHA浓度为120和180 μmol·L-1时,呈无规则卷曲状。在静置培养过程中,对照组(0 μmol·L-1SAHA组和DMSO组)与试验组均能够分化出孢子(图4-A(Ⅳ—Ⅸ),通过计数发现试验组灵芝孢子数目减少。由图4-B可知,灵芝细胞的产孢量随着静置培养时间的增加而增加。但相对于对照组来看,灵芝细胞的产孢量随着抑制剂浓度的增加而减少。第12天时,对照组的产孢量分别是0.6、60、120和180 μmol·L-1SAHA处理组的1.49、1.75、1.9和2.9倍。可见,SAHA添加改变灵芝组蛋白乙酰化水平,进而影响了灵芝菌丝体发育。
2.3 SAHA对灵芝酸和灵芝多糖合成的影响
SAHA处理提高了灵芝组蛋白乙酰化水平,对灵芝代谢产生不同影响。图5为不同处理对总灵芝酸产量的影响,由图可知,培养12 d时,对照组的总灵芝酸产量达到最大值,为(0.77±0.05)g∙L-1。试验组(0.6、60、120和180 μmol·L-1SAHA)中总灵芝酸产量最大值分别为(0.58±0.03)(9 d)、(0.67±0.01)(9 d)、(0.56±0.05)(9 d)、(0.56±0.04)(12 d)g∙L-1,分别比对照组降低25%、13%、27%和27%。同时也测定了灵芝多糖产量(9 d),胞外多糖(EPS)和胞内多糖(IPS)产量在SAHA的作用下都有所增加,结果如图6所示。0.6、60、120和180 μmol·L-1SAHA试验组的灵芝多糖产量(EPS+IPS)分别比对照组((4.91±0.13)g∙L-1)增加了约33%、44%、47%和50%。

A:灵芝组蛋白免疫印迹;B:灵芝组蛋白乙酰化水平比较。不同小写字母表示差异显著(P<0.05)。下同

图5 SAHA对灵芝酸产量的影响

图6 SAHA对灵芝多糖总产量的影响
2.4 SAHA对灵芝多糖和灵芝酸生物合成关键酶基因表达的影响
图7为SAHA对灵芝多糖生物合成关键酶基因()的影响,由图可知,SAHA处理后灵芝多糖的3个关键酶基因的表达量均有所提高,当SAHA浓度为60 μmol·L-1时,3个基因的表达量分别为对照组的2.6倍、3.5倍和2.5倍,180 μmol·L-1处理时为2.1倍、3.2倍和1.5倍。图8为灵芝酸生物合成关键酶基因()的表达情况,由图可知SAHA的处理提高了灵芝酸生物合成关键酶基因的表达水平,当SAHA浓度为60 μmol·L-1时,4个基因的表达量分别为对照组的5.8倍、2.0倍、8.8倍和2.1倍,180 μmol·L-1处理时分别为对照的4.2倍、2.1倍、12.1倍和1.8倍。
2.5 SAHA对灵芝vet和LaeA表达的影响
为了进一步探究SAHA对灵芝生长发育和代谢的影响,测定了和转录水平。从图9可以看出,60和180 μmol·L-1SAHA处理组培养至第9天时,灵芝和表达量均有不同程度降低,其中表达量分别为对照的62.4%和11.3%,表达量分别为对照的45.5%和24.2%。和表达量的降低,表明SAHA诱导的灵芝细胞形态、孢子数量和GA产量的变化可能受及转录水平变化的影响。

图7 SAHA对ugp、gls和pgm表达的影响

图8 SAHA对hmgr、sqs、se和ls表达的影响
3 讨论
组蛋白乙酰化修饰在真核生物生长周期中扮演着重要角色,其动态平衡控制着染色体状态,与基因的沉默和抑制密切相关。SAHA属于异羟肟酸类修饰剂,可抑制Ⅰ、Ⅱ类HDACs,提高组蛋白乙酰化水平,从而影响生物体的生命过程。研究表明,组蛋白乙酰化与真菌的生长和发育息息相关,通过添加SAHA抑制HDAC,橘青霉菌落颜色加深,同时分生孢子梗和分生孢子数目也增加,孢子聚集密实,周边的孢子部分有出芽现象[38];而黄曲霉气生菌丝变成毛绒状,有较小的分生孢子头,随着SAHA浓度增加(2—50 μmol·L-1),孢子数目越来越少,且菌落颜色变浅[18]。在本研究中,静置培养时,SAHA的处理提高了灵芝组蛋白H4乙酰化水平,导致灵芝生长延迟且抑制了糖耗速率和细胞生长速率;随着SAHA浓度增加,其生物量逐渐降低,孢子数目减少。同时显微镜观察SAHA处理后灵芝气生菌丝形态发生变化,在生长后期抑制了上层白色菌丝体膜色素产生。
组蛋白乙酰化在真菌次级代谢产物调控中发挥着重要作用[13-15],通过曲古抑菌素A(TSA,一种HDAC抑制剂)[39]抑制黄曲霉HDAC,黄曲霉毒素生物合成受到抑制,检测黄曲霉产毒基因,除外,、和转录下调。LEE等[40]通过敲除烟曲霉(Ⅱ类HDAC基因)提高了几种次级代谢产物的生物合成,而胶霉毒素的合成则受到抑制,测定非核糖体多肽合成途径14个相关基因的表达发现9个上调。本研究中,通过抑制HDAC,增加了组蛋白H4乙酰化水平,对灵芝的代谢产物也产生了不同的影响,其中灵芝多糖合成增加,其关键酶基因表达上调;灵芝酸生物合成则受到抑制,但其关键酶基因表达上调。现有信息表明灵芝酸生物合成途径上游过程基本清楚,但从羊毛甾醇之后灵芝酸合成的下游过程仍不清楚,且羊毛甾醇作为灵芝酸前体外,还用于合成麦角固醇[41]。本研究中测定的灵芝酸生物合成关键酶基因均位于上游过程,且试验组灵芝细胞生长处于旺盛期(图1),推测相关基因表达量增加,产生的羊毛甾醇可能更多的合成用于细胞膜组成成分的麦角固醇。大量研究表明,真菌次级代谢与灵芝发育紧密相连,真菌孢子的分化与次级代谢表现出共调节[42-43],且在ZHANG等[28]的研究中发现,上层菌丝体膜孢子中灵芝酸含量显著高于菌丝,是其20倍。本研究中SAHA处理影响了灵芝菌丝体形态以及抑制孢子生产,这也可能是灵芝酸生物合成降低的原因。在丝状真菌的研究中,全局调控因子蛋白Vet蛋白和LaeA蛋白在调控真菌生长发育和次生代谢中发挥重要作用[44-46]。本研究发现SAHA处理后,和的转录明显下调,这可能是SAHA通过抑制组蛋白去酰化酶活性,改变了组蛋白乙酰化水平导致灵芝染色质重塑和相关基因表达的转变,从而影响了灵芝菌丝的生长,孢子分化和次级代谢。
4 结论
组蛋白去乙酰化酶抑制剂辛二酰苯胺异羟肟酸(SAHA)处理,增加了灵芝组蛋白H4的乙酰化水平,导致灵芝生长延迟且降低了糖耗速率和细胞生长速率,同时降低了灵芝孢子产生。SAHA处理后灵芝气生菌丝形态发生变化,在生长后期抑制了上层白色菌丝体膜色素产生。次级代谢产物灵芝酸的合成与灵芝孢子的产生有较紧密联系,SAHA可能通过影响组蛋白乙酰化水平,进而下调全局性调控因子和的表达,抑制灵芝孢子生成,进而降低灵芝酸的合成;同时影响灵芝多糖途径关键基因表达,进而促进灵芝多糖生物合成。
[1] SHIAO M S, LEE K R, LIN L J, WANG C T. Natural products and biological activities of the Chinese medicinal fungus., 1994, 547: 342-354.
[2] SONE Y, OKUDA R, WADA N, KISHIDA E, MISAKI A. Structures and antitumor activities of the polysaccharides isolated from fruiting body and the growing culture of mycelium of., 1985, 49(9): 2641-2653.
[3] XU J W, ZHAO W, ZHONG J J.Biotechnological production and application of ganoderic acids., 2010, 87(2): 457-466.
[4] 刘高强, 赵艳, 王晓玲, 朱朝阳. 灵芝多糖的生物合成和发酵调控. 菌物学报, 2011, 30(2): 198-205.
LIU G Q, ZHAO Y, WANG X L, ZHU C Y. Biosynthesis and fermentation control of polysaccharides from., 2011, 30(2): 198-205. (in Chinese)
[5] 宁玉波, 王红艳, 乔康, 刘秀梅, 王开运. 灵芝多糖对番茄抗灰霉病的诱导效应. 中国农业科学, 2016, 49(11): 2103-2112.
NING Y B, WANG H Y, QIAO K, LIU X M, WANG K Y. Induced resistance by polysaccharides isolated fromin tomato against gray mold., 2016, 49(11): 2103-2112. (in Chinese)
[6] 张中霄, 王红艳, 王开运, 王东, 姜莉莉. 灵芝多糖拌种对小麦抗纹枯病的诱导效应及生长发育影响. 中国农业科学, 2018, 51(1): 96-104.
ZHANG Z X, WANG H Y, WANG K Y, WANG D, JIANG L L. Induction effect of sharp eyespot of wheat and the effect of wheat growth afterpolysaccharides (GLP) seed dressing., 2018, 51(1): 96-104. (in Chinese)
[7] FANG Q H, ZHONG J J. Two-stage culture process for improved production of ganoderic acid by liquid fermentation of higher fungus., 2002, 18(1): 51-54.
[8] KONG X J, VAN DIEPENINGEN A D, VAN DER LEE T A J, WAALWIJK C, XU J S, XU J, ZHANG H, CHEN W Q, FENG J. Thehistone acetyltransferases are important for morphogenesis, DON biosynthesis, and pathogenicity., 2018, 9: 654. doi: 10.3389/fmicb.2018.00654.
[9] HEDTKE M, RAUSCHER S, ROHRIG J, RODRIGUEZ-ROMERO J, YU Z Z, FISCHER R. Light-dependent gene activation inis strictly dependent on phytochrome and involves the interplay of phytochrome and white collar-regulated histone H3 acetylation., 2015, 97(4): 733-745. doi: 10.1111/mmi.13062.
[10] BRENNA A, GRIMALDI B, FILETICI P, BALLARIO P. Physical association of the WC-1 photoreceptor and the histone acetyltransferase NGF-1 is required for blue light signal transduction in., 2012, 23(19): 3863-3872. doi: 10.1091/mbc.E12-02-0142.
[11] CANOVAS D, MARCOS A T, GACEK A, RAMOS M S, GUTIERREZ G, REYES-DOMINGUEZ Y, STRAUSS J. The histone acetyltransferase GcnE (GCN5) plays a central role in the regulation ofasexual development., 2014, 197(4): 1175-1189. doi: 10.1534/genetics.114.165688.
[12] TRIBUS M, BAUER I, GALEHR J, RIESER G, TROJER P, BROSCH G, LOIDL P, HAAS H, GRAESSLE S. A novel motif in fungal class 1 histone deacetylases is essential for growth and development of., 2010, 21(2): 345-353. doi: 10.1091/mbc.E09-08-0750.
[13] LAN H H, SUN R L, FAN K, YANG K L, ZHANG F, NIE X Y, WANG X N, ZHUANG Z H, WANG S H. Thehistone acetyltransferase AflGcnE regulates morphogenesis, aflatoxin biosynthesis, and pathogenicity., 2016, 7: e74030. doi: 10.3389/fmicb.2016.01324.
[14] ZHANG Q, CHEN L F, YU X, LIU H, AKHBERDI O, PAN J, ZHU X D. A B-type histone acetyltransferase HAT1 regulates secondary metabolism, conidiation, and cell wall integrity in the taxol-producing fungus., 2016, 56(12): 1380-1391.
[15] GóMEZ-RODRíGUEZ E Y, URESTI-RIVERA E E, PATRóN- SOBERANO O A, ISLAS-OSUNA M A, FLORES MARTíNEZ A, RIEGO-RUIZ L, ROSALES-SAAVEDRA M T, CASAS-FLORES S. Histone acetyltransferase TGF-1 regulatessecondary metabolism and mycoparasitism., 2018, 13(4): e0193872. doi: 10.1371/journal.pone.0193872.
[16] TURNER E L, MALO M E, PISCLEVICH M G, DASH M D, DAVIES G F, ARNASON T G, HARKNESS T A A. Theanaphase-promoting complex interacts with multiple histone-modifying enzymes to regulate cell cycle progression., 2010, 9(10): 1418-1431. doi: 10.1128/ec.00097-10.
[17] CAI Q, WANG J J, SHAO W, YING S H, FENG M G. Rtt109-dependent histone H3 K56 acetylation and gene activity are essential for the biological control potential of., 2018, 74(11): 2626-2635. 10.1002/ps.5054. DOI: 10.1002/ps.5054.
[18] BASIMIA T, REZAEE S, ZAMANIZADEH H R, MOUSAVI A. SAHA, histone deacetylase inhibitor causes reduction of aflatoxin production and conidiation in the., 2013, 30(3): 357-366.
[19] YANG K L, ZHUANG Z H, ZHANG F, SONG F Q, ZHONG H, RAN F L, YU S, XU G P, LAN F X, WANG S H. Inhibition of aflatoxin metabolism and growth ofin liquid culture by a DNA methylation inhibitor., 2015, 32(4): 554-563. doi: 10.1080/19440049. 2014.972992.
[20] 邓茂常. 食用菌生产新技术——液体深层发酵. 资源开发与市场, 1987(2): 34-36.
DENG M C.New technology of edible fungi production-submerged liquid fermentation., 1987(2): 34-36. (in Chinese)
[21] 林忠平, 孙安慈, 刘永安. 灵芝深层培养的研究. 微生物学报, 1973, 13(2): 63-68.
LIN Z P, SUN A C, LIU Y A. Studies on the submerged cultivation of Ling Chin()., 1973, 13(2): 63-68. (in Chinese)
[22] 王谦, 李育岳, 杨晓先, 汪麟, 冀宏, 汪虹. 灵芝深层培养及应用研究. 食用菌学报, 1994, 1(2): 41-44.
WANG Q, LI Y Y, YANG X X, WANG L, JI H, WANG H. Submerged culture ofand its application., 1994, 1(2): 41-44. (in Chinese)
[23] LEE S Y, KANG T S, LEE M C. Condition of exo-polysacchride production from submerged mycelial culture ofby using air-lift fermenter system., 1998, 13: 547-553.
[24] 方庆华, 钟建江. 灵芝真菌发酵生产灵芝多糖和灵芝酸. 华东理工大学学报(自然科学版), 2001, 27(3): 254-257.
FANG Q H, ZHONG J J. Simultaneous production of polysaccharide and ganoderic acid by submerged fermentation of., 2001, 27(3): 254-257. (in Chinese)
[25] LIANG C X, LI Y B, XU J W, WANG J L, MIAO X L, TANG Y J, GU T Y, ZHONG J J. Enhanced biosynthetic gene expressions and production of ganoderic acids in static liquid culture ofunder phenobarbital induction., 2010, 86(5): 1367-1374.
[26] XU Y N, ZHONG J J. Impacts of calcium signal transduction on the fermentation production of antitumor ganoderic acids by medicinal mushroom., 2012, 30(6): 1301-1308.
[27] ZHAO W, XU J W, ZHONG J J. Enhanced production of ganoderic acids in static liquid culture ofunder nitrogen- limiting conditions., 2011, 102(17): 8185-8190.
[28] ZHANG W X, ZHONG J J. Effect of oxygen concentration in gas phase on sporulation and individual ganoderic acids accumulation in liquid static culture of., 2010, 109(1): 37-40.
[29] JI S L, LIU R, REN M F, LI H J, XU J W. Enhanced production of polysaccharide through the overexpression of homologous uridine diphosphate glucose pyrophosphorylase gene in a submerged culture of Lingzhi or Reishi Medicinal Mushroom,(Higher Basidiomycetes)., 2015, 17(5): 435-442.
[30] 刘柯, 李焕军, 张德怀, 岳同辉, 李娜, 徐军伟. 灵芝细胞中异源表达透明颤菌血红蛋白基因提高胞外多糖的产量. 食用菌学报, 2017, 24(3): 35-41.
LIU K, LI H J, ZHANG D H, YUE T H, LI N, XU J W. Heterologous expression of vitreoscilla hemoglobin in ganoderma lingzhi for increased exopolysaccharide production., 2017, 24(3): 35-41.(in Chinese)
[31] WILLIAMS R B, HENRIKSON J C, HOOVER A R, LEE A E, CICHEWICZ R H. Epigenetic remodeling of the fungal secondary metabolome., 2008, 6(11): 1895-1897.
[32] FISCH K M, GILLASPY A F, GIPSON M, HENRIKSON J C, HOOVER A R, JACKSON L, NAJAR F Z, W GELE H, CICHEWICZ R H. Chemical induction of silent biosynthetic pathway transcription in., 2009, 36(9): 1199-1213.
[33] TANG Y J, ZHONG J J. Role of oxygen supply in submerged fermentation offor production of Ganoderma polysaccharide and ganoderic acid., 2003, 32: 478-484.
[34] 蓝丽雯. DNA甲基化调控灵芝酸生物合成代谢的研究[D]. 福州: 福建师范大学, 2016.
LAN L W. Study on the regulation of DNA methylation on ganoderic acids biosynthesis in[D]. Fuzhou: Fujian Normal University, 2016. (in Chinese)
[35] 胡彬彬, 林连兵, 魏云林, 季秀玲, 张琦. 一种高效的真菌总蛋白质提取方法. 中国生物工程杂志, 2013, 33(9): 53-58.
HU B B, LIN L B, WEI Y L, JI X L, ZHANG Q. An efficient fungal protein extraction method., 2013, 33(9): 53-58.
[36] ZHANG W X , TANG Y J , ZHONG J J . Impact of oxygen level in gaseous phase on gene transcription and ganoderic acid biosynthesis in liquid static cultures of., 2010, 33(6): 683-690.
[37] LIVAK K J, SCHMITTGEN T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCTmethod., 2001, 25(4): 402-408.
[38] 张虎. 组蛋白去乙酰化酶Rpd3对橘青霉美伐他汀生物合成和生长发育的调控[D]. 重庆: 西南大学, 2017.
ZHANG H. The regulation of histone deacetylase Rpd3 on meiastatin biosynthesis and growth and development in[D]. Chongqing: Southwest University, 2017. (in Chinese)
[39] 叶柳青. 曲古抑囷素A对黄曲霉产毒和生长调控机制的初步研究[D]. 福州: 福建农林大学, 2015.
YE L Q. Regulation of Trichostatin A on toxin production and growth of[D]. Fuzhou: Fujian Agriculture and Forestry University, 2015. (in Chinese)
[40] LEE I, OH J H, SHWAB E K, DAGENAIS T R, ANDES D, KELLER N P. HdaA, a class 2 histone deacetylase of, affects germination and secondary metabolite production., 2009,46(10): 782.
[41] 陈慧, 杨海龙, 刘高强. 灵芝三萜的生物合成和发酵调控. 菌物学报, 2015, 34(1): 1-9.
CHEN H, YANG H L, LIU G Q. Biosynthesis and fermentation control of triterpenoids from Ganoderma lingzhi., 2015, 34(1): 1-9. (in Chinese)
[42] CALVO A M, WILSON R A, WOO B J, KELLER N P. Relationship between secondary metabolism and fungal development., 2002, 66(3): 447-459.
[43] KALE S P, MILDE L T, MARISA K, FRISVAD J C, KELLER N P, BOK J W. Requirement of LaeA for secondary metabolism and sclerotial production in., 2008, 45(10): 1422-1429.
[44] ATOUI A, KASTNER C, LAREY C M, THOKALA R, ETXEBESTE O, ESPESO E A, FISCHER R, CALVO A M. Cross-talk between light and glucose regulation controls toxin production and morphogenesis in., 2010, 47(12): 962-972.
[45] CHETTRI P, CALVO A M, CARY J W, DHINGRA S, GUO Y, MCDOUGAL R L, BRADSHAW R E. Thegene of the pine needle pathogenregulates sporulation and secondary metabolism., 2012, 49(2): 141-151.
[46] BAYRAM O S, BAYRAM O, VALERIUS O, PARK H S, IRNIGER S, GERKE J, NI M, HAN K H, YU J H, BRAUS G H. LaeA control of velvet family regulatory proteins for light-dependent development and fungal cell-type specificity., 2010, 6(12): e1001226.
Effects of Histone Acetylation onGrowth, Polysaccharide and Ganoderic Acid Biosynthesis
ZHANG ZongYuan1, 2, JIANG YongMei1, ZHANG WenXian1
(1College of Life Sciences, Fujian Normal University, Fuzhou 350108;2Institute of Biology Co., Ltd., Henan Academy of Sciences, Zhengzhou 450008)
【Objective】Histone acetylation modification plays an important role in the growth, development and metabolic synthesis of fungi. Few epigenetic studies of higher medicinal fungi were reported at present. In this study, the effects of histone acetylation on the growth and development of() and the synthesis of its main metabolites were studied by adding chemical epigenetic inhibitor octanedianiline hydroxamic acid (SAHA), which provided a theoretical basis for improving the biosynthesis ofpolysaccharides and Ganoderma acid (GA) by epigenetic means.【Method】A two-stage cultivation, liquid fermentation combined with static cultivation, was applied to culture. The cell was treated with different concentration of SAHA (0, 0.6, 60, 120, and 180 μmol∙L-1) during the liquid static cultivation of.Biomass, sugar consumption, mycelial mat formation, mycelial morphology, sporulation and biosynthesis of GA andpolysaccharides were measured or observed by conventional methods.Histone acetylation levels ofwere examined by Western blot, the relative expression levels ofpolysaccharides biosynthesis genes (.and), GA biosynthesis genes (and) andglobal regulator,gene were detected by qRT-PCR. 【Result】The acetylation level of histone H4intreated with SAHA increased to 1.6 times as much as that under control group. SAHA inhibited the growth ofmycelia and the production of pigments, and changed the morphology of mycelia. The formation of spores was also inhibited, and the higher the concentration of SAHA, the more obvious the inhibition degree. SAHA treatment significantly increased the yield ofpolysaccharides, up to 50%, and the biosynthesis of GA was inhibited, which decreased by 13%-27% compared with the control. The results of qRT-PCR analysis showed that the gene expression of the key enzymes inpolysaccharides and GA synthesis were up-regulated in different degrees under SAHA treatment. The gene expression of the key enzymes in polysaccharides synthesis were increased by 1.5-3.5 times and that of the key enzymes in GA synthesis by 1.8-12.1 times. The expression ofandgenes, the global regulators, were inhibited, which was 11.3%-62.4% of the control group.【Conclusion】Histone acetylation could regulate the growth and development ofthrough global regulatory factors, thus affecting the biosynthesis of GA, while histone acetylation also had an effect onpolysaccharides biosynthesis.
; histone acetylation; ganoderic acid;polysaccharide; SAHA
2019-05-21;
2019-10-30
国家自然科学基金(31370099)、福建省自然科学基金(2017J01624)
张宗源,E-mail:xuanyizzy@163.com。通信作者章文贤,E-mail:huzx@fjnu.edu.cn
(责任编辑 赵伶俐)
猜你喜欢
中国生物化学与分子生物学报(2022年8期)2022-09-08
小学生学习指导(低年级)(2022年3期)2022-03-28
复旦学报(自然科学版)(2021年5期)2021-11-17
医药前沿(2020年29期)2020-12-04
山西医科大学学报(2020年8期)2020-09-16
金桥(2019年11期)2020-01-19
当代陕西(2019年5期)2019-03-21
安全(2015年7期)2016-01-19
天津农业科学(2015年10期)2015-10-20
天津农业科学(2015年10期)2015-10-20
