
黑暗下贮藏温度对西瓜幼苗叶片超微结构及光合特性的影响
2020-03-11 08:33段青青黄丹枫贺洪军
核农学报 2020年2期
段青青 丁 明 黄丹枫 贺洪军
(1德州市农业科学研究院,山东 德州 253015;2西北农林科技大学园艺学院,陕西杨凌 712100;3上海交通大学农业与生物学院,上海 200240)
西瓜[Citrullus lanatus(Thunb.)Matsum.and Nakai]是经济效益比较显著的一种园艺作物,2017年,我国西瓜栽培面积约185.97 万hm2,总产量7 948.70万t,分别占世界西瓜栽培面积和总产量的53.5%和67.1%,均居世界第一。西瓜种苗市场需求量的日益增大加强了不同地域间的种苗流通,因此,种苗的及时运输是种苗足量供应的关键。但有时也会出现茬口安排不合理、缺乏劳动力、恶劣天气变化等意外情况,使得种苗不能按期定植或销售,需要进行贮藏,贮藏温度是影响种苗质量的关键因素之一[1-2]。研究表明,在贮藏期间补充光照对种苗质量的保持及定植后的恢复效果优于低温黑暗贮藏[3-6]。但是相对于温度的控制,在种苗贮藏期间安装补光装置对设备的要求更高,安装更复杂。因此,国内种苗运输多在无降温措施的黑暗条件下进行。研究发现,种苗贮藏期间的低温、黑暗环境会对种苗质量产生不同程度的影响,主要表现为植株干物质消耗、叶绿素破坏[7-9]、碳水化合物贮量减少[10-11]、膜脂过氧化水平上升[12-14]等,但贮藏环境对种苗超微结构及定植后光合性能的恢复情况研究相对较少[15-16]。本试验选取黑暗15℃、黑暗25℃两种模拟贮藏条件,通过分析叶片的光合结构,探讨不同贮藏温度对西瓜种苗叶片叶绿体结构及定植后光合性能恢复的影响差异,旨在为西瓜种苗贮运技术及幼苗定植后的恢复提供理论依据。
1 材料与方法
1.1 试验材料
西瓜嫁接苗品种为早佳84-24,砧木为南瓜(Cucurbita moschataDuch.),品种为壮士,均来自上海源怡种苗有限公司。基质配比为珍珠岩∶蛭石∶草炭=2∶2∶1(v/v/v)。砧木比接穗提前5 d 播种,砧木长至一片真叶,接穗两片子叶完全展开时采用插接法进行嫁接。嫁接苗在27℃,相对湿度95%,光强50 μmol·m-2·s-1条件下愈合6 d 后,转入温室内生长,温室内昼夜温度为26/20℃,相对湿度60%~80%,5 d后,选取长势一致的幼苗转移至HP1000GS-D 型植物生长箱(武汉瑞华仪器设备有限责任公司)中进行培养,光周期12 h/12 h,生长箱内设定光强300 μmol·m-2·s-1,昼夜温度25/18±1℃,相对湿度70%。幼苗每天浇灌1/2 浓度的霍格兰氏营养液,待嫁接苗长至两叶一心时,选取均匀一致的幼苗进行贮藏试验。
1.2 试验设计
试验用西瓜苗均为72 孔穴盘苗,在生长箱中进行模拟贮藏试验,生长箱内光强设为0 μmol·m-2·s-1,温度分别设为15℃(storage at 15℃,S15)和25℃(storage at 25℃,S25)两个处理;另有一组穴盘苗继续在正常生长条件(与贮藏前一致)下培养,作为对照(control,CK),每个处理设3 次重复。
试验期间,每次取样前一天,将穴盘从生长箱中拿出,在1/2 浓度的霍格兰氏营养液中浸盘5 min,随后将穴盘随机换位放回生长箱中,所有操作在暗光(<1 μmol·m-2·s-1)下进行。贮藏4 d 和6 d 后,两个处理分别取出144 株幼苗,移栽至塑料盆(直径20 cm)中在正常生长条件下恢复6 d。
取样时间为开始处理后的0、2、4、6 d,以及处理4 d 和6 d 后分别恢复6 d,取第一片真叶进行相关指标的测定。
1.3 测定项目与方法
1.3.1 叶绿体超微结构样品制备 取各处理3 株植物的叶片,避开主脉,切成1 mm×1 mm 的小块,采用戌二醛和锇酸双固定法制备电镜样品[17],利用JEM-2100 型透射电镜(日本JEOL 有限公司)观察叶绿体结构并拍照,计算叶绿体长、宽。
1.3.2 叶绿素荧光参数的测定 取12 株长势均一的幼苗,用便携式OS1-FL 型叶绿素荧光仪(美国OptiScience 公司)测定同一部位叶片的PSⅡ实际光化学效率(actual photochemical,ФPSⅡ)、初始荧光(original fluorescence,Fo)和最大荧光产量(maximal fluoroscence,Fm),计算 PS Ⅱ最大光化学效率(maximum photochemical,Fv/Fm)。
1.3.3 光合参数的测定 测定开始前每个处理选取均一的待测植株先在CK 条件下恢复2 h,然后采用CIRAS-2 型光合仪(英国PP Systems 公司)进行测定。条件设定如下:光强800 μmol·m-2·s-1、CO2浓度375 μmol·mol-1、叶室面积1.7 cm2、叶室温度25℃、大气相对湿度45%±5%;测定项目:净光合速率(net photos ynthetic rate,Pn,μmol·m-2·s-1)、气孔导度(stomatal conductance,Gs,mmol·m-2·s-1)、胞间CO2浓度(intercellular CO2concentration,Ci,μmol·mol-1)和蒸腾速率(transpiration rate,Tr,mol·m-2·s-1)。每处理测定3 株,每张叶片记录10 个值。
1.3.4 地上部干重的测定 选取12 株幼苗,剪取幼苗地上部,放入烘箱105℃杀青10 min 后80℃烘干至恒重,取幼苗的平均值为干重。
1.4 数据统计
用SPSS 16.0 统计软件进行数据计算及处理,Tukey 法检验进行(P<0.05)显著性分析,用Origin 8.5软件绘图。
2 结果与分析
2.1 不同贮藏温度对西瓜幼苗叶绿体超微结构的影响
贮藏前的叶绿体呈长椭圆形,基质和基粒类囊体垛叠丰富,片层沿叶绿体长轴方向排列整齐且紧密,淀粉粒清晰(图1-A)。与贮藏前(0 d)相比,黑暗贮藏2 d,S15 和S25 幼苗叶片叶绿体开始膨大,淀粉粒消失,基粒垛叠减少(图1-B、C);只有靠近叶绿体被膜的地方有片层分布,内缘片层消失,两种处理无明显差异。随着贮藏时间延长,叶绿体逐步膨胀,到贮藏第6 天,两种贮藏温度的叶绿体形状均近似球形(图1-D、E、F、G)。不同的是,S25 幼苗类囊体模糊不清,片层排列产生扭曲现象,且有较多的嗜锇颗粒出现(图1-D、F);而S15 幼苗叶绿体内基粒类囊体垛叠较多,与基质类囊体界限清晰,嗜锇颗粒较少(图1-E、G)。
由图2 可知,与贮藏0 d 相比,黑暗贮藏2 d,两种处理幼苗的叶绿体长、宽和叶绿体长/宽无显著变化。贮藏4 d 时,S15 和S25 幼苗叶绿体长度缩小,宽度增加,叶绿体长/宽降低,且S25 比S15 变化更明显。贮藏6 d 时,S25 和S15 叶绿体长/宽分别比贮藏前(0 d)降低了63.2%和57.5%,这也是黑暗贮藏条件下叶绿体形状变圆的原因。

图1 黑暗下不同贮藏温度对西瓜幼苗叶片叶绿体超微结构的影响Fig.1 Effect of different different storage temperature in darkness on chloroplast ultrastructure of watermelon seedlings leaves
2.2 不同贮藏温度对西瓜幼苗叶绿素荧光参数的影响
由图3 可知,随着黑暗贮藏时间的延长,西瓜幼苗叶片的Fv/Fm和ФPSⅡ呈下降趋势,贮藏4 d 内,S25 的Fv/Fm和ФPSⅡ下降缓慢,且与S15 之间无显著差异;贮藏6 d 时,S25 的Fv/Fm和ФPSⅡ急速下降,且显著低于S15,与贮藏0 d 相比,S25 的Fv/Fm和ФPSⅡ分别降低了24.72%和23.04%,而S15 的Fv/Fm和ФPSⅡ分别降低了7.35%和13.82%。表明贮藏温度对短期黑暗贮藏的幼苗光化学活性无显著影响,但随着贮藏时间的延长,低温能相对延缓黑暗贮藏造成的幼苗Fv/Fm和ФPSⅡ的降低。
2.3 不同贮藏温度对西瓜幼苗光合特性的影响
黑暗贮藏降低了西瓜幼苗叶片的Pn、Gs 和Tr,贮藏时间越长,降低越明显(图4)。贮藏4 d 内,两种处理幼苗叶片的Gs 和Ci 均降低,可知黑暗贮藏降低了气孔的开度,从而引起了Pn 的降低;贮藏6 d 时,Gs继续下降,Ci 却显著上升(P<0.05),气孔开度的降低并未引起Ci 的下降,但Pn 持续降低,推测是非气孔因素导致的结果,这可能与黑暗贮藏6 d 叶绿体结构被破坏有关。此外,从贮藏温度来看,贮藏4 d 时,S25 幼苗叶片的Pn、Gs、Ci 和Tr 与S15 均无显著差异;贮藏6 d 时,S15 幼苗叶片的Pn、Gs 和Tr 均显著高于S25,但二者之间的Ci 无显著差异,这可能与S15 幼苗叶绿体内片层结构保存相对良好有关。
2.4 不同贮藏温度对西瓜幼苗叶片超微结构恢复的影响
黑暗贮藏4 d 的幼苗定植后6 d,其细胞壁及质膜结构清晰可见,叶绿体恢复到椭圆形,类囊体排列紧密且有序,片层结构清晰,基粒沿叶绿体长轴排列,淀粉粒在叶绿体内重新积累,且S15 和S25 两处理恢复后的叶绿体结构无明显差异(图5-A、C)。从贮藏温度来看,S25 处理贮藏6 d 的幼苗叶绿体结构在定植后6 d 仍不能恢复正常,叶绿体形状膨大近似半球形,叶绿体内基粒较少,排列相对紊乱,且叶绿体内出现较多的嗜锇颗粒(图5-B)。S15 处理贮藏6 d 的幼苗定植后6 d,其叶绿体内含有较少的嗜锇颗粒(图5-D),叶绿体的形状与正常叶绿体(图1-A)无明显差异,叶绿体内的片层结构虽排列疏松,但基粒类囊体与基质类囊体界限清晰,类囊体排列正常有序。
2.5 不同贮藏温度对西瓜幼苗光合及荧光恢复的影响
黑暗贮藏不仅影响西瓜幼苗的叶绿体结构,还会导致PSⅡ光化学活性和气体交换参数下降,且贮藏时间的长短影响定植后西瓜幼苗生长及光合特性的恢复。由图6 可知,贮藏4 d 的幼苗恢复6 d,S15 和S25两处理幼苗叶片的Fv/Fm和ФPSⅡ均能恢复到CK 水平;贮藏6 d 的幼苗恢复6 d,S15 幼苗叶片的Fv/Fm和ФPSⅡ能恢复到CK 水平,但S25 显著低于CK。黑暗贮藏对Pn 的影响与对PSⅡ的影响不同,S25 处理4 d 和6 d 的幼苗在定植后6 d 其叶片Pn 均不能恢复到CK水平;而S15 处理4 d,其Pn 在定植后6 d 可恢复,S15处理6 d,Pn 在定植后6 d 虽不可恢复,却显著高于S25,可见低温贮藏提高了幼苗定植后的光合恢复能力。幼苗植株干物质重的变化能反映其体内物质积累和消耗的情况,黑暗贮藏能影响西瓜幼苗定植后的生长,S15 和S25 两处理黑暗贮藏4 d 和6 d 的幼苗,地上部干重在6 d 内都不能恢复到CK 水平,但S15 幼苗地上部干重高于S25。

图2 不同贮藏温度对叶绿体长、宽及叶绿体长/宽的影响Fig.2 Effect of storage temperature on the chloroplast length,chloroplast width and the ratio of chloroplast length/width of watermelon seedlings leaves

图3 黑暗下不同贮藏温度对西瓜幼苗叶片Fv/Fm 和ФPSⅡ的影响Fig.3 Effect of storage temperature on the Fv/Fm and ФPSⅡof watermelon seedlings leaves
3 讨论
3.1 黑暗下贮藏温度对西瓜幼苗叶绿体结构、叶绿素荧光及光合特性的影响
黑暗是诱发植物衰老的因子之一。Niu 等[18]研究发现,野生型拟南芥离体叶片黑暗处理4 d,叶绿体内基质片层排列松散,有较多嗜锇颗粒出现。Hilda-Araceli 等[19]发现小麦叶片经黑暗诱导4 d,叶绿体膨胀呈圆形,类囊体排列松散,质体小球变大且增多;黑暗诱导6 d 后,叶绿体消失。本试验结果显示,黑暗贮藏2 d,西瓜幼苗叶片叶绿体内淀粉粒消失,基粒垛叠减少,内缘片层消失;黑暗贮藏4 d,叶绿体开始膨大;黑暗贮藏6 d,叶绿体内嗜锇颗粒增多,叶绿体膨胀成近似球形。研究表明,叶绿体的大小、数量、形态及类囊体的垛叠程度都会随温度的改变发生变化[20],且叶绿体对温度的响应因处理时间和植物种类不同有所差异。李国强等[21]研究表明,12℃和9℃夜间低温处理7 d 的番茄叶片叶绿体结构与对照(15℃)无显著差异,但6℃低温处理7 d 的叶绿体基粒数减少,片层排列松散。而枇杷小苗在-2℃低温处理时叶绿体内的类囊体才有垛叠程度降低的现象出现[22]。芳樟一年生幼苗在-10℃胁迫24 h,叶绿体严重损伤,而猴樟的叶绿体结构尚能维持完整[23]。本试验中,黑暗贮藏4 d,S25 与S15 的叶绿体片层结构才出现差异;黑暗贮藏6 d,S25 的叶绿体片层排列、扭曲,嗜锇颗粒较多,而S15 的叶绿体内基粒较多,与基质类囊体界限清晰,嗜锇颗粒较少。可见,随着贮藏时间延长,低温可相对减轻黑暗对西瓜幼苗叶绿体结构的破坏,进而贡献于植物的光合性能。
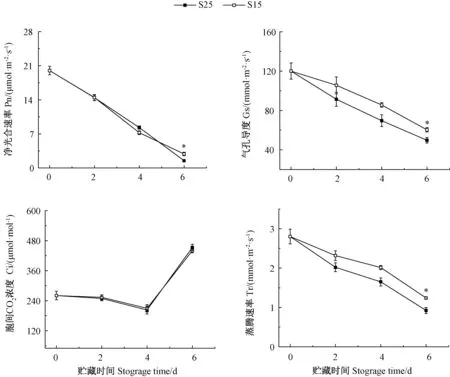
图4 不同贮藏温度对西瓜幼苗叶片光合作用的影响Fig.4 Effect of storage temperature on photosynthetic parameter of watermelon seedlings leaves
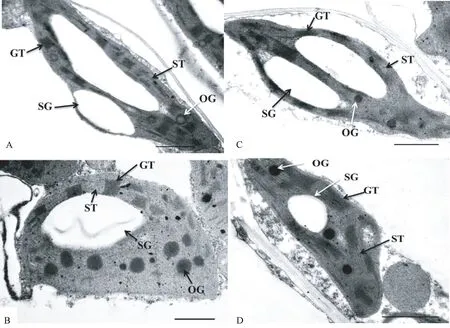
图5 不同贮藏温度对西瓜幼苗叶片叶绿体超微结构恢复的影响Fig.5 Effect of storage temperature on the recovery of chloroplast ultrastructure from the watermelon seedlings leaves

图6 不同温度黑暗贮藏对西瓜幼苗Fv/Fm、ΦPSⅡ、Pn 和地上部干重恢复的影响Fig.6 Effect of storage temperature on the recovery of Fv/Fm,ΦPSⅡ,Pn and shoot dry weight of watermelon seedlings during storage in darkness
叶绿体作为植物细胞内进行光合作用的细胞器,对衰老的响应最敏感[24],其衰老往往导致光合系统的衰退[25-27]以及PSⅡ光化学效率的降低[28-30]。本试验结果显示,黑暗贮藏造成叶绿体结构破坏的同时,也伴随着Fv/Fm、和ФPSⅡ的下降。黑暗贮藏4 d 内,S15和S25 二者的Fv/Fm和ФPSⅡ无显著差异;黑暗贮藏6 d 时,S15 的Fv/Fm和ФPSⅡ显著高于S25,说明低温能减缓长期黑暗贮藏对幼苗PSⅡ化学反应中心活性的损伤。
光合作用的下降主要有气孔限制和非气孔限制两种原因,Gs 和Ci 同时下降,才可说明是气孔限制因素导致的光合速率降低[31],且有研究表明,植物遭受胁迫加重时,Pn 与Ci 呈现相反的变化趋势[32]。本试验结果显示,黑暗贮藏引起西瓜幼苗光合速率的下降是由气孔限制和非气孔限制两种因素共同作用的,黑暗贮藏4 d,气孔限制是光合速率降低的主要因素;黑暗贮藏6 d,S15 和S25 的幼苗叶绿体均膨大接近球形,类囊体片层均受损,此时是非气孔限制占主导作用,这与前人对矮牵牛的研究结果相符[33]。此外,从贮藏温度来看,黑暗贮藏4 d 内,S25 幼苗叶片的Pn、Gs、Ci 和Tr 与S15 无显著差异;贮藏6 d,处理之间的Ci 值无显著差异,但是S15 幼苗叶片的Gs、Tr 和Pn 均显著高于S25,这正好与S15 幼苗保存有相对良好的叶绿体片层结构的现象相吻合。可见短期黑暗(4 d 内),贮藏温度对西瓜幼苗的叶绿体结构、光化学活性和光合作用的影响差异不明显,而低温对长期黑暗(6 d)贮藏引起的叶绿体结构、PSⅡ反应中心及光合作用的破坏能起到减缓作用。
3.2 黑暗下贮藏温度对定植后西瓜幼苗叶绿体结构、叶绿素荧光及光合特性恢复的影响
Macijej 等[34]研究发现,黑暗低温对叶绿体造成的破坏短时间还可以恢复,长时间不可恢复。本试验结果显示,黑暗贮藏4 d 的西瓜幼苗恢复光照6 d,其叶绿体结构和形状均可恢复。但是,黑暗贮藏6 d,S25的叶绿体结构在定植后6 d 仍不能恢复正常。相比较而言,S15 处理的幼苗定植后6 d,叶绿体内含有较少的嗜锇颗粒,叶绿体的形状与正常叶绿体无明显差异。此外,黑暗下,两种温度处理的西瓜幼苗叶绿体淀粉粒均消失,因为幼苗在黑暗中无法进行光合作用储存能量,呼吸作用的持续进行使原本储存在叶绿体中作为能量储备的淀粉粒不断被消耗、分解和利用,用于植株的生长代谢,因此淀粉粒的消失是对黑暗逆境的一种适应;恢复光照后,叶绿体内淀粉粒重新积累,说明叶绿体作为光合作用场所的功能开始恢复。
叶绿体结构的优劣与PSⅡ反应中心的光化学能力密切相关,叶绿体结构良好,PSⅡ的光化学活性也相对稳定[35]。本试验结果显示,定植后6 d,黑暗贮藏4 d 的西瓜幼苗叶片Fv/Fm和ФPSⅡ均恢复到CK 水平;黑暗贮藏6 d 的幼苗恢复情况因贮藏温度不同有所差异,S15 幼苗叶片的Fv/Fm和ФPSⅡ可以恢复到CK 水平,S25 则不能,这与其叶绿体结构的不完全恢复有关。可见,短期黑暗贮藏下,适度低温使PSⅡ中心发生可逆性失活,并未造成PSⅡ反应中心的永久性破坏,但25℃黑暗贮藏6 d 的幼苗PSⅡ反应中心是遭受了永久性破坏还是需要更长的时间恢复还有待研究。
研究认为,光合作用的降低与叶绿体结构的紊乱有关[36]。本研究结果显示,贮藏温度对黑暗贮藏后幼苗光合能力的恢复影响较大,西瓜幼苗定植后6 d,S25处理4 d 和6 d 的幼苗Pn 均不能恢复到CK 水平,S15处理4 d 的幼苗Pn 能够恢复,而S15 处理6 d 的幼苗Pn 虽然未恢复到CK 水平,但显著高于S25,可见适度低温提高了种苗贮藏后的光合恢复能力。种苗贮藏期间较高的光合能力有助于幼苗定植后的生长[37],虽然在幼苗定植后6 d,黑暗贮藏4 d 和6 d 的幼苗地上部干重均显著低于CK,但S25 地上部干重显著高于S15,可见低温能提高黑暗贮藏的植株定植后的生长速度。
4 结论
西瓜种苗经黑暗贮藏均导致叶片叶绿体结构破坏、PSⅡ反应中心活性下降、光合速率降低,进而影响贮藏幼苗定植后的恢复速度。黑暗条件下,与常温(25℃)贮藏相比,低温(15℃)贮藏有助于维持幼苗质量的原因在于当贮藏时间适当延长时能较好地保持叶绿体的结构,具有较高的PSⅡ光化学活性和碳同化能力,从而使幼苗定植后的光合性能和物质积累快速恢复。建议西瓜种苗黑暗常温下贮藏不宜超过4 d,黑暗低温下贮藏不宜超过6 d。本研究结果可为西、甜瓜等瓜类种苗贮藏提供一定的理论参考。但由于本试验只分析了两种贮藏温度下幼苗的光合生理变化,关于不同低温或施加外源物质等对幼苗的物质代谢、信号传导、激素变化等方面的影响还有待进一步研究。
猜你喜欢
园艺与种苗(2022年3期)2022-05-13
园艺与种苗(2022年2期)2022-04-09
园艺与种苗(2021年11期)2021-12-30
军事文摘(2021年16期)2021-11-05
园艺与种苗(2021年5期)2021-06-30
今日农业(2020年23期)2020-12-15
课程教育研究·学法教法研究(2019年18期)2019-10-08
少儿科技(2019年9期)2019-09-10
劳动保护(2018年8期)2018-09-12
山东工业技术(2016年15期)2016-12-01
