
绿豆产量性状的QTL定位
2020-03-13 11:55陈吉宝
中国农业科技导报 2020年10期
陈吉宝
(南阳师范学院, 河南省南水北调中线水源区生态安全重点实验室, 河南 南阳 473061)
绿豆由于富含优质蛋白质和具有清热解毒等功效,是我国人民喜爱的医食两用食用豆类。长期以来,由于单产低,和大宗作物的比较效益低,导致绿豆种植面积逐年减少,严重影响了我国绿豆种植业的发展[1]。目前,我国绿豆平均单产为1 500 kg·hm-2,虽然高于世界平均水平(721 kg·hm-2),但和世界高单产(2 250 kg·hm-2)相比仍有很大差距[2],因此,培育高产绿豆品种仍然是我国绿豆育种的重要目标之一。密度、单株荚数、单荚粒数和百粒重是豆科作物产量形成的4要素,而株高和单株分枝数通过影响株型而影响种植密度,也是决定豆科作物产量的重要农艺性状[3-4]。王兰芬等[5]发现,正常水分条件下绿豆单株产量与单荚粒数呈极显著正相关(0.476);刘长友等[6]研究表明,单株产量与株高呈极显著正相关(0.368);Banger等[7]研究也显示,产量与株高、单荚粒数、单株荚数的相关系数分别为0.40*、0.39*、0.39*。因此,培育高产绿豆新品种,需要综合考虑这些性状的影响[4]。
作物产量性状是受多基因控制的数量性状,QTL(quantitative trait locus)定位是挖掘数量性状基因、开发分子标记辅助育种的有效方法。目前,应用QTL技术已在大豆、菜豆、鹰嘴豆等豆科作物中定位出大量产量性状基因,提高了育种的准确性[8-10]。在绿豆中,利用QTL挖掘出抗绿豆象、抗白粉病等数量性状基因[11],也有研究者对绿豆产量相关性状进行了QTL分析。Fatokun等[12]利用F2群体将百粒重基因定位在1、2、3、6连锁群上;梅丽等[13]利用RIL群体,分离出多个决定株高、单株分枝数、单株粒数、百粒重等性状的基因;Kajonphol等[14]利用QTL在第1连锁群发现2个单荚粒数基因、在第7、8连锁群各发现1个粒长基因位点。尽管对绿豆产量性状基因定位已进行有效的探索,但都仅限于个别性状,尚未对构成绿豆产量性状进行统一研究,特别是对百粒重有重要决定意义的籽粒特性(粒长、粒宽、籽粒直径、籽粒周长)研究更少。
“VC2917”绿豆是来自亚洲蔬菜研究与发展中心(AVRDC)的直立型地方品种,具有粒大、分枝数高、苗期抗旱特点;“鹦哥绿豆”是来自张家口市的蔓生型地方品种,具有单产高、分枝数少、成株期抗旱性特点[6]。Liu等[15]以“VC2917”为母本,“鹦哥绿”为父本,杂交后通过单粒传方法构建了包含256个家系的“VC2917/鹦哥绿”RIL群体,并用312个SSR标记构建了遗传连锁图谱,成功定位出干旱胁迫决定下绿豆株高、最大叶面积、生物量、叶片相对含水量和产量性状的 50个QTL位点,证明该群体适合用于绿豆数量性状基因定位。本研究以“VC2917/鹦哥绿”RIL群体为材料,在自然条件下通过3年的田间表型评价,研究绿豆产量性状的遗传特征,并利用Liu等[15]构建的遗传连锁图谱对产量性状进行QTL定位,旨在尽可能发掘有利用价值的等位基因,为绿豆高产品种选育提供理论依据。
1 材料与方法
1.1 试验材料
绿豆RIL群体由中国农业科学院作物科学研究所食用豆课题组提供。
1.2 试验方法
1.2.1田间试验设计与管理 大田试验于2017—2019年春季在河南省南阳市潦河镇农田进行,于每年4月下旬播种,完熟后收获。播种前2周浇透水,播前1周深翻耕土壤一次,翻耕深度为30 cm,并随机耕施复合肥375 kg·hm-2。单行播种,每行10穴,每穴播种3粒,行距0.6 m,株距0.4 m,于出苗后一周定苗,每穴留1株。绿豆生长期间按照正常田间管理措施,于分枝形成前中耕一次,并起垄培土,整个生长期间不再浇水。试验采用完全随机区组设计,重复3次。
粒长(seed length, SL)、粒宽(seed width, SW)、籽粒直径(seed diameter, SD)、籽粒周长(seed perimeter, SP)用SC-G谷物籽粒考种仪(杭州万深)测量,每个材料取50粒种子进行扫描,读取数据。粒长是指平行于种脐纵线、自籽粒基部至顶端的距离;粒宽是指自种脐纵线中点至籽粒种脐正对面的直线长度;籽粒周长是指起始于种脐纵线中点、切面垂直于种脐纵线、围绕绿豆种子一周的长度;籽粒直径是指起始于种脐纵线中点、切面垂直于种脐纵线、环绕籽粒的圆的直径。
1.2.3产量性状的遗传分析 首先,对不同年份12个性状的表型数据进行方差同质性Bartlett检验,根据检验结果计算每份材料12个性状的最佳线性无偏估计值(BLUE)。
式中,Wj为第j个重复的权重,Yj为第j个重复下性状的均值,e为总的重复数。
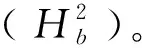
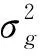
1.2.4产量相状的QTL分析 以12个性状的最佳线性无偏估计值对产量相关性状进行QTL分析,定位采用QTLIciMappingV4.0软件改进复合区间作图法[17],Permutation次数设置为1 000次,LOD≥3.0为标准。
2 结果与分析
2.1 绿豆产量性状遗传分析
2.1.1绿豆产量性状统计分析 从表1可以看出,12个性状的表型BLUE在RIL群体内均呈极显著或显著差异;RIL群体内各性状的最大值和最小值均分别高于亲本最大值和小于亲本最小值,群体内单株荚数和单株产量的均值显著高于双亲均值(P<0.05),其他性状的均值与双亲均值差异不显著。性状变异系数为 4.9%~36.1%,其中变异系数最小的是粒宽,最大的是单株产量,以上结果说明不同性状在RIL群体个体之间均存在较大程度的分离。12个性状的遗传力都较大,在57.6%~96.6%之间,其中单株分枝数、单荚粒数、荚长、荚宽、籽粒直径、籽粒周长和百粒重的遗传力均大于90%,说明这几个性状的遗传稳定性高,在早代选择有利于获得稳定高产材料。
(3)思维的逻辑性不强。有的学生在地理学习中虽然知道了某一问题的地理知识,但思维条理不清,在组织语言或者答题中思维混乱,不够严谨,结果可能是错误的结论。从学生近几年的答题考试中发现学生缺乏地理逻辑思维。学生答题过程中要么是基础知识不牢固,缺乏答题角度或者是罗列知识点,逻辑性差;要么是学生的空间方向不清,地理事物的空间方位颠倒,没有清晰的地理区位空间;或者答题不能用简练的专业术语表达,答题语言不规范等,都是因为学生的思维不活跃,逻辑思维能力差的表现,这就导致的高考考分无法提高。
2.1.2绿豆产量性状群体分布 从表1可以看出,除株高、单株分枝数、单株荚数、单株产量和荚长的偏度大于0.5外,其他性状的偏度绝对值小于0.5;单荚粒数、荚宽、粒宽、籽粒直径、籽粒周长的偏度小于0,表型数据分布右偏,其他性状偏度大于0,表型数据分布左偏,其中荚长的偏度为2.421,呈明显左偏分布。株高、单荚粒数、荚宽、粒长、籽粒直径、籽粒周长峰度绝对值均小于0.5,其他性状的峰度绝对值均大于0.5;粒长和籽粒周长的峰度小于0,表明这两个性状表型分布不太集中,而荚长和单株荚数的峰度大于1,特别是荚长的峰度达到10以上,说明这两个性状的表型存在极端值。从图1可以看出,RIL群体各性状值均呈连续变化,都出现超亲现象,说明这12个性状均属于数量性状,受多基因控制或基因互作的影响。
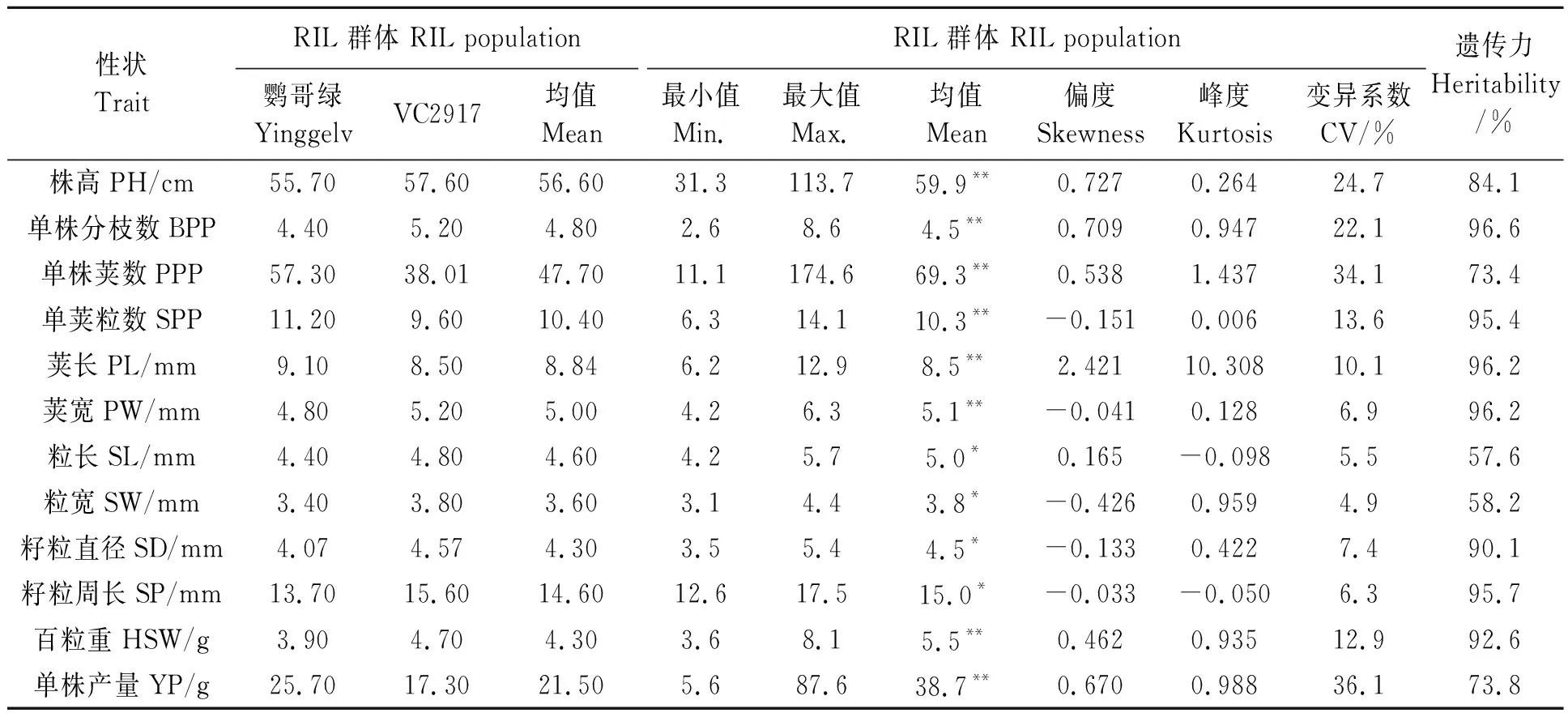
表1 重组自交系群体各产量相关性状的分布特征Table 1 Distribution characteristics of yield traits in RIL population
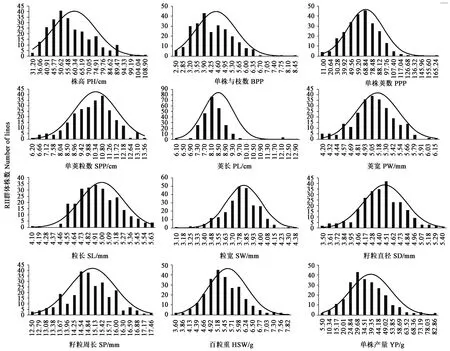
图1 绿豆产量相关性状频率分布Fig.1 Frequency distribution of mung bean yield traits
2.2 绿豆产量性状相关性分析
对12个性状进行相关性分析,结果如表2所示。单株产量与单株荚数、单荚粒数、粒长、籽粒周长、百粒重呈显著正相关,其中,与单株荚数相关性最强(0.837),表明单株荚数在决定绿豆产量中起关键作用,其次为百粒重(0.294);单株产量与荚长相关系数为-0.072,暗示其可能对单株产量的形成起负作用。
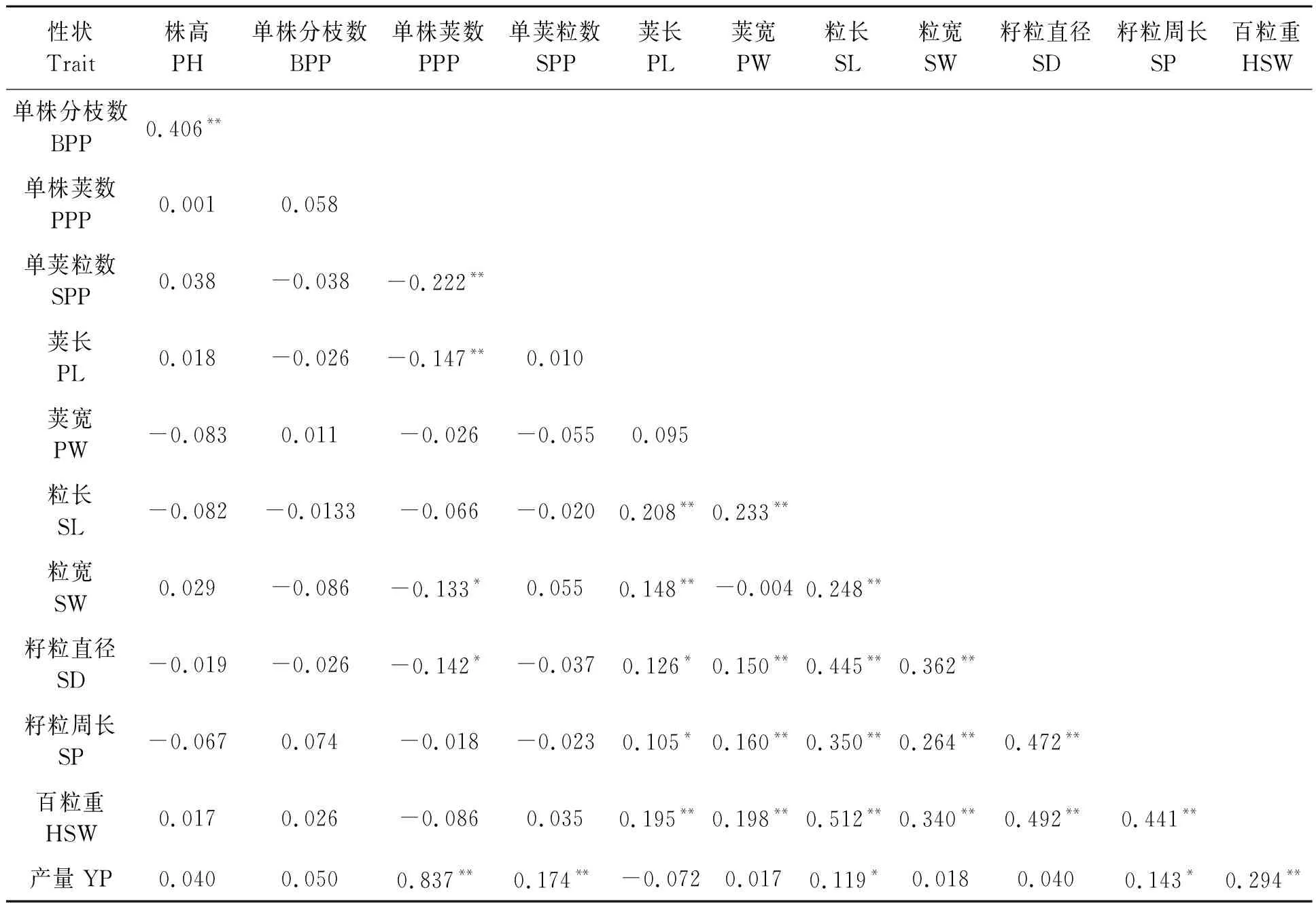
表2 绿豆产量相关性状间的相关性分析Table 2 Correlation analysis of mung bean yield traits
百粒重与4个籽粒性状(粒长、粒宽、籽粒周长、籽粒直径)和2个荚性状(荚长与荚宽)均呈显著正相关,其中籽粒性状与百粒重的相关系数显著大于2个荚性状,说明籽粒性状对百粒重起决定性作用;百粒重与株高、单株荚数呈不显著正相关。
另外,株高与单株分枝数之间呈极显著正相关(0.406),而这两个性状与其他10个性状之间呈不显著正或负相关,说明株高通过单株荚数对产量的形成起关键作用。单株荚数与除单株产量、株高、单株分枝数外的9个性状均呈负相关,其中与单荚粒数、荚长、粒宽和籽粒直径之间呈极显著负相关,特别是与单荚粒数之间的负相关最强(-0.222),说明绿豆产量形成中,单株荚数和单株粒数呈互斥关系。
2.3 绿豆产量性状QTLs定位分析
2.3.1绿豆产量相关性状连锁群分布 对12个产量相关性状进行QTL分析,共检测到56个QTLs位点,其中2017、2019年各检测到20个QTLs,2018年检测到16个QTLs。56个QTLs分布在绿豆除8号外的10个连锁群上(图2)。
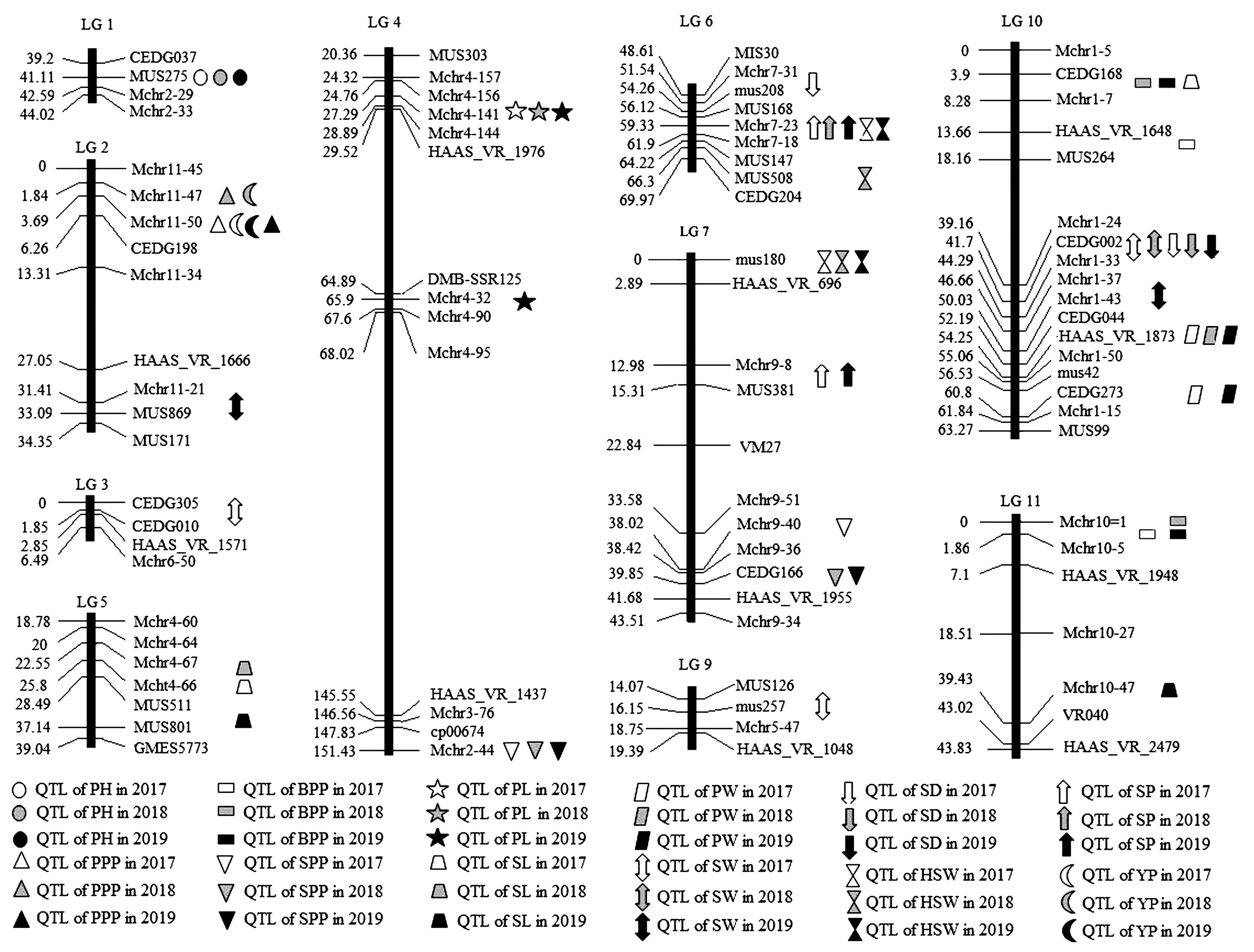
图2 绿豆产量相关性状的QTLs在连锁群上的分布Fig.2 Distribution of QTLs for yield traits on mung bean genetic map
2.3.2株高和单株分枝数QTLs 3个年份均在第1连锁群的CEDG037~MUS275标记之间检测到株高(PH) QTL(PH1a、PH1b、PH1c),这三个位点完全重合,说明可能由一个基因控制;单个位点在贡献率分别为9.48%、14.52%和15.42%,遗传效应较大。三个QTL位点加性效应均为负值,平均加性效应为-6.85,表明该位点的基因累加效应可以使株高减少6.85 cm(表3)。3个年份均在第10和11连锁群上各检测到 1 个单株分枝数(BPP)QTL位点,其中位于第11连锁群的QTL位点(BPP11a、BPP11b、BPP11c)完全重合在Mchr10-1~Mchr10-5区间,暗示在该区间可能存在1个控制单株分枝数的基因;2018和2019位于第10连锁群的QTL位点(BPP10b、BPP10c)均在CEDG168~Mchr1-7区间,两个位点相距1 cM,可能由不同基因控制;2017年位于第10连锁群的QTL位点(BPP10a)位于HAAS_VR_1648~MUS264标记之间,距BPP10b位点10 cM;6个BPP位点的加性效应均为负值(表3)。
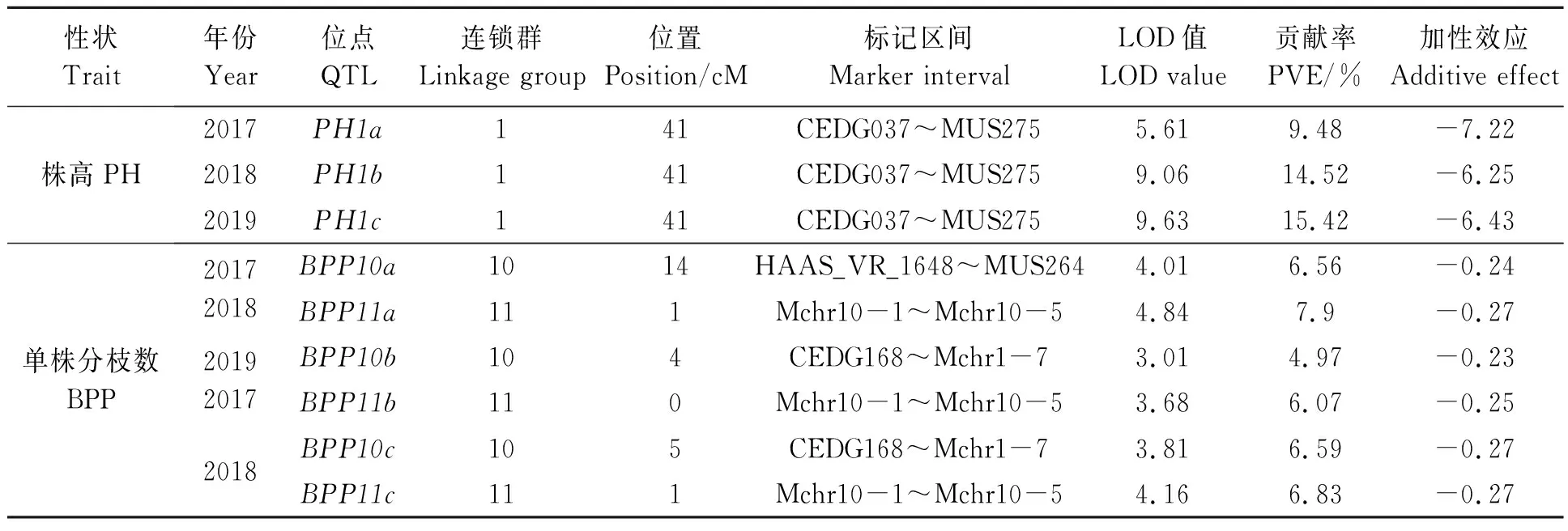
表3 株高和单株分枝数QTLsTable 3 QTLs for mung bean plant high (PH) and number of branches per plant (BPP)
2.3.3果荚性状QTL 2017、2018和2019年各在第1连锁群均检测到单株荚数(PPP)QTL位点(PPP1a、PPP1b、PPP1c),其中PPP1a和PPP1c重合于Mchr11-50~CEDG198之间,与PPP1b的标记区间(Mchr11-47~CEDG198)相邻,单个位点的表型贡献率均超过10%,属于大效应位点,表明该位点对单株荚数有重要贡献,存在决定绿豆产量的关键基因。三个位点的加性效应均为正,平均加性效应为11.11,表明该区间的基因可使单株荚数增加11.11个(表4)。
3个年份均在第4连锁群的Cp00674~Mchr2-44区间检测到1个单荚粒数(SPP)QTL位点(SPP4a、SPP4b、SPP4c),单个位点贡献率较高最小为14.81%,最高达17.18%,表明该位点对SPP有较大贡献,是决定SPP的大效应位点。在第7连锁群的CEDG198~Mchr11-34区间也检测到SPP的QTL位点(SPP7a、SPP7b、SPP7c),平均贡献率比第4连锁群QTL位点少10.23%,说明该位点对SPP的贡献不大,可能属于微效基因。第4连锁群的QTL加性效应均为负,可平均使SPP减少0.58粒;第7连锁群加性效应为正,可平均使SPP增加0.34粒(表4)。
共检测到4个荚长(PL)QTLs,均位于第4连锁群;其中2017、2018位点(PL4a、PL4b)重合于Mchr4-156~Mchr4-141标记之间,贡献率分别为6.49%、5.47%;2019检测到位点PL4c.1也位于Mchr4-156~Mchr4-141标记之间,但是距离PL4a为1 cM,贡献率较大(8.72%),PL4c.2所在标记区间(Mchr4-32~Mchr4-90)与PL4c.1的标记区间相距较远(表4)。以上结果说明,控制PL的基因可能存在3个,其中PL4a(PL4b)和PL4c.1的加性效应为正,PL4c.2加性效应为负。
共检测到 5 个荚宽(PW)QTLs,分别分布于第10连锁群CEDG044~HAAS_VR_1873区间(PW10a.1、PW10b、PW10c.1)和CEDG273~Mchr1-15区间(PW10a.2和PW10c.2);前者的平均贡献率为16.94%,属于大效应位点,后者的平均贡献率为9.64%;前者加性效应均为正,后者加性效应为负,表明前一个位点对荚宽的增长有促进作用,后一个位点对荚宽的增长起减效作用(表4)。
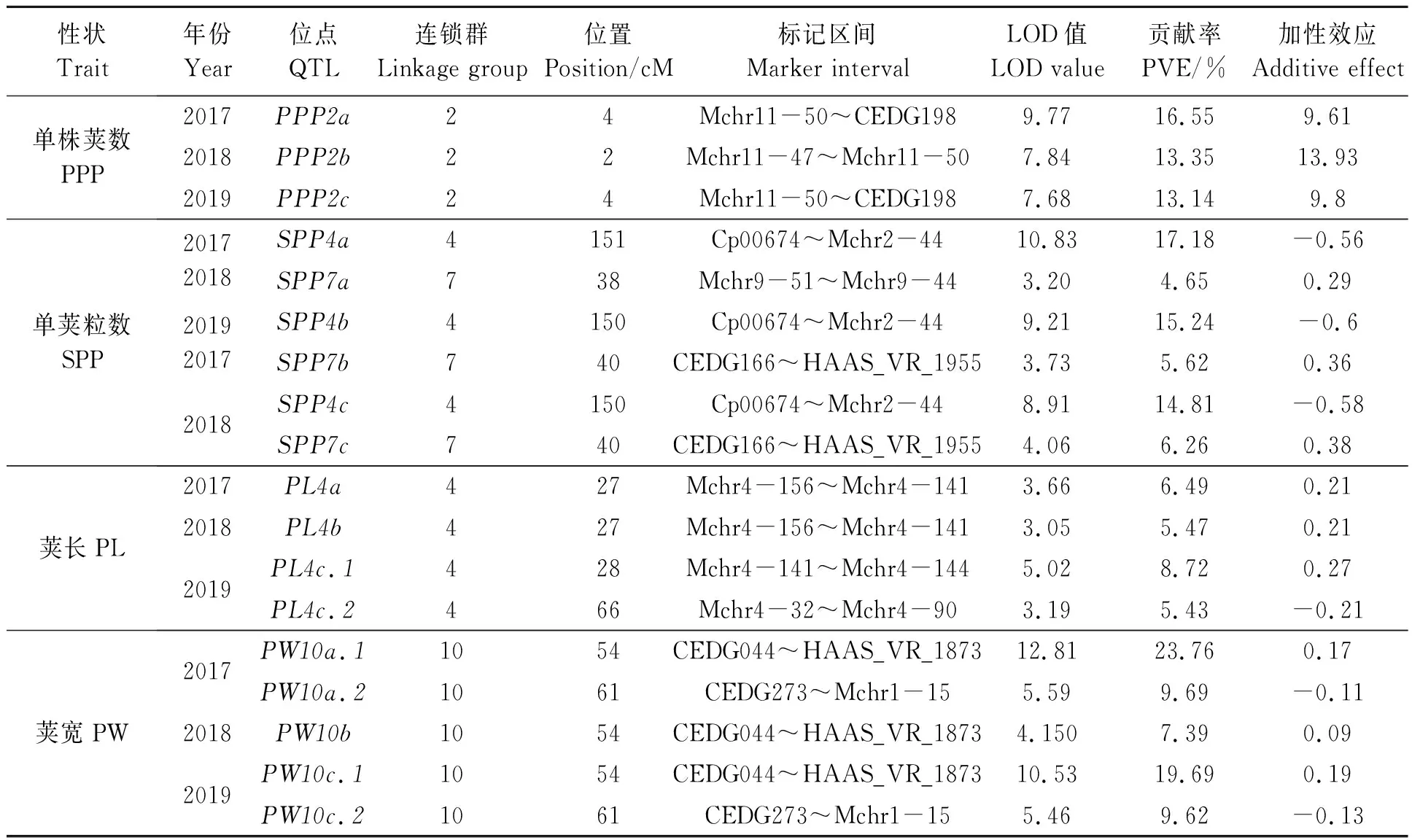
表4 绿豆果荚性状QTLsTable 4 QTLs for mung bean pod characters
2.3.4籽粒性状QTL 3个年份,分别在第5连锁群检测到3个粒长(SL) QTL位点(SL5a、SL5b、SL5c),在第10连锁群检测到1个(SL10a),在第11连锁群检测到1个(SL11c)。其中,位于第5连锁群的3个QTL遗传距离距离较远,可能不是由同一基因控制(表5)。5个位点的总贡献率为40%,单个贡献率最大为10.08%,最小为5.96%,加性效应均为正,表明决定SL的单个基因对SL的贡献不大,但综合贡献较大。
共检测到6个粒宽(SW)QTL,位于第2、3、9、10连锁群上,其中第10联锁群分布3个位点,这3个位点相距较远,表明这5个位点分别存在决定SW的基因(表5)。平均贡献率为8.13%,其中SW10a位点贡献率为16.19%,是决定SW大效应位点,具有重大的育种利用价值。
共检测到4个控制籽粒直径(SD)的QTL位点(SD6a、SD10a、SD10b、SD10c),其中位于第10联锁群的三个QTL位点均在CEDG013~VR304标记之间,平均贡献率较大,加性效应均为正值,说明该区间存在对SD具有较大增效作用的基因(表5)。
共检测到5个籽粒周长(SP)QTLs(SP6a、SP7a、SP6b、SP6c、SP7c),SP6a、SP6b、SP6c重合于第6连锁群的MUS168~Mchr7-23标记之间,单个位点贡献率最大23.39%、最小为16.66%、平均贡献率为20.13%;SP7a、SP7c重合于第7连锁群的Mchr9-8~MUS381标记之间,贡献率远小于SP6a、SP6b、SP6c;5个SP QTL位点的加性效应均为负,对籽粒周长表现减效作用(表5)。
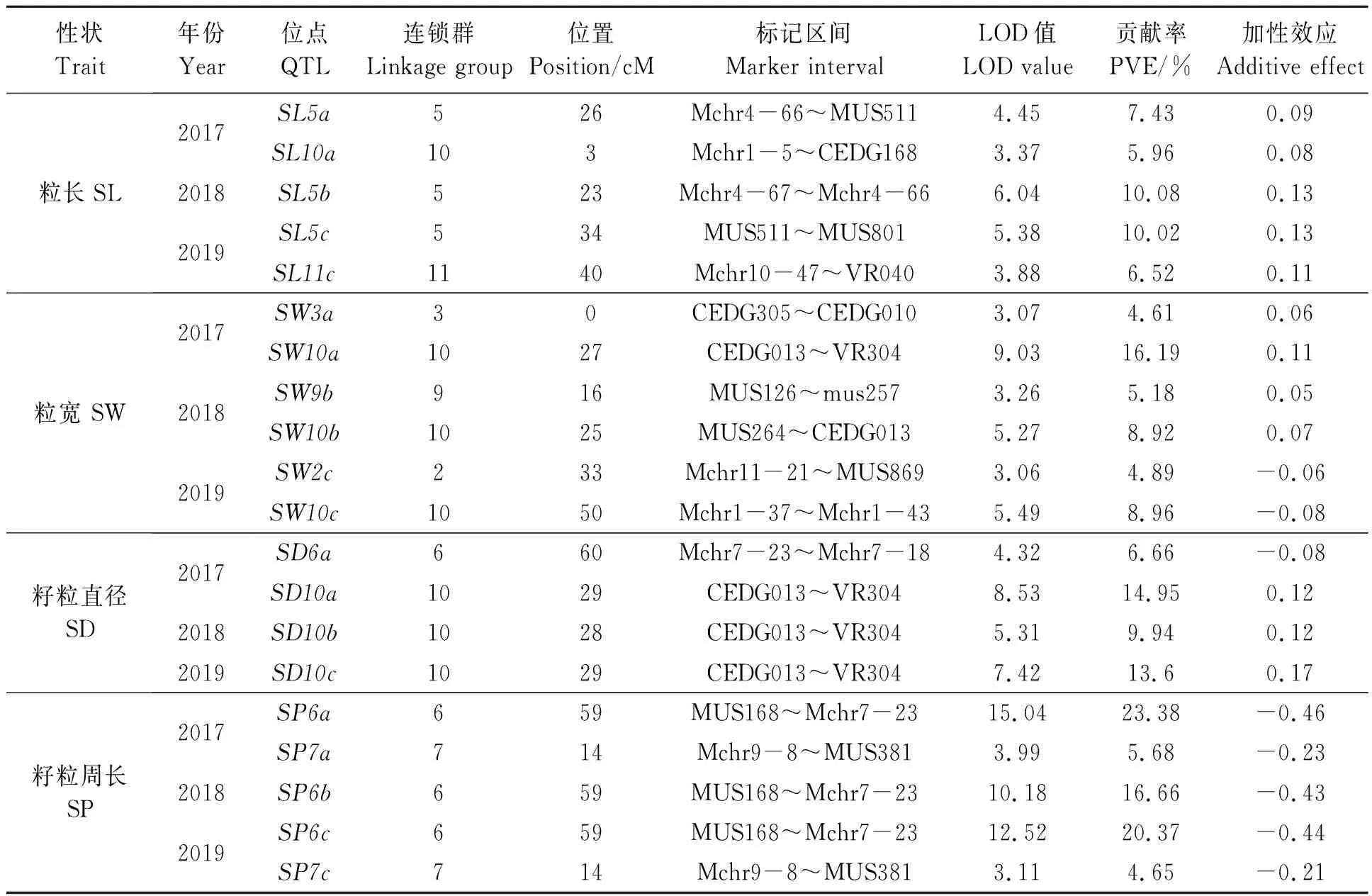
表5 绿豆籽粒性状QTLsTable 5 QTLs for mung bean seed characters
2.3.5百粒重和单株产量QTL 3个年份均在第6和第7连锁群上各检测到1个百粒重(HSW)QTL。HSW6a和HSW6c虽然重合于Mchr7-23~Mchr7-18标记之间,但相距1 cM;HSW6b位于MUS147~MUS508标记之间,距离HSW6a和HSW6c分别为5和6 cM;HSW6a、HSW6b和HSW6c的贡献率分别为8.38%、8.55%和7.65%,加性效应均为负。HSW7a、HSW7b和HSW7c均位于第7连锁群的0 cM处,其标记区间为mus180~HAAS-VR-696,加性效应也均为负,该位点使百粒重平均减少0.18 g(表6)。
每个年份均在第2连锁群的Mchr11-47~CEDG198区间检测到1个单株产量(YP)QTL(YP2a、YP2b、YP2c),贡献率分别为11.71%、8.41%、7.38%;YP2a和YP2c位点完全重合,距YP2b为2 cM;三个位点加性效应均为正,其中YP2b加性效应最大(6.99)(表6)。
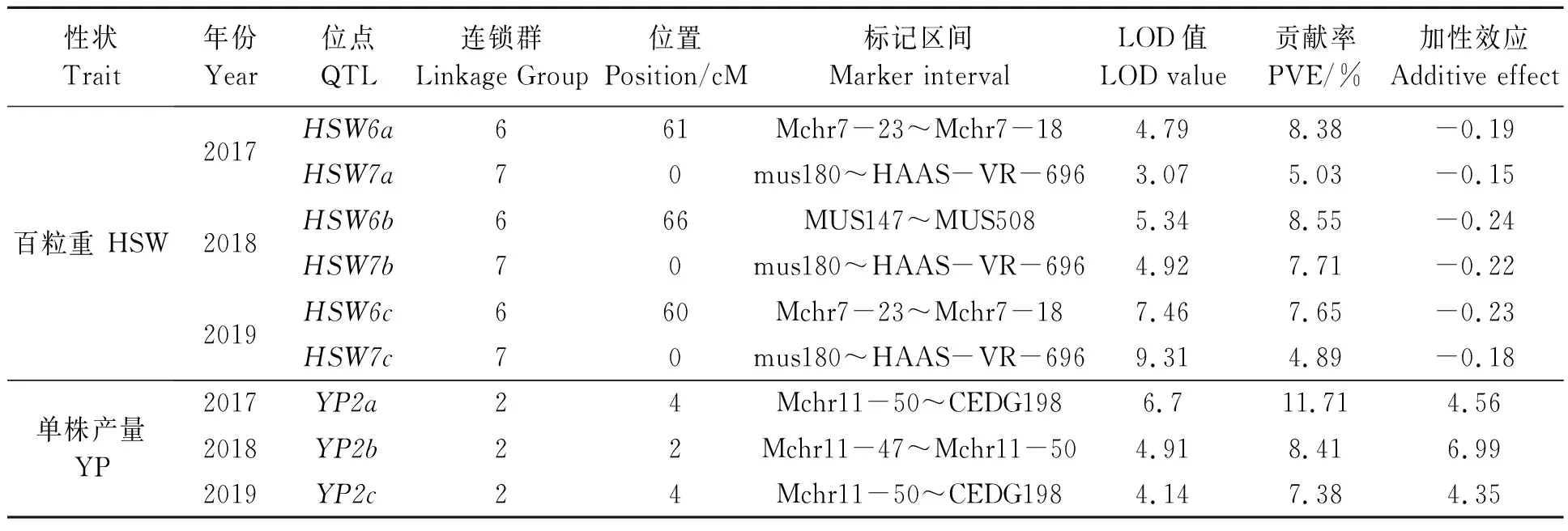
表6 绿豆产量性状的QTLsTable 6 QTLs for mung bean yield traits
3 讨论
豆类作物株高、单株分枝数、单株荚数、单荚粒数和百粒质量是衡量产量的核心要素,它们通过影响单株产量性状影响单位面积产量[18]。本研究中,株高、单株分枝数、单株荚数、单荚粒数和荚长的变异系数均大于10%,最高达34.1%,而籽粒性状变异系数均小于10%,说明农艺性状的遗传受环境影响大于籽粒性状,这与刘兴叶等[19]和方路斌等[20]的研究结果相似。其中,单株产量的变异系数最大(36.1%),其次为单株荚数(34.1%),郑海泽等[21]也发现,绿豆单株产量和单株荚数的变异系数高达50%以上。同时,在本研究中也发现,单株产量与籽粒周长、粒长、百粒重、单荚粒数和单株荚数呈显著正相关,其中与单株荚数相关性最强(0.837),其次为百粒重(0.294)。刘兴叶等[19]结果显示,绿豆产量与单荚粒数和荚长呈极显著正相关;郑海泽等[21]发现,单株产量与单株荚数和荚长呈极显著正相关关系;王官等[22]研究表明,与单株产量的关联度由高到低排序为单株荚数、株高、主茎节数、百粒重、单荚粒数、主茎分枝、生育期。以上结果说明,单株产量、单株荚数等产量相关性状变异丰富,在早代不易稳定,但在早代可选择出单个性状表现突出的材料;籽粒性状的变异系数最小,早期世代稳定下来的可能性较大,适宜早代选择[23];在所分析的几个形状中,单株荚数和百粒重在绿豆单位面积产量形成中扮演重要角色,育种实践中关注单株荚数和百粒重状对选育高产绿豆新品种更有效。
数量性状一般由多个微效基因控制,性状的表型容易受环境影响,在不同群体和不同环境条件下定位到QTL位置可能不一致,在不同环境条件下都能检测到的QTL位点被称为稳定QTL[24]。稳定QTL位点一般是大效应QTL,在这些位点存在相关基因的可能性更大,是利用QTL技术挖掘数量性状基因的关键区域[18]。本研究通过一点3年大田实验发现,12个性状都存在稳定QTL位点,其中株高、单株分枝数、百粒重、荚宽、籽粒直径和籽粒周长各有一个QTL位点在3个年份都在相同位置检测到,单株分枝数、荚长、荚宽、单荚粒数、籽粒周长和单株产量各有1个QTL在2个年份在相同位置检测到(表3~6)。同时,本研究将百粒重基因锁定在第6连锁群上,这与前人的结果类似[25-28],说明6号染色体上存在决定百粒重性状的关键基因;本研究在4号染色体上定位到荚长和单荚粒数基因、在11号染色体上定位到单株分枝数基因,这与梅丽等[13]的定位结果类似。这些稳定QTL说明,本研究定位的QTL具有较大育种利用价值,可作为今后利用分子标记辅助选择培育高产绿豆新品种或克隆相关基因优先考虑位点。特别是在第2连锁群的Mchr11-50~CEDG198标记之间和第4连锁群的Cp00674~Mchr2-44标记之间均在2个年份分别检测到大效应单株荚数QTL和单荚粒数QTL,而单株荚数和单荚粒数与单株产量性状密切相关(表2),因此,这两个区间是未来分子标记辅助选育高产绿豆新品种和克隆产量性状基因的关键区域。
QTL簇(QTL cluster)或QTL富集区是一因多效或者多个基因紧密连锁的结果[29]。本研究共发现3个QTL富集区,分别是第2 连锁群的Mchr11-47~CEDG198区间富集了单株分枝数和单株产量性状QTL(QTL之间的距离为0 cM)、第6连锁群的mus208~MUS147区间富集了百粒重和籽粒周长性状QTL、在第10连锁群的CEDG 002~ HAAS-VR-1873区间富集了荚宽、粒宽和籽粒直径QTL(图2)。以上结果说明,在这些区域可能存在同时决定绿豆单株分枝数和单株产量、百粒重和籽粒周长、籽粒性状的通用性基因,若对这些区域进一步精细定位有可能分别发掘出决定这些性状键基因,这些QTL富集区是未来绿豆产量性状基因图位克隆优先考虑的区域。
猜你喜欢
烟草科技(2022年9期)2022-09-24
安徽农学通报(2022年6期)2022-04-07
作物学报(2022年3期)2022-01-22
山西农业科学(2021年4期)2021-04-19
今日农业(2020年16期)2020-12-14
农民致富之友(2020年27期)2020-09-12
作文评点报·低幼版(2020年18期)2020-05-28
种子(2020年4期)2020-05-20
江苏农业科学(2020年22期)2020-03-03
广西农学报(2019年1期)2019-07-18
