
靶向cGAS-STING信号通路药物的研究进展
2020-07-23 06:02梅家豪
中国药科大学学报 2020年3期
梅家豪,洪 泽,王 琛
(中国药科大学生命科学与技术学院中国药科大学天然药物活性组分与药效国家重点实验室,南京210009)
固有免疫系统是人体免疫系统的第一道防线,可以迅速识别外源病原体入侵,并且诱导巨噬细胞和自然杀伤细胞对病原体进行吞噬和降解,同时释放大量干扰素和炎症因子,促进树突状细胞对抗原的递呈作用,进而激活适应性免疫,形成长期的免疫记忆[1]。在固有免疫系统中,机体能通过细胞上存在的模式识别受体(pattern recognition receptors,PRRs),比如Toll样受体(Toll-like receptors,TLRs),RIG-I 样 受 体(RIG-I-like receptors,RLRs),NOD 样受体(NOD-like receptors,NLRs),DNA 感受器等,特异性识别微生物表达的病原相关的分子模式(pathogen-associated molecular patterns,PAMPs),比如脂多糖(lipopolysaccharide,LPS),单链核糖核苷酸(single strand ribonucleic acid,ssRNA),双链脱氧核糖核酸(double strand deoxyribonucleic acid,dsDNA)等,调控下游细胞因子的表达和释放[1]。
环化核苷酸合成酶(cyclic GMP-AMP synthase,cGAS)就是一种位于细胞质中的dsDNA 识别受体,和下游主要调节蛋白干扰素基因刺激因子(stimulator of interferon genes,STING)介导了细胞质DNA 引起的固有免疫反应。DNA-cGASSTING 信号通路的激活促进细胞内Ⅰ型干扰素(interferons,IFNs)与肿瘤坏死因子α(tumor necrosis factor,TNF-α)和白介素6(interleukin 6,IL-6)等炎症因子的表达和分泌[2]。越来越多的研究表明,cGAS-STING 信号通路在病原体感染、肿瘤和自身免疫疾病等疾病中充当重要角色。本文将对近年来cGAS-STING 信号通路的最新研究进展和靶向cGAS 和STING 蛋白的激动剂和抑制剂开发进行综述,为后续研究提供理论参考。
1 cGAS-STING 信号通路
人源cGAS 由522 个氨基酸组成,相对分子质量大小为60 kD,位于N 末端的130-155 aa 为非保守序列,功能研究尚不明确,而155-522 aa 构成了cGAS 的C 末端核酸转移酶结构域,含有一个位于中心的催化区域和分散在两侧的阳离子表面区域[3]。cGAS 通过上述区域与双链DNA 螺旋中的脱氧核糖磷酸骨架以2∶2 的形式结合成多聚体。此外,cGAS 上的锌指区域通过离子键进一步稳定其与DNA 的结合[4]。DNA 的结合能诱导cGAS 核酸转移酶催化区域的结构重排,进而以ATP 和GTP 为底物催化形成2′-5′和3′-5′环状GAMP 分子(cyclic GMP-AMP),即2′,3′-cGAMP(图1-A)[3]。值得注意的是cGAS 对DNA 的识别并不依赖于DNA 序列的特异性,长链DNA(>45 bp)能更高效地激活cGAS 酶活,且病原体入侵、线粒体受损或基因组不稳定性导致的细胞质中DNA 聚集都能导致cGAS激活[5]。
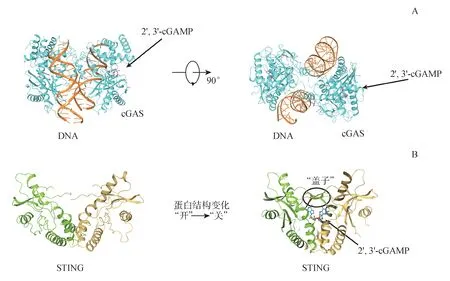
图1 环化核苷酸合成酶(cGAS)和干扰素基因刺激因子(STING)的蛋白结构
2′,3′-cGAMP 在与下游受体蛋白STING 结合后完成信号的传递[6]。人源STING 蛋白由379 个氨基酸组成,包含N 端4 次跨膜区(N terminal transmembrane domain,NTD,1-137 aa),环化核苷酸结合区域(cyclic dinucleotides binding domain,CBD,138-340 aa)和C 末端尾巴(C terminal tail,CTT,341-379 aa)[7]。静息状态下STING 蛋白以同源二聚体形式存在,两个分子的NTD 相互交错将蛋白锚定在内质网膜内,CBD 区域在细胞质中形成“V”形 口 袋 用 于 识 别2′,3′-cGAMP。2′,3′-cGAMP 的结合诱导STING 蛋白的空间构象由“V形”向“U”形口袋转变,且口袋顶部的开口处形成帽子结构促进STING 二聚体形成四聚体和多聚复合物(图1-B),多聚化的STING蛋白脱离内质网向高尔基体转移并在此过程中以其CTT 区域招募TANK 结合激酶(TANK-binding kinase,TBK1),TBK1 在与STING 多聚体结合后发生自磷酸化(172 位丝氨酸发生磷酸化,S172),激活其激酶活性后催化STING蛋白的磷酸化(366位丝氨酸发生磷酸化,S366),磷酸化后的STING 蛋白进一步以其CTT 区域招募转录因子干扰素调节因子3(interferon regulatory factors 3,IRF3),促进IRF3 磷酸化(396 位丝氨酸发生磷酸化,S396)和二聚化入核,激活Ⅰ型IFNs 相关基因的表达[8-9]。同时STING 将信号传给肿瘤坏死因子受体相关蛋白6(TNF receptor associated factor 6,TRAF6),激活经典核因子κB(nuclear factor kappa-B,NF-κB)信号通路,诱导TNF-α 和IL-6 等炎症因子释放[2]。与2′,3′-cGAMP 类似,细菌的代谢产物环鸟苷二磷酸(cyclic diguanylate monophosphate,c-di-GMP)和环腺苷二磷酸(cyclic diadenylate monophosphate,c-di-AMP)分子也能结合到STING 的CBD区域,诱导信号通路的激活[10]。cGAS-STING 信号通路的信号传递过程见图2。

图2 cGAS-STING信号通路示意图
2 cGAS-STING 信号通路紊乱与疾病
在机体的抗病毒、抗细菌方面,cGAS-STING信号通路发挥着重要作用。但是细菌和病毒已经进化出一系列逃逸机制,抑制了固有免疫信号通路的激活。Su 等[11]发现,2 型人类免疫缺陷病毒(human immunodeficiency virus-2,HIV-2)的毒力蛋白X(viral protein X,Vpx)可以和STING 结合,选择性抑制cGAS-STING 信号通路介导的下游经典NF-κB 信号通路,减少TNF-α 和IL-6 等炎症因子释放。Aguirre 等[12]发现登革热病毒的非结构蛋白2B(non-structural protein 2B,NS2B)可以靶向并引起cGAS 溶酶体降解,使其不能识别感染过程中泄露的线粒体DNA,抑制了Ⅰ型IFNs 表达和释放。正是因为cGAS-STING 信号通路在抗病原微生物中可以发挥重要作用,所以开发靶向STING的免疫佐剂已逐渐成为研究热点。
cGAS-STING 信号通路通过促进Ⅰ型IFNs,TNF-α和IL-6等炎症因子表达,一方面能诱导细胞产生大量干扰素刺激基因(interferon-stimulated genes,ISGs),而另一方面能促进树突细胞向T 细胞呈递抗原,加强抗肿瘤免疫[13]。但是肿瘤细胞能从多个层面抑制cGAS-STING 信号通路从而逃避免疫监察。通过生物信息学研究发现,在肾癌、乳腺癌、肺癌、肝癌等多种人肿瘤细胞系中,cGAS和STING 的基因通过表观遗传学降低了翻译效率,而且在蛋白水平通常是以不能行使正常功能的突变体形式存在[14]。而且Wu 等[15]发现肿瘤细胞中人上皮生长因子受体2(human epidermal growth factor receptor 2,HER2)可以招募RAC-β丝氨酸/苏氨酸蛋白激酶(RAC-beta serine/threonine-protein kinase,AKT),抑制STING 信号通路,促进了肿瘤发展。同时也有研究发现如果NF-κB信号通路过度激活,炎症反应会参与肿瘤的发生、转移、免疫逃逸以及肿瘤微环境中的血管生成,促进肿瘤的恶化[16]。比如致癌物二羟甲基丁酸(dimethylolbutanoic acid[2,2-bis(hydroxymethyl)butyric acid],DMBA)会导致表皮角质细胞核DNA 发生损伤,并使其释放到细胞质,从而激活细胞内的cGAS-STING 信号,促进皮肤的癌变[17]。还有在肺癌细胞的脑转移模型中,在转移到脑部的肺癌细胞中,细胞质中的DNA 可以激活cGAS,而生成的2′,3′-cGAMP能通过Gap Junction向大脑部的星形胶质细胞转移,进一步激活cGAS-STING 信号通路,诱导Ⅰ型IFNs 和TNF-α 等炎症因子表达和释放,而释放的炎症因子会通过旁分泌的方式,作用于肿瘤细胞上,促进了肿瘤细胞的生长[18]。综上,cGAS-STING 信号通路在肿瘤发生、发展和转移过程中发挥重要作用,而且在大部分的肿瘤模型中,在细胞和动物水平已经证实cGAS-STING信号通路能够发挥很好的抗肿瘤免疫作用,但是不同肿瘤中STING 激动剂的安全性和有效性需要谨慎评价。
机体内STING 信号通路的持续异常激活能导致多种自身免疫疾病。第一类是因基因突变导致细胞内核酸代谢酶功能异常,比如TREX1,RNASEH2A,SAMHD1等基因发生突变,促进DNA 在细胞质中的聚集,诱导炎症细胞因子的持续释放,导致自身免疫疾病发生[19-21]。此类疾病包括AGS 综合征(Aicardi-Goutieres syndrome,AGS)、系统性红 斑 狼 疮(systemic lupus erythematosus,SLE)等[22]。另一类是由编码STING 蛋白发生V147L,N154S 和V155M 等点突变(第147 位缬氨酸突变成亮氨酸,第154位天冬酰胺突变成丝氨酸,第155位缬氨酸突变成甲硫氨酸),导致STING 蛋白不依赖上游2′,3′-cGAMP即可持续激活,促进炎症因子持续表达和释放,导致组织损伤[23]。在临床上发现此类患者早发于婴儿时期,主要表现为皮肤红疹和呼吸急促,发热,外周血管病变,肺部炎症和血液中存在自身抗核抗体等,此类疾病也被统一命名为诱发于婴儿时期的STING 相关血管病变(STING-associated vasculopathy with onset in infancy,SAVI)。
除了上述疾病外,近年来人们发现越来越多的代谢与炎症疾病与cGAS-STING 信号通路调控密切相关。在急性胰腺炎模型中,坏死组织泄露的外源DNA 会诱导胰腺中的巨噬细胞分泌大量炎症因子,造成组织损伤,而这一现象在cGAS 和STING 基因敲除小鼠中均有改善[24]。在DNASE2双等位基因突变的患者中,研究人员发现Ⅰ型IFNs 过度表达,有核红细胞数量增加,同样在cGAS 和STING 基因敲除小鼠中疾病症状得到缓减,也说明了由cGAS-STING 介导的Ⅰ型IFNs分泌可能参与了血液生成[25]。在帕金森病小鼠模型中,人们发现由于缺少E3 泛素连接酶parkin(E3 ubiquitin-protein ligase parkin,PRKN)和蛋白激酶PINK1(serine/threonine-protein kinase PINK1),受损线粒体中渗露的DNA 可能诱导炎症因子释放,而在小鼠模型中将STING 蛋白进行基因敲除,可以减慢疾病发展[26]。目前已与cGAS-STING 信号通路相关的疾病见表1。
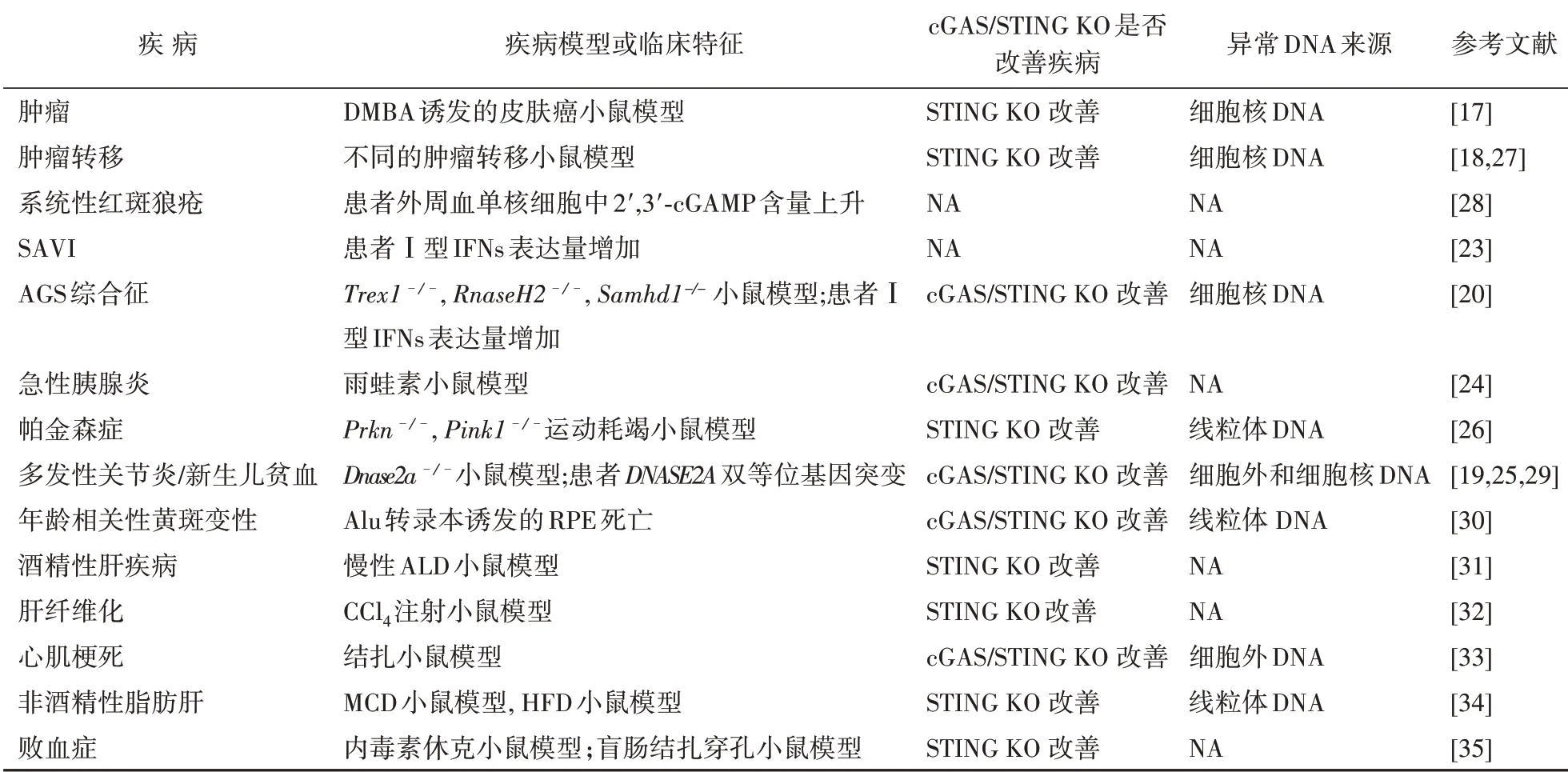
表1 与cGAS-STING信号通路相关的疾病
3 靶向cGAS-STING 信号通路的药物研发
鉴于cGAS-STING 信号通路在疾病的发生发展中发挥的重要作用,而且一些免疫佐剂如铝盐、明矾和壳聚糖等能够通过诱导细胞死亡或线粒体损伤导致细胞质内的DNA 聚集进而激活cGASSTING 信号通路,促进机体的免疫功能[36-37],所以靶向cGAS 和STING 蛋白的药物开发也逐渐成为研究热点。
3.1 cGAS抑制剂
目前,直接靶向cGAS 的激动剂还没有报道,虽然一些金属离子(例如锰离子、锌离子等)可以增加cGAS 的酶活,但其所需的有效浓度过高且易在体内产生毒性不利于后续药物研发[38]。因此,有关抑制剂的研究较多。Vincent等[39]通过建立体外cGAS 酶活检测方法,高通量筛选出的化合物RU.521 能高效且特异性地抑制cGAS 酶活。在小鼠单核巨噬细胞(RAW 264.7)的体外模型中RU.521 对下游Ⅰ型IFNs 表达的半数抑制率(IC50)为700 nmol/L。共结晶结构表明,RU. 521 通过占据cGAS 的催化位点影响2′,3′-cGAMP 合成,从而发挥抑制作用。进一步的研究表明RU. 521 对鼠源cGAS 酶活性的抑制要显著高于人源cGAS 蛋白,所以该研究团队用新的实验方法筛选出了高效抑制人源cGAS 酶活性的先导化合物G140 和G150,且在人原代巨噬细胞体外模型中,G150 表现出良好的生物学活性(IC50为600 nmol/L)[40]。此外,Hall 等[41]以荧光法检测2′,3′-cGAMP的产生为依据进行高通量筛选,筛选出的化合物PF-06928215与人源cGAS 蛋白的亲和力为0.2 μmol/L,对外源cGAS 酶活的半数抑制率为4.9 μmol/L。但是在细胞实验中,PF-06928215并不能抑制DNA 诱导细胞内IFN-β表达,这可能与化合物细胞透膜能力和与血浆蛋白结合能力有关。
除了全新发现的小分子抑制剂,一些已经上市的药物也被发现有类似的功能。An 等[42]通过cGAS 和DNA 的结构数据,进行计算机模拟筛选,发现了一系列抗疟疾药物,比如喹吖因(quinacrine,QC)、羟 氯 喹(hydroxychloroquine,HCQ)等,可以阻碍cGAS 和DNA 相互结合,抑制Ⅰ型IFNs 表达。但是在后续实验中发现,这些化合物可能会插入DNA 中,并在10 μmol/L 浓度下就会出现明显细胞死亡,所以抗疟疾药物用于抑制Ⅰ型IFNs 的安全性还有待考虑。在QC 和HCQ 的基础上,该研究团队优化一系列抗疟疾药物衍生物并筛选出具有高效生物活性和细胞安全性化合物X6。实验证明,化合物X6 在Trex1-/-小鼠的骨髓来源的巨噬细胞(bone marrow-derived macrophage,BMDM)和SLE 患者的外周血单核细胞(peripheral blood mononuclear cell,PBMC)中能显著下调ISGs表达[43]。由于没有具体的结构学和药理学数据,利用抗疟疾药物用于治疗自身免疫性疾病的具体机制,有待进一步实验研究。Liu 等[44]发现,茶多酚中的表没食子儿茶素没食子酸酯(epigallocatechin gallate,EGCG)可以通过抑制GTP 酶激活蛋白1(GTPase-activating protein SH3 domain-binding protein 1,G3BP1)蛋白活性,间接抑制cGAS 和DNA结合。同一团队的Dai等[45]发现,乙酰水杨酸可以直接乙酰化cGAS,抑制下游Ⅰ型IFNs 生成,并且在Trex1-/-小鼠体内和AGS 患者来源的成纤维细胞中都有明显的抑制炎症效果。
综上,现有针对cGAS 酶活区域的结构或者cGAS/DNA 复合物的结构进行设计筛选的化合物具有较弱的生物活性,所以建立起一个具有高通量、高灵敏度的检测平台,获得更加全面的蛋白质结构数据,将极大促进靶向cGAS 的小分子研究进程。而且,越来越多的数据证明,通过影响蛋白质翻译后修饰过程,可以有效调控蛋白质介导的信号转导,所以开发靶向蛋白翻译后修饰的化合物也是未来的研究方向。
3.2 STING激动剂
由于STING 信号通路的激活促进下游Ⅰ型IFNs 的表达在抗病毒免疫和抗肿瘤免疫中发挥着重要的作用,所以靶向STING 蛋白激动剂开发一直是该领域的研究热点。STING 激动剂的研究主要集中在环化核苷酸(cyclic dinucleotides,CDNs)类似物的优化和新型小分子激动剂的筛选。CDNs最早在细菌中发现,是由腺嘌呤核糖核苷酸(adenosine monophosphate,AMP)和鸟嘌呤核糖核苷酸(guanosine monophosphate,GMP)通过经典磷酸二脂键连接形成的环化二鸟嘌呤c-di-GMP 和环化二腺嘌呤c-di-AMP 或者cGAMP。由cGAS 催化生成的2′,3′-cGAMP 是 哺 乳 动 物 体 内 唯 一 已 知 的CDNs。相对于3′,3′-cGAMP 和其他的CDNs,2′,3′-cGAMP 和STING 具有更强的亲和力,在解离常数(Kd)方面,比c-di-GMP,3′,2′-cGAMP 和3′,3′-cGAMP 低99%,比2′,2′-cGAMP 低98%[46]。2′,3′-cGAMP 以非共价的形式结合到STING 二聚体形成的口袋内,诱导STING 蛋白的变构激活。天然存在的环化核苷酸化学结构式见图3。

图3 天然环化核苷酸化学结构式
CDNs 可以在肿瘤微环境中促进CD8+T 细胞活化,产生持续的杀伤肿瘤效果,是一类十分具有临床潜力的抗肿瘤化合物[47-48]。目前已经有多种CDNs 进入临床前或临床研究(表2)。有关STING激动剂的药物化学性质和生物学效应在其他综述 中已详细讨论,在这里不再赘述[49]。

表2 处于临床前和临床研究阶段的新颖STING激动剂
3.3 STING抑制剂
近年来研究发现STING 的异常激活导致的免疫功能紊乱能诱发AGS、SLE 和SAVI 等自身免疫疾病,这也使得研究者开始重视STING 抑制剂的开发。Merck 公司的Siu 等[53]通过ALIS(automated ligand identification system,ALIS)系统筛选出了靶向STING 蛋白的小分子抑制剂Compound 18。共结晶结构表明Compound 18 结合在STING 的CBD口袋内,并竞争性抑制2′,3′-cGAMP与蛋白的结合。在人单核细胞(THP-1)内,Compound 18抑制2′,3′-cGAMP 诱导细胞IFN-β基因表达的IC50为11 μmol/L。此外在植物环肽化合物库中筛选发现紫菀根茎的天然提取物Astin C 能通过阻止STING对下游转录因子IRF3 的招募,抑制多种刺激物诱导的STING 信号通路的激活[54]。在小鼠胚胎成纤维细胞(MEFs)和人源肺部成纤维细胞(IMR90)中Astin C 对IFN-β 基因表达的IC50为分别为4 和10 μmol/L,且在自身免疫疾病模型Trex1-/-小鼠体内注射Astin C 分子能显著改善小鼠多组织器官的免疫损伤。
除了上述靶向STING 蛋白CBD 区域的小分子抑制剂外,靶向抑制STING N 端跨膜区内半胱氨酸的棕榈酰化修饰的小分子抑制剂也逐渐成为研究热点。Mukai 等[55]报道,STING 蛋白N 端结构域的88 位和91 位半胱氨酸(后面分别缩写为Cys88和Cys91)的棕榈酰化修饰为STING 蛋白激活所必须,且其在细胞抗病毒免疫过程中发挥着重要作用。随后Hansen等[56]报道在病毒入侵细胞时产生的内源硝基脂肪酸(NO2-FAs)能与Cys88和Cys91共价结合,阻止STING 蛋白棕榈酰化,抑制STING信号通路激活。在SAVI 患者来源的成纤维细胞中,NO2-FAs能显著抑制疾病伴随的Ⅰ型IFNs及相关炎症因子表达。此外,Haag 等[57]在体外筛选化合物发现硝基呋喃衍生物C-178 和C-176 可以共价结合鼠源STING 蛋白的中Cys91 位点,抑制STING棕榈酰化,阻断下游信号传递。进一步他们筛选出化合物H-151,以相同的方式共价结合人源STING,抑制Ⅰ型IFNs 表达,上述化合物在Trex1-/-小鼠中都能显著改善自身免疫疾病的进展。靶向cGAS和STING蛋白的抑制剂见表3和图4。
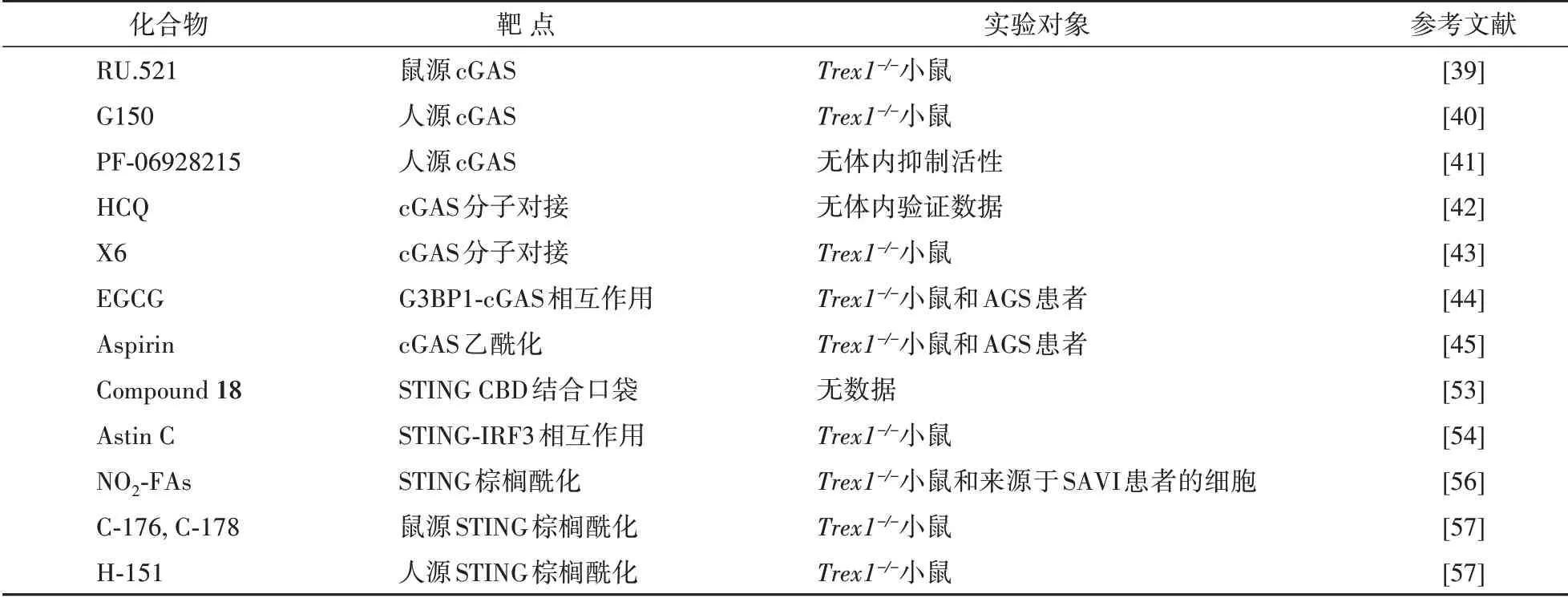
表3 cGAS-STING信号通路抑制剂的炎症抑制活性

图4 cGAS-STING信号通路抑制剂
4 总结和展望
作为固有免疫系统的重要组成部分,cGASSTING 信号通路在维持机体的稳态过程中发挥着重要作用。相应的cGAS-STING 功能的紊乱导致机体免疫功能的异常会诱发包括肿瘤、自身免疫疾病在内的多种疾病。迄今为止,研究人员通过多种筛选方法发现了多种新型的靶向调控cGAS或STING 蛋白功能的小分子激动剂或抑制剂,且这些化合物在临床前的研究中也表现出很好的生物学活性。但是,研究者也发现了现有小分子中存在的一些问题,例如cGAS 的抑制剂抗疟疾药物衍生物的细胞毒性,STING 激动剂CDNs 类似物的细胞通透性差且易被降解,靶向STING 蛋白半胱氨酸的共价抑制剂具有脱靶作用以及在筛选化合物过程中发现的种属特异性问题等。因此,靶向cGAS和STING的新颖激动剂和抑制剂依然有待开发,筛选出具有更好生物活性的先导化合物,为临床研究提供更多的候选分子。
靶向调控cGAS-STING 信号通路的小分子化合物在多种疾病的治疗中都具有显著疗效。比如在小鼠多种肿瘤模型中,STING 激动剂与PD-1(programmed cell death protein 1,PD-1)或者PDL1(programmed cell death 1 ligand 1,PD-L1)抑制剂联用能协同促进机体的抗肿瘤免疫且在体内能形成长期的免疫记忆,有效地控制肿瘤复发;在自身免疫疾病治疗中,AGS 和SAVI 等罕见病并没有有效的治疗药物,临床上目前以使用Janus 激酶抑制剂为主控制疾病进展,与其相比靶向cGAS 或STING 的抑制剂不仅能控制Ⅰ型IFNs 诱导的组织损伤,还能控制TNF-α 和IL-6 等炎症因子导致的组织炎症。此外靶向STING 蛋白的抑制剂也能改善STING 异常激活导致的T,B 细胞和内皮细胞等多种细胞死亡[58]。
此外,在开发调控cGAS-STING 信号通路的激动剂或抑制剂时,其潜在的药物不良反应值得关注。比如在利用STING 激动剂进行肿瘤免疫治疗,很可能爆发“免疫风暴”,过度激活的免疫系统导致机体损伤甚至死亡[2]。在使用抑制剂治疗自身免疫疾病时,过度抑制STING 信号通路抑制剂将导致机体免疫系统低下,使病原体感染和肿瘤发生的风险显著增加。因此,在药物开发中还应注意给药的剂量和周期,尽量避免潜在的药物不良反应发生。
越来越多的研究表明,除了发挥固有免疫功能外,cGAS 和STING 蛋白还能在疾病中独立发挥作用。比如:当DNA 发生损伤时,cGAS 可以入核干扰DNA 同源重组修复过程,促进肿瘤生长,而这一过程不依赖于STING;在白血病中,激活STING将通过IRF3,p53或线粒体介导的细胞凋亡信号通路,促进肿瘤细胞凋亡[58-60]。同时,Caspase 蛋白家族以及microRNA 都可以和固有免疫信号通路发生相互作用,暗示它们也可能影响DNA 识别信号的转导[61-62]。所以在不同疾病的发展中对cGAS和STING蛋白功能进行更加深入研究,将有助于更好地指导靶向药物开发。
猜你喜欢
中老年保健(2022年1期)2022-08-17
保健医苑(2022年5期)2022-06-10
食品安全导刊(2021年20期)2021-08-30
天津医科大学学报(2021年4期)2021-08-21
昆明医科大学学报(2021年2期)2021-03-29
现代临床医学(2021年2期)2021-03-29
中华养生保健(2020年9期)2021-01-18
自我保健(2019年10期)2019-12-11
中国实验动物学报(2019年4期)2019-09-03
安徽医科大学学报(2015年9期)2015-12-16
