
外源褪黑素和脱落酸对干旱胁迫下葡萄生理特性的影响
2020-07-28 04:04王云梅周桂虹刘芯伶林芷仪
干旱地区农业研究 2020年3期
王云梅, 夏 惠, 周桂虹, 刘芯伶, 林芷仪, 梁 东
(1.四川农业大学园艺学院, 四川 成都 611130;2.汶川县科学技术和农业畜牧局, 四川 阿坝藏族羌族自治州 623000)
干旱胁迫导致葡萄生长发育产生生理障碍,产量降低,浆果品质受到影响,是制约葡萄产业发展的重要环境因子之一[1]。褪黑素(melatonin,MT)是一种吲哚类小分子物质,作为一种广谱的生理调节剂存在于绝大多数生物有机体中[2]。在植物体中,MT可以调节植物的生长发育[3],并通过清除活性氧和自由基的强抗氧化作用提高植物对干旱[4]、盐害[5]、重金属[6]、UV辐射[7]、高温[8]、冷害[9]等多种逆境的抗性。脱落酸(abscisic acid,ABA)作为五大植物激素之一,主要起着抑制生长的作用;另一方面,ABA作为植物体细胞内信号传导物质对非生物逆境响应发挥着重要作用。在植物的干旱胁迫信号传导途径中,有ABA-依赖型和ABA-非依赖型两种途径。ABA通过调节植株体内保护酶系统,增强其抵御活性氧毒害的能力,从而缓解干旱胁迫的伤害[10]。而外施ABA对植物的影响与ABA浓度密切相关[11-13]。目前,MT和ABA抗旱作用的研究方法多以PEG渗透胁迫模拟干旱为主,不能真正反映干旱这一自然过程;此外,MT和ABA两种物质结合对干旱胁迫的效果及互作用关系还不清楚。
本研究以盆栽‘阳光玫瑰’葡萄扦插苗为材料,进行MT根灌、ABA喷施、MT+ABA混施和干旱处理,通过测定葡萄叶片MDA、H2O2、光合色素和渗透调节物质的含量以及抗氧化酶活性的变化,探究外源MT和ABA对葡萄干旱胁迫响应的生理机制以及MT和ABA之间的相互作用关系,以期为MT和ABA在果树逆境中的生产应用提供理论依据。
1 材料与方法
1.1 试验材料及处理
试验材料为‘阳光玫瑰’(Shine-Muscat)葡萄扦插苗。于2017年冬季从四川农业大学试验基地采集健壮的‘阳光玫瑰’1 a生枝条,沙藏处理。于2018年3月进行扦插,将枝条剪成长约5 cm左右、带一个饱满芽的枝段,扦插在装有营养土、蛭石和沙子(体积比1∶1∶1)且直径为13 cm、高度为15 cm的营养钵内。待生根后移入规格为(26 cm×22 cm)的塑料盆中,进行常规浇水施肥管理。
待扦插苗长至12~15片真叶时,选取长势一致的‘阳光玫瑰’扦插苗60盆,平均分成5组,每组12盆,每盆1株,放入避雨棚进行处理:(1)正常供水对照(CK);(2)干旱对照处理(Dck);(3)100 μmol·L-1的MT根灌处理(MT);(4)50 μmol·L-1的ABA叶面喷施处理(ABA);(5)100 μmol·L-1MT处理和50 μmol·L-1ABA处理(MT+ABA)。首先,MT组和MT+ABA组用MT预处理:19∶00根灌500 ml的MT溶液,每2 d根灌1次,共4次;同时其它处理组浇500 ml清水。然后,对(2)~(5)组进行自然干旱处理,即不浇水;(1)组正常浇水,每2 d 1次,每次500 ml。ABA处理:在干旱处理第0、2、4、6、8 d和10 d的19∶00对ABA和MT+ABA处理组叶面分别喷施10 ml的50 μmol·L-1的ABA溶液;其它处理组喷施10 ml清水。在干旱胁迫处理的第0 d(以最后一次根灌MT溶液的第二天作为干旱胁迫第0d)、第18 d分别取各组植株4~8叶位的叶片,一部分用于测定相关生理指标,另一部分-80℃液氮速冻后保存于-80℃,用于后续指标的测定。
1.2 测定指标
1.2.1 土壤含水量和水势及叶片相对含水量和水势的测定 土壤含水量采用烘干称重法测定;叶片水势和土壤水势使用型号为WP4C的水势仪进行测定;叶片相对含水量的测定参照Zegaoui等[14]的方法。
1.2.2 丙二醛(MDA)、H2O2和光合色素含量以及渗透调节物质的测定 MDA和H2O2含量、相对电导率测定参照Bai等[15]的方法;渗透调节物质脯氨酸、可溶性糖和可溶性蛋白的含量测定参照李合生等[16]的方法;光合色素含量的测定参照Sariyeva等[17]的方法。
1.2.3 抗氧化酶活性的测定 超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性的测定采用Mirzaee[18]的方法。
1.3 数据处理及统计分析
数据均采用平均值±标准差(SD)表示,比较每个处理组平均值,以P<0.05为差异显著,使用SPSS 20. 0进行单因素方差分析。
2 结果与分析
2.1 不同处理对干旱胁迫下葡萄形态特征的影响
干旱胁迫下植物叶片呈现出不同形态变化,发生叶片黄化卷曲、萎蔫等,胁迫严重时甚至叶片会脱落。从图1可以看出,在控水处理18 d时,干旱对照处理组(Dck)的葡萄植株中、下部出现部分叶片脱落、萎蔫和黄化;MT处理组仅出现叶片黄化,无落叶;ABA处理组葡萄叶片脱落和黄化;MT+ABA处理组与MT处理组表型相似,叶片黄化。正常供水对照处理(CK)的叶片形态特征没有发生明显的变化。

图1 干旱胁迫下‘阳光玫瑰’形态特征的变化Fig.1 The changes of morphological characteristics of ‘Shine-Muscat’ under drought stress
2.2 不同处理对干旱胁迫下土壤水分、葡萄叶片及根系水分的影响
表1数据表明,在控水条件下水分不断蒸发,土壤含水量逐渐降低。控水18 d,干旱对照(Dck)处理的土壤含水量比正常供水处理降低了64.35%。土壤水势的变化与含水量变化一致,呈显著性降低,但各处理间无显著性差异。
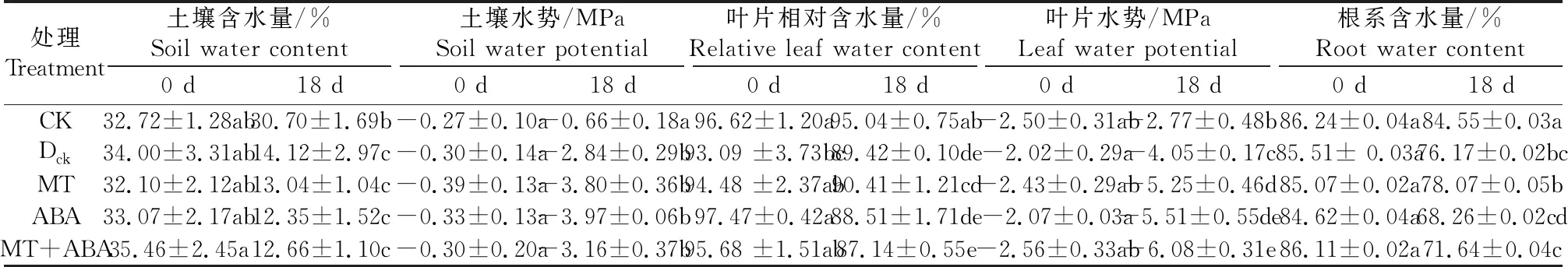
表1 不同处理对干旱胁迫下葡萄土壤含水量、土壤水势、叶片相对含水量、叶片水势和根系含水量的影响
干旱处理下Dck、MT、ABA和MT+ABA处理的叶片相对含水量均显著低于对照(CK)处理。其中MT+ABA处理组最低,仅为CK处理的91.66%,其次是Dck和ABA组,两者无显著差异,MT组的叶片相对含水量高于Dck、ABA和MT+ABA。在干旱胁迫下叶片水势较CK显著降低。与干旱对照Dck相比,MT、ABA和MT+ABA处理进一步降低了叶片水势,在MT+ABA处理下达到最低值-6.08 MPa。
干旱胁迫导致植株根系含水量显著降低。控水处理18 d后,Dck、MT、ABA和MT+ABA处理组的根系含水量都显著低于对照处理(CK);ABA和MT+ABA处理组的幼苗根系含水量显著低于其他干旱处理组,分别达到68.26%和71.64%,说明ABA处理加速了葡萄根系中水分的散失。
2.3 不同处理对干旱胁迫下葡萄叶片膜脂过氧化程度的影响
干旱胁迫导致活性氧大量累积,造成细胞膜脂过氧化。由图2A可知,干旱处理0 d时,MT和MT+ABA处理的葡萄叶片MDA的含量低于其他处理,但差异不显著;正常供水对照处理(CK)的MDA含量在干旱胁迫18 d时显著高于0 d。干旱处理18 d后的葡萄叶片MDA含量显著高于正常供水处理;MT和MT+ABA处理的MDA含量却显著低于干旱对照处理,而ABA处理的MDA含量显著高于其他处理,达到最高值,为20.49 μmol·g-1。
干旱处理0 d,ABA处理的‘阳光玫瑰’幼苗H2O2含量高于CK和MT+ABA处理,但CK、Dck、MT和MT+ABA之间差异不显著。干旱处理18 d后,正常供水对照处理的H2O2含量显著低于干旱处理0 d;Dck处理的H2O2含量显著高于CK处理,达到66.10 mmol·L-1;MT、ABA和MT+ABA处理下H2O2含量显著低于Dck处理,且ABA处理下的含量最低,为27.69 mmol·L-1(图2B)。
植物在逆境胁迫下细胞膜透性增大,相对电导率升高。干旱处理0 d各处理的叶片相对电导率差异不显著。干旱处理18 d后的CK处理与0 d无明显差异;Dck、MT、ABA和MT+ABA处理的相对电导率高于CK对照处理,在ABA单一处理下的相对电导率最高,达到43.15%;MT处理和MT+ABA处理下相对电导率显著低于Dck处理,缓解了干旱胁迫导致的电解质泄漏(图2C)。
2.4 不同处理对干旱胁迫下葡萄光合色素的影响
干旱胁迫会导致植物叶片发黄失绿、萎蔫,甚至脱落。从图3可以看出,干旱处理0 d不同处理葡萄幼苗叶片叶绿素a、叶绿素b、总叶绿素和类胡萝卜素含量均无显著差异。干旱处理18 d后,正常供水处理(CK)相比于0 d各光合色素含量显著增加;干旱胁迫导致各光合色素含量分别下降了14.30%、25.04%、17.23%和20.67%,而在外源MT处理和MT+ABA处理下叶片各光合色素含量均显著增加;ABA处理的各光合色素含量较低,但与MT和MT+ABA处理相比差异不显著,说明外源施用MT有效抑制了干旱胁迫造成的叶片光合色素的降解。
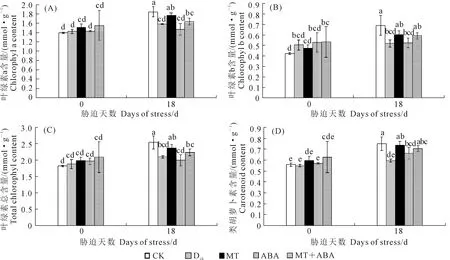
图3 褪黑素和脱落酸对干旱胁迫下葡萄叶片叶绿素a(A)、叶绿素b(B)、总叶绿素(C)和类胡萝卜素(D)含量的影响Fig.3 Effects of MT and ABA on chlorophyll a (A), Chlorophyll b (B), total chlorophyll (C), and carotenoid (D) contents in leaves of grape under drought stress
2.5 不同处理对干旱胁迫下葡萄渗透调节物质的影响
脯氨酸是植物体内非常重要的渗透调节物质,其含量的多少与植物抗旱性密切相关。由图4A可以看出控水处理0 d时,经MT预处理的幼苗游离脯氨酸含量略高于其他处理,但差异不显著。控水处理18 d,干旱处理导致葡萄幼苗游离脯氨酸含量升高了64.82%;MT处理和MT+ABA处理的游离脯氨酸含量显著降低,在MT+ABA处理下达到最低值5.50 mg·g-1,而外源喷施ABA使其含量显著上升,达到最大值15.41 mg·g-1。正常供水处理(CK)其脯氨酸含量与0 d相比差异不显著。
干旱处理0d的葡萄幼苗叶片可溶性糖含量和可溶性蛋白含量均无明显变化。干旱处理18 d时,正常供水(CK)处理的可溶性糖含量与0 d相比无显著性差异,但可溶性蛋白含量降低了26.97%;干旱胁迫(Dck)导致幼苗可溶性糖含量和可溶性蛋白含量迅速增加,分别上升了25.69%和80.03%。在MT、ABA和MT+ABA处理下的可溶性糖含量显著降低;MT+ABA处理的幼苗可溶性糖含量最低,为1.36 mg·g-1。经MT单独预处理,其幼苗可溶性蛋白的含量最低,为251.03 mg·g-1(图4B、4C)。

图4 褪黑素和脱落酸对干旱胁迫下葡萄叶片游离脯氨酸(A)、可溶性糖(B)和可溶性蛋白(C)含量的影响Fig.4 Effects of MT and ABA on the contents of proline (A), soluble sugar (B), and soluble protein (C) in leaves of ‘Shine-Muscat’ under drought stress
2.6 不同处理对干旱胁迫下葡萄抗氧化酶活性的影响
在干旱胁迫下超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)作为保护细胞的重要酶类,能够清除细胞内的活性氧。SOD参与植物体内的氧化代谢,能够催化超氧阴离子发生歧化反应,生成O2和H2O2,保护细胞免遭逆境造成的过氧化伤害。从图5可以看出,干旱处理18 d后,干旱对照处理(Dck)的SOD活性比正常供水处理CK降低了8.88%,而MT、ABA和MT+ABA处理的幼苗SOD活性显著升高,且在MT+ABA处理下其活性显著低于MT处理和ABA处理(图5A)。
POD和CAT能够通过分解H2O2从而对植物起到一定的保护作用。控水处理18 d,幼苗POD活性显著高于正常供水(CK)处理,外源物质处理显著增加了POD活性,且ABA处理下其活性最高,达到了17.91 U·min·mg-1;MT+ABA处理的POD活性显著低于MT处理和ABA处理(图5B)。由图5C可以看出,干旱处理0 d,MT处理降低了CAT活性,干旱18d 时CAT活性相比于正常供水(CK)处理显著升高了59.31%,MT、ABA和MT+ABA处理显著降低了其活性,但3组处理之间差异不显著。

图5 褪黑素和脱落酸对干旱胁迫下葡萄叶片超氧化物歧化酶(SOD)(A)、过氧化 物酶(POD)(B)和过氧化氢酶(CAT)(C)活性的影响Fig.5 Effects of MT and ABA on the activities of SOD (A), POD (B) and CAT (C) activity in leaves of ‘Shine-Muscat’ under drought stress
3 讨论与结论
干旱胁迫下不同外源物质处理会对植物缺水反应产生不同的影响,缓解或加重干旱的伤害,同时使用它们可能会产生协同或拮抗作用。因此,在本研究中,通过外源施用MT、ABA及其相互结合处理,评价与干旱适应相关的表型和生理指标的变化,对葡萄幼苗抗旱性及MT与ABA的相互作用关系进行了研究。
干旱胁迫下单独或共同施用MT和ABA对葡萄幼苗的表型及生理特性有显著的影响。如图1表型所示,MT处理和MT+ABA处理减轻了干旱胁迫对葡萄幼苗叶片产生的黄化、脱落等毒性症状。研究表明光合色素很容易遭受逆境胁迫的伤害[19],本研究中MT处理增加了干旱胁迫下光合色素的含量(图3),这与Kostopoulou等[20]对于MT的研究结果一致,说明MT削弱了植物体内的过氧化作用,缓解了干旱胁迫造成的损伤。50 μmol·L-1的ABA处理对光合色素的影响可能是由于对葡萄幼苗来说该浓度的ABA过高,对植物产生了毒害作用,降低了植物光合系统的稳定性和光合速率,从而阻碍了光合色素的合成或加速了光合色素的降解[12]。MT+ABA处理的作用与MT处理趋近,说明在MT+ABA处理下MT的促进作用缓解了ABA浓度过高所造成的伤害。
叶片水势的高低和相对含水量的多少可以准确地反映干旱胁迫下植物叶片的水分状况和受到土壤缺水的影响程度[21]。水分胁迫下,植物不仅要从低水势的介质中吸收水分来维持植物体内的水分平衡,还要维持压力势基木不变以保证生理生化过程的正常运转,所以通过降低渗透势来进行调节是极其重要的一种方式,而渗透势与细胞内多种渗透调节物质密切相关[22]。干旱胁迫下外源物质处理进一步降低了叶片的水势(表1),可能是MT和ABA的施用虽然降低了脯氨酸、可溶性糖和可溶性蛋白等渗透调节物质的含量,却大幅度提高了山梨醇、淀粉和蔗糖等其他渗透调节物质的水平[23],导致细胞渗透势降低,从而降低了叶片水势。叶君等[24]研究表明MT处理可增加干旱下小麦幼苗的根冠比,而根冠比增加有助于提高植物的吸水能力,维持植物组织含水量。适宜浓度ABA可以促进小麦幼苗的生长,而高浓度ABA则会产生抑制作用[25]。本研究中ABA处理降低了根系含水量,可能是ABA浓度过高对葡萄幼苗产生了抑制作用,加速了葡萄叶片水分的蒸腾,植物为了维持正常的生理代谢从而增强了地下根部水分向地上部的运输。
脯氨酸、可溶性糖和可溶性蛋白作为重要的渗透调节物质,可以维持细胞状态,最大限度地减少水分亏缺带来的不利影响[26]。干旱胁迫导致葡萄幼苗渗透调节物质脯氨酸、可溶性蛋白和可溶性糖的含量显著升高,这种变化是由于渗透调节物质的积累增强了葡萄幼苗自身的渗透调节水平以应对干旱胁迫[27]。外源MT处理降低了渗透调节物质的含量,维持了细胞内渗透调节水平的稳定。ABA处理显著增加了游离脯氨酸的含量,其渗透调节物质整体水平高于MT处理,提高了渗透胁迫的整体水平,而MT+ABA处理的效果与MT处理相似(图4)。由此推测,外源MT与ABA相互作用减轻了ABA浓度过高带来的抑制作用,有效地调节了逆境下植物体内的渗透调节物质水平,维持了植物的正常生理代谢,从而提高了植物的抗旱性。
逆境会打破植物体细胞内自由基产生和清除的平衡,造成活性氧产生增加,膜透性增大,从而破坏细胞膜结构[28-29]。适宜浓度的MT可降低活性氧积累,减轻膜脂过氧化程度,提高植物氧自由基清除能力[30]。王娟[31]研究发现,适宜浓度的ABA能够维持烟草幼苗叶片细胞内环境的稳定性,从而缓解干旱胁迫对烟草幼苗的损伤,而随着干旱处理时间的延长,高浓度ABA喷施处理效果不明显。本试验中,虽然干旱胁迫增加了活性氧对植株造成明显的损伤,但MT处理和MT+ABA处理能够降低MDA和相对电导率水平,控制细胞膜的结构完整性,减轻干旱伤害。相反,ABA处理下MDA含量显著增加,相对电导率升高(图2),推测是由于ABA的浓度过高加重了幼苗的氧化损伤,这与赵春章等[32]的研究结果相似,其研究表明长期高浓度ABA(20 mg·L-1)处理不利于云杉幼苗生长,而MT清除活性氧的能力抵消或者部分减轻了ABA的效应。
植物叶片中抗氧化酶SOD、POD和CAT的协同作用可以有效清除植物体内多余的活性氧,共同抵抗干旱胁迫所造成的氧化损伤,在逆境胁迫中发挥着重要的作用[33]。已有研究表明,干旱胁迫下甘蓝型油菜抗氧化酶活性提高,外源MT显著提高了SOD和POD的活性[34]。适宜浓度ABA还可以诱导某些酶的重新合成从而提高植物的抗旱性[35]。在本研究中,干旱胁迫显著降低了SOD活性,使POD和CAT活性升高,而外源MT处理通过提高SOD和POD活性,并降低CAT活性加强了酶促系统的抗旱能力,对清除植物体内多余活性氧的积累、减轻膜脂过氧化程度产生了一定的积极作用。ABA处理下抗氧化酶的变化与MT相似,这与本试验中其他胁迫指标的变化相反,有可能是ABA浓度过高,长期处理对葡萄产生的消极影响超出了抗氧化系统防御能力的限度,致使SOD和POD的活性达到最高值。与其他胁迫相关指标的变化相似,MT与ABA结合处理下,两者相互作用减弱了高浓度ABA造成的伤害,同时削弱了MT对于干旱胁迫的作用效果,但其具体的作用机理有待于进一步研究。
猜你喜欢
中老年保健(2022年3期)2022-11-21
舰船科学技术(2022年11期)2022-07-15
中国土壤与肥料(2021年5期)2021-12-02
恋爱婚姻家庭·养生版(2021年10期)2021-10-28
安徽化工(2021年3期)2021-05-29
种子(2020年9期)2020-10-22
棉花学报(2020年3期)2020-08-08
农家科技(2020年3期)2020-05-11
现代园艺(2020年7期)2020-04-22
科学与财富(2019年15期)2019-10-21
