
岱衢洋鱼类索饵场仔稚鱼的生态位及其分化研究
2020-09-10 20:53樊紫薇蒋日进徐开达张琳琳印瑞王好学
海洋开发与管理 2020年4期
樊紫薇 蒋日进 徐开达 张琳琳 印瑞 王好学

摘要:为掌握重要鱼类索饵场仔稚鱼的生态分布,促进海洋渔业的发展,文章基于2011—2012年岱衢洋海域4个航次的定量采样,运用生态位宽度、生态位重叠和典范对应分析等方法,分析该海域仔稚鱼群落与海水环境因子的关系。研究结果表明:岱衢洋海域仔稚鱼的生态位宽度可分为广生态位、中生态位和窄生态位,其中广生态位种对资源环境的适应能力强且分布范围较广,而窄生态位种对资源环境的适应能力较弱且分布不均匀;生态位宽度与相对重要性指数呈极显著的正相关关系;仔稚鱼群落生态位重叠指数大于0.6的种对占总种对数的24.24%,生态位重叠程度较低;仔稚鱼分布存在明显的生态分化现象,仔稚鱼的生态位特征和生态习性受温度、盐度、悬浮物和叶绿素a等环境因子的影响较大。
关键词:岱衢洋;仔稚鱼;生态位宽度;生态位重叠;生态位分化
Abstract:Based on the quantitative sampling of 4 voyages in the Daiquyang from 2011 to 2012,using the niche breadth,niche overlap and canonical correspondence analysis,the relationship between the community of fish larvae and water environment factors was analyzed.The results showed that the niche breadth value of the fish larvae can be divided into niche breadth groups,middle niche breadth groups and narrow niche breadth groups.Among them,the species with wide niche which have strong adaptability to the environment and a wide range of distribution,narrow niche species have poor adaptability to the environment.Pearson correlation analysis showed that the niche breadth value was significantly correlated with the relative importance index.The pairs which the niche overlap above 0.6 accounting for 24.24% of the total number,and the degree of ecological overlap was low.The fish larvae showed obvious ecological differentiation.The niche characteristics changes and ecological habit of fish larvae are greatly affected by environmental factors such as temperature,salinity,suspended and chlorophyll a matter.
Key words:Daiquyang,Fish larvae,Niche breadth,Niche overlap,Niche differentiation
0 引言
生态位是指物种繁殖、发育和生存等资源环境特征的组合或排列[1],是生态学研究的重要思想之一[2]。有關生态位的理论已被广泛应用于陆生生物群落的研究[3-8],近年来生态位的研究对象已逐步扩展到海洋生态系统。Korsu等[9]运用生态位测度分析芬兰北部溪流土著种和入侵种鱼类的竞争对栖息地资源环境的利用状况;Galparsoro等[10]通过生态位因子分析法,对西班牙北部比斯开湾内的欧洲龙虾(Homarus gammarus)的适宜栖息地模型进行预测,并绘制栖息地适宜性分布图;于振海等[11]的研究表明,生态位宽度值可更好地反映物种生物量在不同尺度的分布状况;何雄波等[12]认为闽江口主要鱼类生态位的变动受季节变化的影响比较明显;彭松耀等[13]运用生态位宽度和重叠,对南黄海大型底栖动物利用资源环境的状况进行分析,表明物种生态位的动态特征与物种自身的生态习性和栖息环境有密切联系。
上述研究表明,生态位理论在分析海洋生物对资源环境的利用状况、种间竞争关系和种群群落地位等方面具有重要的应用意义和进一步的拓展空间。其中,浮游生物生态位可反映物种和种群在群落中的时空位置及其功能和地位[14-15],是理解浮游生物与生态环境协同适应的重要理论基础,对于认知海洋生态系统的客观发展规律具有深远的意义,然而目前有关浮游生物生态位的研究较少[16-18]。
仔稚鱼是海洋生态系统的重要组成部分,是鱼类资源的补充群体,是海洋生物食物链中的主要饵料生物,在海洋生态系统物质能量传递中扮演关键角色,是渔业生物资源可持续利用的重要基础[19-20]。同时,仔稚鱼的数量分布和生长等与海洋环境变化密切相关。为更好地了解岱衢洋海域仔稚鱼的资源生态状况,本研究进行4个航次的定量采样,综合分析该海域鱼类资源的早期补充机制以及仔稚鱼的生态位及其分化特征,以期为该海域海洋生态系统的稳定和渔业水域环境的修复提供科学依据。
1 材料和方法
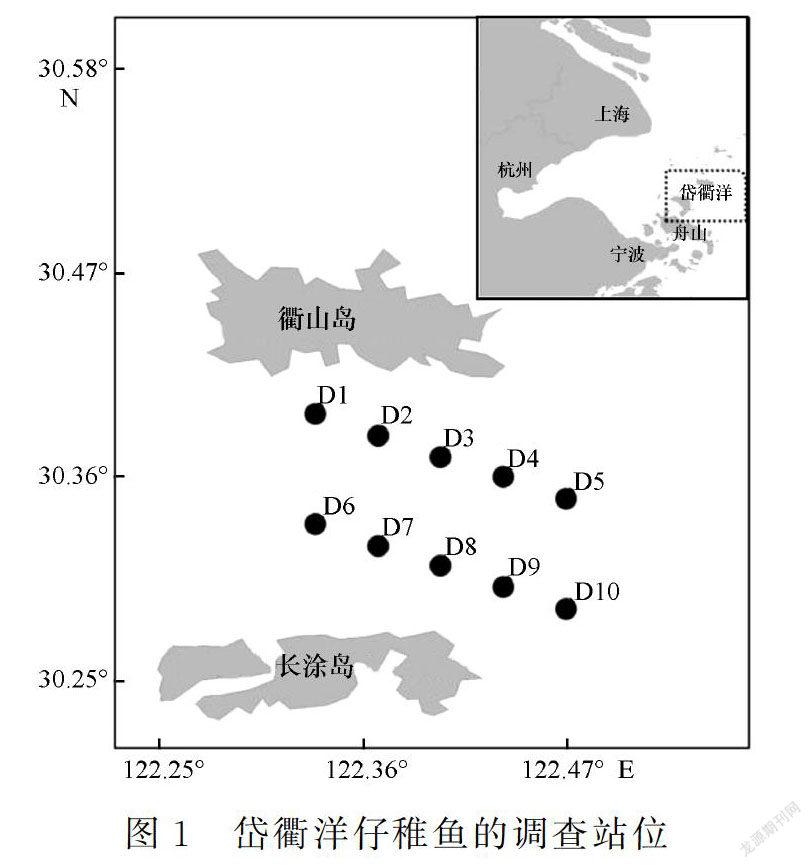
1.1 调查站位和样品采集
岱衢洋位于舟山群岛的长涂岛和衢山岛之间,优越的海洋环境造就著名的“岱衢族”大黄鱼(Larimichthys crocea)的产卵场和索饵场,同时是带鱼(Trichiurus japonicus)等经济鱼类的栖息地[21]。
本研究在岱衢洋海域设置10个调查站位,分别于2011年5月(春)、8月(夏)、11月(秋)和2012年2月(冬)进行4个航次的仔稚鱼调查(图1)。
仔稚鱼的采集方法依据《海洋调查规范:海洋生物调查》(GB/T 12763.6—1991)[22]。利用大型浮游生物网(口径80 cm、长280 cm、网目尺寸0.505 cm)逐站进行表层水平拖网采样,每站拖10 min,拖速为2 n mile/h,网口均配备水平流量计;采用多功能水质检测仪(JFE-AAQ171),对海水的温度(T)、盐度(S)、悬浮物(SPM)、叶绿素a(Chl a)、活性硅酸盐(As)和活性磷酸盐(Ni)等相关环境因子数据进行同步采集;采集的仔稚鱼样品用5%福尔马林溶液固定后带回实验室,根据仔稚鱼的形态特征进行种类鉴定[23-24]并计数。
1.2 数据处理
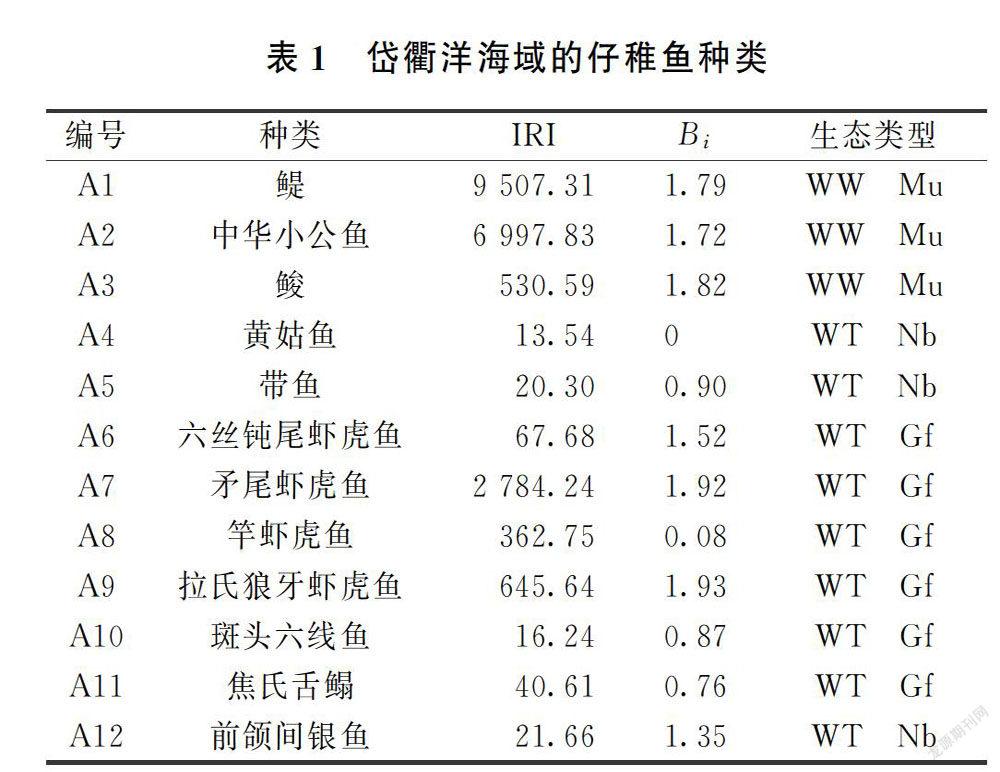
1.2.1 优势种
岱衢洋海域仔稚鱼的优势种采用相对重要性指数[25],计算公式为:
1.2.2 生态位宽度和生态位重叠
生态位宽度指数(Shannon-Weaver指数[26])的计算公式为:
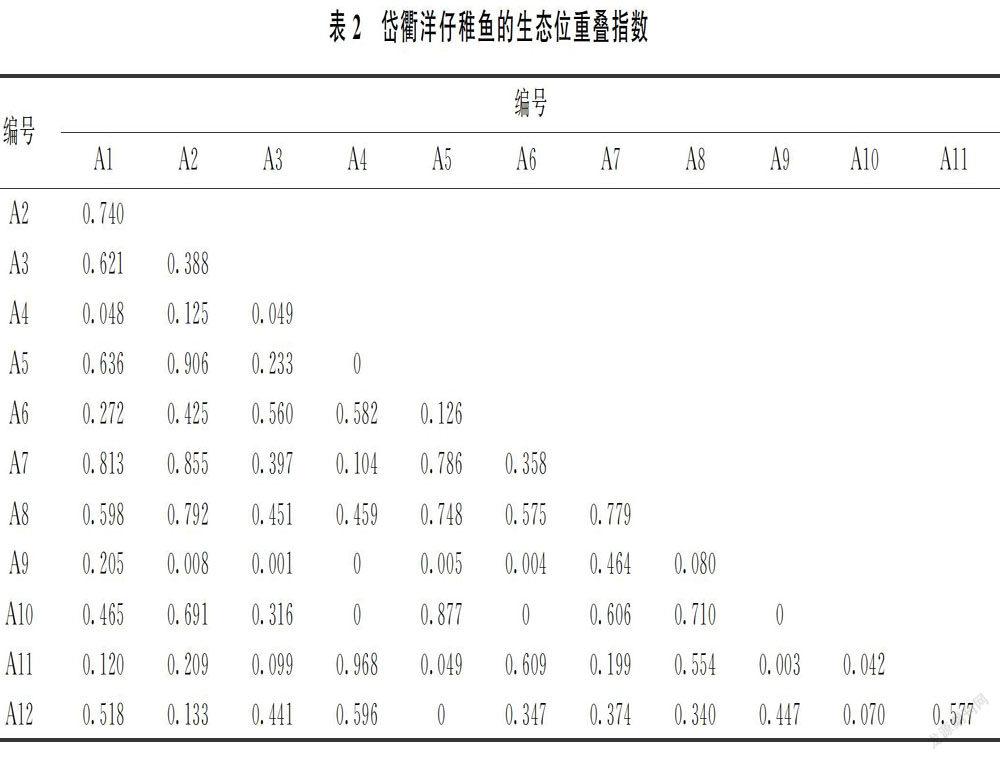
生态位宽度指数(Bi)的取值范围为0~R,其值越大,表示该物种的生态位宽度越大;生态位重叠指数(Qik)的取值范围为0~1。
1.2.3 生态位分化
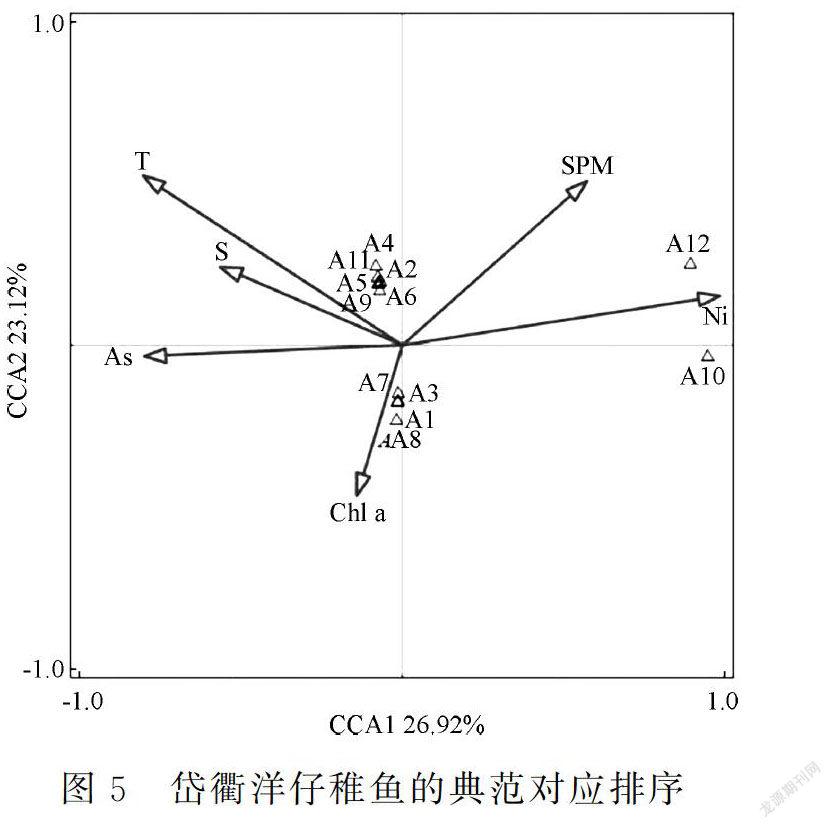
运用Canoco 5.0软件分析仔稚鱼与环境因子之间的关系并绘制排序图,以揭示仔稚鱼在该海域的生态位分化状态。
对物种数据进行除趋势对应分析(DCA),根据每个轴的梯度长度(LGA)选择适宜的排序方法:当LGA<3时,选择冗余分析(RDA);当LGA>4时,选择典范对应分析(CCA);当3 采用Excel 2016软件计算相对重要性指数,采用R语言计算生态位宽度指数和生态位重叠指数;采用Primer 5.0软件对生态位宽度进行聚类分析,采用SPSS 17.0软件进行Pearson相关性分析;在数据分析前对原始数据进行标准化处理。 2 结果 2.1 种类组成 2011年共鉴定出仔稚鱼29种(不包括1个未定名物种),隶属于9目17科,其中有2种鉴定到科。鲈形目种类最多,为7科15种;鲱形目种类次之,为1科4种。 本研究选取IRI>10的仔稚鱼种群作为研究对象。其中,优势种包括鳀(Engraulis japonicus)、中华小公鱼(Stolephorus chinensis)和矛尾虾虎鱼(Chaeturichthys stigmatias)3种,常见种包括(Liza haematocheilus)、拉氏狼牙虾虎鱼(Odontamblyopus lacepedii)和竿虾虎鱼(Luciogobius guttatus)3种,一般种包括黄姑鱼(Nibea albiflora)、带鱼(Trichiurus lepturus)、六丝钝尾虾虎鱼(Amblychaeturichthys hexanema)、斑头六线鱼(Agrammus agrammus)、焦氏舌鳎(Cynoglossus joyneri)和前颌间银鱼(Hemisalanx prognathus)6种(表1)。 2.2 生态位宽度 根据生态位宽度可将仔稚鱼分为3类,即广生态位种(Bi≥1.0)、中生态位种(0.4≤Bi<1.0)和窄生态位种(0 由于黄姑鱼的生态位宽度为0,无法进行聚类分析。对其余11种仔稚鱼的生态位宽度进行聚类分析,可将其划分为3组:①第一组为窄生态位种,包括竿蝦虎鱼1种;②第二组为中生态位种,包括带鱼、斑头六线鱼和焦氏舌鳎3种;③第三组为广生态位种,包括拉氏狼牙虾虎鱼、矛尾虾虎鱼、、鳀、中华小公鱼、六丝钝尾虾虎鱼和前颌间银鱼7种。各种类的生态位宽度越接近,其相似性也越接近(图3)。 由图2和图3可以看出,仔稚鱼种类的聚类分析结果与生态位宽度的分布相一致。 2.3 生态位宽度与相对重要性指数的关系 对岱衢洋海域仔稚鱼的相对重要性指数和生态位宽度进行Pearson相关性分析,二者之间存在极显著的正相关关系(F=0.741,P=0.006<0.01)(图4)。 2.4 生态位重叠 岱衢洋海域仔稚鱼的生态位重叠指数如表2所示。 由表2可以看出,岱衢洋海域仔稚鱼物种之间的生态位重叠不均衡。仔稚鱼种对共有66对,生态位重叠指数的范围为0~0.968。其中:Qik>0.6的种对有16对,占总种对数的24.24%;0.3≤Qik≤0.6的种对有22对,占总种对数的33.33%;Qik<0.3的种对有28对,占总种对数的42.42%;重叠指数为0的种对有6对。黄姑鱼(A4)和焦氏舌鳎(A11)的重叠指数最大(0.968),中华小公鱼(A2)和带鱼(A5)的重叠指数其次(0.906),表明这些种类之间的资源环境利用能力具有相似性。总体而言,岱衢洋海域仔稚鱼的生态位重叠程度较低。 2.5 生态位分化 岱衢洋海域仔稚鱼的群落结构受悬浮物和活性磷酸盐的影响较大。根据仔稚鱼与环境因子的CCA分析结果,第一排序轴(CCA1,特征值为0.99)和第二排序轴(CCA2,特征值为0.85)表现出很高的物种-环境相关性(CCA1和CCA2与环境因子的相关性分别为0.99和0.93)。根据显著性检验结果,第一排序轴和所有排序轴均呈现极显著性差异(第一排序轴:F=1.3,P=0.001;所有排序轴:F=7.0,P=0.002)。其中,第一排序轴与悬浮物和活性磷酸盐呈较大正相关关系,而与活性硅酸盐和温度呈较大负相关关系。中华小公鱼、黄姑鱼、带鱼、六丝钝尾虾虎鱼、拉氏狼牙虾虎鱼和焦氏舌鳎等仔稚鱼受温度和盐度的影响较大,而鳀、、矛尾虾虎鱼和竿虾虎鱼等仔稚鱼受叶绿素a的影响较大。在排序图中,受不同环境因子的影响,仔稚鱼的群落分布呈明显的两极分化状态(图5)。 3 讨论 3.1 生态位宽度 生态位是物种和种群对资源环境生态适应的结果,为某区域内物种群落的生态动态特征提供简单和可靠的解释[29]。生态位宽度是反映物种和种群利用资源环境多样化程度的重要指标[30]。 鳀、中华小公鱼和矛尾虾虎鱼是岱衢洋海域的优势种,且为广生态位种。一方面,该海域的鱼类资源结构正向小个体、低营养级和低经济价值的中上层鱼类种群演替,群落中的优势种如大黄鱼、小黄鱼和带鱼等经济种类逐渐被鳀、小公鱼和虾虎鱼等饵料性鱼类所取代[31];另一方面,该海域位于亚热带季风性气候区,同时受台湾暖流的影响,优势种为暖水种和暖温种,其均为广生态位种,分布均匀且活动范围较大,这些种类的分布与其利用资源环境的能力和环境适应性有较大关系。 根据Pearson相关性分析,岱衢洋仔稚鱼的相对重要性指数与生态位宽度存在极显著的正相关关系,但二者并非为完全的正相关关系,如常见种和拉氏狼牙虾虎鱼的生态位宽度均大于优势种鳀和中华小公鱼,这可能与计算方法和考虑的生态特征有所不同有关。相对重要性指数与物种的丰度、生物量和出现站位占比等相关,但生态位宽度不仅考虑上述因素,而且综合考虑不同站位之间物种的分布差异。 根据聚类分析,可将仔稚鱼的生态位宽度分为3组,其中第三组的生态位宽度最大,主要包括鳀、、矛尾虾虎鱼和拉氏狼牙虾虎鱼等分布广和环境适应性强的物种;第二组的生态位宽度较大,但斑头六线鱼、带鱼和焦氏舌鳎在各站位的数量分布不均匀;第一组的生态位宽度最小,这是由于近年来受环境恶化和过度捕捞的影响,产卵场破坏严重,黄姑鱼的资源量减少[32],而竿虾虎鱼生长缓慢,物种在各站位的数量分布极不均匀。李显森等[33]认为生态位宽度的聚类分析可反映群落中物种的分布格局,即生态位宽度越大,该物种的数量分布越均匀。这与本研究结果一致。 3.2 生态位重叠 生态位重叠体现物种对同一生境的利用和物种间的配置状况,可反映物种对资源环境利用的相似程度和竞争程度[34]。根据对生态位宽度的分析,生态位宽度较大的物种对资源环境的利用能力和竞争能力较强,反之则较弱。然而本研究发现,生态位重叠指数最大的种对,其生态位宽度并不一定较大,例如:黄姑鱼(A4)和焦氏舌鳎(A11)的生态位重叠指数最大(0.968),而前者的生态位宽度为0,后者为中生态位种。黄姑鱼和焦氏舌鳎的生态位重叠指数最大的原因在于二者具有极为相似的栖息环境和相同的生态习性:二者均为暖温性中下层鱼类;黄姑鱼栖息于泥沙底质较浅的沿岸海域,最适宜的生长水温为24℃~28℃,产卵期为5—7月[35];焦氏舌鳎生活于砂泥底质海域,最适宜的生长水温为24℃~28℃,鱼卵和仔稚鱼出现于5—9月[36]。此外,黄姑鱼(A4)和拉氏狼牙虾虎鱼(A9)的生态位重叠指数为0,这是由于:①拉氏狼牙虾虎鱼栖息于水深为2~8 m的泥沙底质海域,于每年2—4月和7—9月在咸淡水水域产卵,与黄姑鱼的产卵时间不同;②黄姑鱼为洄游性种类,而拉氏狼牙虾虎鱼为地方性种类,二者生态习性不同。 同时,生态位重叠指数与物种分布环境位点的重合情况密切相关[37]。Wathne等[38]认为当生态位重叠指数大于0.6时,种间生态位重叠较显著,且物种分布环境位点的重合度较高。岱衢洋海域仔稚鱼的生态位重叠指数差异较大,物种之间存在不同程度的重叠:①鳀、中华小公鱼、和矛尾虾虎鱼等广生态位种与其他物种的生态位重叠指数大于0.6的概率较高;②生态位重叠指数为0的种对有6对,表明这些物种分布环境位点的差异较大,这与物种自身的生态特征和栖息环境等密切相关。 3.3 生态位分化 海洋生态系统与资源环境之间的相互作用基于多重的时间和空间尺度。相对于其他海洋生物群落而言,仔稚鱼的自主运动能力较弱,具有随波逐流的特点,因此环境因素对仔稚鱼的生态分布发挥主导作用[17]。 沿第一排序轴,仔稚鱼在温度、盐度、悬浮物和叶绿素a 4个梯度上的分化十分明显:温度是影响鱼类游泳能力的重要因素,鱼类通过调整自身的生理和生化过程以适应温度的变化,从而影响其游泳能力[39];盐度不仅影响鱼类的代谢活动和渗透压调节,而且影响鱼类的性腺发育、组织结构和胚胎孵化等生理功能[40];叶绿素a的浓度与浮游植物的分布密切相关,而浮游植物是仔稚鱼的饵料生物,从而间接影响仔稚鱼的数量[41]。中华小公鱼是中上层鱼类,对温度的反应非常敏感;黄姑鱼和带鱼是洄游性种类,春、夏季沿岸海水升温,鱼类由深水区向近岸海域洄游,而秋、冬季则由近岸海域向深水区洄游;鳀和等在早期发育阶段的食物来源与浮游植物密切相关,因此受叶绿素a浓度的影响较大。 典范对应分析(CCA)以环境因子为主体,通过排序来判断物种在生境中的生态分化情况,可对生态位及其重叠无法合理解释的结果做出补充说明[42]。例如:①黄姑鱼(A4)和焦氏舌鳎(A11)对资源环境利用的相似程度较低,但二者均分布在温度较高的海域,具有相似的资源环境需求,因此二者的生态位宽度不同而生态位重叠指数较大;②六丝钝尾虾虎鱼(A6)分布于温度和盐度较高的海域,而前颌间银鱼(A12)受悬浮物的影响较大,因此二者虽均属广生态位种,但存在空间分布的差异,生态位重叠指数较小;③2011年岱衢洋海域水体呈中度污染和轻度富营养化状态[43],因此大部分仔稚鱼生活在活性硅酸盐和活性磷酸盐浓度较低的海域;④前颌间银鱼(A12)为广生态位种,但与其他种类的生态位重叠指数均较低,CCA排序图显示其分布于悬浮物浓度较高的海域,而其他种类分布于悬浮物浓度较低的海域,生态适应性的差异使其存在空间分布的差异,并呈现生态位分化的特征;⑤在排序轴上分布差异较大的物种,其生态位重叠指数均较低。综上所述,受不同环境因子的影响,仔稚鱼的分布呈现明显的生态分化现象。 参考文献 [1] 郑光美.鸟类学[M].北京:北京师范大學出版社,1995. [2] 張金屯.数量生态学[M].北京:科学出版社,2004. [3] CODY M.Niche theory and plant growth form[J].Vegetatio,1991,97:39-55. [4] DECANS T,MARGERIE P,RENAULT J,et al.Niche overlap and species assemblage dynamics in an ageing pasture gradient in north-western France[J].Acta Oecologica,2011,37(3):212-219. [5] 刘金福,洪伟.格氏栲群落生态学研究:格氏栲林主要种群生态位的研究[J].生态学报,1999,19(3):59-64. [6] 苏志尧,吴大荣,陈北光.粤北天然林优势种群生态位研究[J].应用生态学报,2003,14(1):25-29. [7] 刘贵河,王国杰,汪诗平,等.内蒙古典型草原主要草食动物食性及其营养生态位研究:以大针茅群落为例[J].草地学报,2013,21(3):439-445. [8] 张晶,赵成章,任悦,等.张掖国家湿地公园优势鸟类种群生态位研究[J].生态学报,2018,38(6):2213-2220. [9] KORSU K,HEINO J,HUUSKO A,et al.Specific niche characteristics facilitate the invasion of an alien fish invader in boreal streams[J].International Journal of Ecology,2012,1:1-10. [10] GALPARSORO I,BALD J,LIRIA P,et al.Predicting suitable habitat for the European lobster(Homarus gammarus),on the Basque continental shelf(Bay of Biscay),using Ecological-Niche Factor Analysis[J].Ecological Modelling,2009,220(4):556-567. [11] 于振海,金显仕,李显森.黄海中南部主要鱼种的生态位分析[J].渔业科学进展,2010,31(6):1-8. [12] 何雄波,李军,沈忱,等.闽江口主要渔获鱼类的生态位宽度与重叠[J].应用生态学报,2018,29(9):3085-3092. [13] 彭松耀,李新正,王洪法,等.南黄海春季大型底栖动物优势种生态位[J].生态学报,2015,35(6):1917-1928. [14] ROSENTHAL G.Selecting target species to evaluate the success of wet grassland restoration[J].Agriculture,Ecosystems and Environment,2003,98(1/3):227-246. [15] JORGE S A,TOWNSEND P.Interpretation of models of fundamental ecological niches and species′distributional areas[J].Biodiversity Informatics,2005,2:1-10. [16] 徐晓群,曾江宁,陈全震,等.浙江三门湾浮游动物优势种空间生态位[J].应用生态学报,2013,24(3):818-824. [17] 梁淼,姜倩,孙丽艳,等.曹妃甸近岸海域大、中型浮游动物优势种空间生态位研究[J].生态环境学报,2018,27(7):1241-1250. [18] 李兴,李建茹,李畅游.内蒙古乌梁素海浮游植物优势种的生态位分析[J].水生态学杂志,2017,38(6):40-47. [19] 赵静,章守宇,汪振华,等.嵊泗人工鱼礁海域鱼类和大型无脊椎动物群落结构特征[J].中国水产科学,2010,17(5):1045-1056. [20] 戴媛媛,侯纯强,杨森,等.天津海域人工鱼礁区浮游动物群落结构及其与环境因子的相关性研究[J].海洋湖沼通报,2018(5):163-170. [21] 张洪亮,徐开达,朱增军,等.岱衢洋鱼类资源及其群落多样性的季节变化[J].水产学报,2012,36(4):601-607. [22] 国家技术监督局.海洋调查规范:海洋生物调查(GB/T 12763.6-1991)[S].北京:中国标准出版社,1992. [23] 张仁斋,陆穗芳,赵传絪,等.中国近海鱼卵与仔鱼[M].上海:上海科学技术出版社,1985. [24] 万瑞景,张仁斋.中国近海及其邻近海域鱼卵与仔稚鱼[M].上海:上海科学技术出版社,2016. [25] PINKAS L,OLIPHANT M S,IVERSON I L K.Food habits of albacore,Bluefin tuna,and bonito in California waters[J].Fish Bulletin,1971,1971(152):1-105. [26] SHANNON C E,WEAVER W.The mathematical theory of communication[M].Chicago:University of Illinois Press,1963. [27] PIANKA E R.The structure of lizard communities[J].Annual Review of Ecology and Systematics,1973,4:53-74. [28] PETR J L.Multivariate analysis of ecological data using CANOCO[M].New York:Cambridge University Press,2003. [29] HUTCHINSON G E.Cold spring harbor symposium of quantitative biology[J].Concluding Remarks,1957,22:415-427. [30] 宋永昌.植被生态学[M].第二版.北京:高等教育出版社,2017. [31] 赵淑江,吕宝强,李汝伟,等.物种灭绝背景下东海渔业资源衰退原因分析[J].中国科学:地球科学,2015,45(11):1628-1640. [32] 逯云召,宓慧菁,于燕光,等.天津地区黄姑鱼人工繁殖技术[J].河北渔业,2018(2):43-45,56. [33] 李显森,于振海,孙珊,等.长江口及其毗邻海域魚类群落优势种的生态位宽度与重叠[J].应用生态学报,2013,24(8):2353-2359. [34] SPIES T A,FRANKLIN J F,KLOPSCH M.Canopy gaps in Douglas-fir forests of the Cascade Mountains[J].Canadian Journal of Forest Research,1990,20:649-658. [35] 陈庆凯.黄姑鱼规模化人工育苗及养殖技术的研究[D].厦门:集美大学,2016. [36] 杨东莱,吴光宗,庞鸿艳.渤海湾的半滑舌鳎及焦氏舌鳎的鱼卵和仔、稚鱼的季节分布[J].生态学杂志,1984(3):30-33. [37] 葛宝明,鲍毅新,郑祥,等.灵昆岛潮间带大型底栖动物群落结构与生态位分析[J].生态学报,2005,25(11):245-251. [38] WATHNE J A,HAUG T,LYDERSEN C.Prey preference and niche overlap of ringed seals Phoca hispida and harp seals P.groenlandica in the Barents Sea[J].Marine Ecology Progress Series,2000,194:233-239. [39] 徐革锋,牟振波,韩英,等.温度和流速对细鳞鲑(Brachymystax lenok)幼鱼游泳能力的影响及其与呼吸代谢的关系[J].海洋与湖沼,2015,46(4):786-792. [40] 童燕,陈立侨,庄平,等.急性盐度胁迫对施氏鲟的皮质醇、代谢反应及渗透调节的影响[J].水产学报,2007(S1):38-44. [41] 高生泉,林以安,金明明,等.山东半岛以南水域营养盐的时空分布及其与鱼产卵场的关系[J].海洋学报(中文版),2003(S2):157-166. [42] 郑挺,林元烧,曹文清,等.北部湾北部生态系统结构与功能:浮游动物空间生态位及其分化[J].生态学报,2014,34(13):3635-3649. [43] 李振华,张亚洲,周永东,等.岱衢洋海域2010-2011年网采浮游植物群落特征与水质评价[J].浙江大学学报(理学版),2017,44(3):327-338.
