
中国大陆近海菖鲉属鱼类新记录种
——三色菖鲉(Sebastiscus tertius) 的形态特征与DNA 条形码研究*
2020-10-14 04:05贾程豪高天翔徐胜勇
海洋与湖沼 2020年5期
贾程豪 高天翔 徐胜勇 宋 娜①
(1. 中国海洋大学海水养殖教育部重点实验室 青岛 266003; 2. 浙江海洋大学水产学院 舟山 316022)
菖 鲉 属 (Sebastiscus) 隶 属 于 辐 鳍 鱼 纲(Actinopterygii)、新鳍鱼亚纲(Neopterygii)、鲉形目(Scorpaeniformes)、鲉亚目(Scorpaenoidei)、鲉科(Scorpaenidae), 主要分布于西太平洋以及印度尼西亚海域地区(Gloerfeltet al, 1984)。菖鲉属鱼类曾被划分为平鲉属(Sebastes)的一部分, 直到 1904 年Jordan 和Starks 根据鱼鳔的发达程度以及第二眶下骨的不同对其鉴别, 从而将其作为一个独立的属(Jordanet al, 1904; Matsubara, 1943a, b)。也曾经有学者想要把菖鲉属作为平鲉属的亚属进行分类的报道(Barsukovet al, 1978), 但目前多数文献都将菖鲉属作 为 一 个 独 立 的 有 效 属(Amaoka, 1984; Shimizu, 1984a, b; Washingtonet al, 1984)。
目前菖鲉属被认为存在四个物种, 分别是: 白斑菖鲉(Sebastiscus albofasciatus)、褐菖鲉(Sebastiscus marmoratus)、三色菖鲉(Sebastiscus tertius) (Lacepède, 1802; Cuvieret al, 1829; Barsukovet al, 1978)以及日本学者 2018 年报道的一个菖鲉属新种(Sebastiscus vibrantus) (Morishitaet al, 2018)。其中, 褐菖鲉广泛分布于西北太平洋, 其他种类都主要分布于东亚和印度尼西亚的温暖水域(成庆泰等, 1962; 朱元鼎等, 1963; 成庆泰, 1997; Nakaboet al, 2013; Morishitaet al, 2018)。
三色菖鲉(Sebastiscus tertius), 俗称三色石狗公, 是菖鲉属中体型较大的种类, 最大可长到30cm 以上(陈大刚等, 2015; Morishitaet al, 2018)。该物种主要分布于朝鲜半岛、日本、中国台湾以及印度尼西亚东部海域(Nakaboet al, 2013)。目前, 关于三色菖鲉的研究报道较少, 仅见国外关于该物种形态学以及对其温度光照胁迫影响的研究(Tokimuraet al, 2001; Baeket al, 2004; Limet al, 2014)。在我国, 到目前为止所记录到的三色菖鲉均来自台湾地区 (邵广昭等, 1993), 香港地区虽有存在该物种的报道, 但并未有照片和标本为佐证(Yvonneet al, 2000), 而中国大陆其他地区则没有记录(成庆泰等, 1962; 朱元鼎等, 1963; 金鑫波, 1985; 成庆泰, 1997; 李金平等, 1999; 徐学军, 2001; 邓思明等, 2006; 刘敏等, 2013)。
DNA 条形码(DNA barcoding)由Hebert 等(2003a)提出, 是一种对一段或多段目标基因进行分析以识别物种的分子鉴定技术。微型DNA 条形码是指长度明显小于600bp 的一段序列(程鹏等, 2012), 由于部分样品在保存过程中其DNA 发生了降解, 很难得到完整的DNA 条形码, 相比而言, 200bp 左右的序列更易扩增, 进而进行物种鉴定, Cywinska 等(2006)通过应用该技术对蛾类物种进行研究, 结果表明通过选取特定的200bp 左右长度的序列可以有效对物种进行鉴定。当前DNA 条形码和微型DNA 条形码已被广泛运用于新物种和新记录种的发现以及对物种的鉴定等研究当中(彭居俐等, 2009; 程鹏等, 2012; Cywinskaet al, 2006; Gaoet al, 2011; Qinet al, 2013; Fanget al, 2015)。本研究记录了从浙江舟山海域采集的三色菖 鲉, 在对其形态特征进行描述的同时还测定了该物种的COI 基因片段序列以及12S rRNA基因片段短序列, 并与GenBank 中的相关同源序列进行了比较分析, 为中国菖鲉属鱼类的分类提供理论依据, 同时为中国大陆近海菖鲉属鱼类研究提供基础资料。
1 材料与方法
1.1 实验材料
三色菖 鲉标本于2018 年4 月20 日采自浙江省舟山市近海(坐标: 30.0607°N, 122.3546°E), 共19 尾, 体长167.1—254.3mm。标本保存于中国海洋大学渔业生态学实验室(OUC-FEL), 标本编号: OUC_FEL 20180401—20180419。
1.2 实验方法
1.2.1 形态学研究 本研究采用传统形态学测量方法, 用直尺直接测量样品的全长、体长, 精度精确至 0.1mm; 用游标卡尺测量样品的测量性状, 精度精确至 0.01mm, 在本研究中, 具体测量性状包括: 体宽、体高、头长、吻长、尾柄长、尾柄高、上颌长、眼径、眼间距、眼后头长、胸鳍长、腹鳍长、背鳍基长、臀鳍基长; 用电子天平测量样品体质量, 精确至0.01g。对样品的计数性状进行直接计数, 计数性状包括: 背鳍数、胸鳍数、腹鳍数、臀鳍数、尾鳍数、鳃耙数、鳃弓数、脊椎骨数。
三色菖鲉的可量性状及其测度依据见图1。其中, 上颌长(UJL)和眼间距(ID)以实际测度数据为依据, 未在图中标注。

图1 三色菖鲉可量性状特征 Fig.1 The schematic diagram of morphologic measurement of S. tertius
1.2.2 遗传学研究 取19 尾所得样品尾部的适量肌肉组织, 采用酚-氯仿方法提取肌肉中的DNA, 并在将所提取DNA 溶解后置于4°C 冰箱保存备用。
用于扩增样品线粒体DNA 12S rRNA 片段的引物为MiFish-U-F 和MiFish-U-R (Miyaet al, 2015), 引物序列为: MiFish-U-F: 5′-TTTGTCGGTAAAACT CGTGCCAGC-3′, MiFish-U-R: 5′-CAAACTGGGAT TAGATACCCCACTAT AA-3′。用于扩增样品线粒体DNA COI 片段的引物为FishF1 和FishR1 (Wardet al, 2005), 引 物 序 列 为: FishF1: 5′-TCAACCAACCAC AAA GACATTGGCAC-3′, FishR1: 5′-TAGACTTC TGGGTGGCCAAAGAATCA-3′。
PCR 反应体系为25μL, 包括0.25μL 的0.15μL的EasyTaq 酶(北京全式金生物技术有限公司), 各反应体系组分参照俞正森等(2017)研究中所用PCR 反应体系。PCR 扩增所需反应条件参照Fang 等(2015)研究中所用反应条件。所用PCR 仪为TaKaRa PCR Thermal Cycler MP (TP3000)。以上反应均设阴性对照以排除产生引物二聚体以及DNA 污染的情况。在进行1%琼脂糖凝胶电泳检测(U=5V/cm)进行确认之后, 将所得PCR 产物送至青岛擎科生物技术有限公司进行双向测序。
将测得的12S rRNA 以及COI 基因片段序列用DNAStar(USA)软件包中的SeqMan 软件进行人工校正和序列比对。从GenBank 中下载所有拉丁学名为Sebastiscus tertius的同源序列以及部分拉丁学名为Sebastiscus marmoratus的同源序列(表1, 表2)进行分析, 使用MEGA5.0 鲉软件以平 科鱼类的同源序列(表1, 表2)作为外群构建系统发育树, 并计算组群间和组群内的遗传距离。
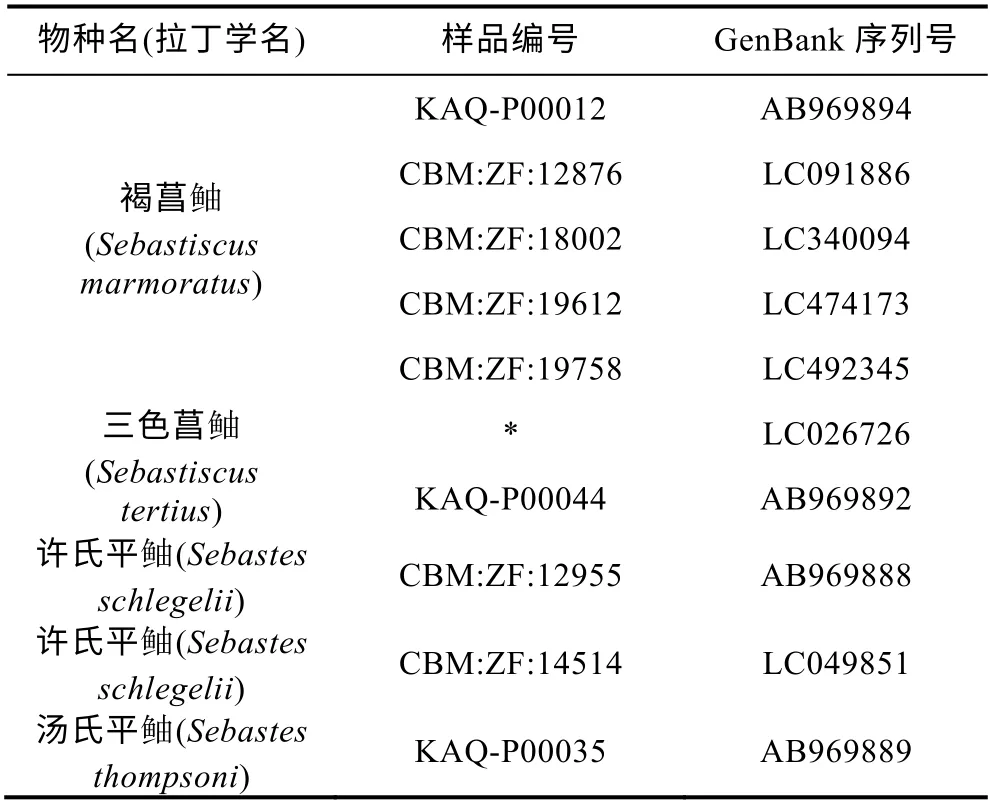
表1 本研究所用12S rRNA 序列信息(*表示信息缺失) Tab.1 Information of the 12S rRNA sequences cited in this study (*: data missing)
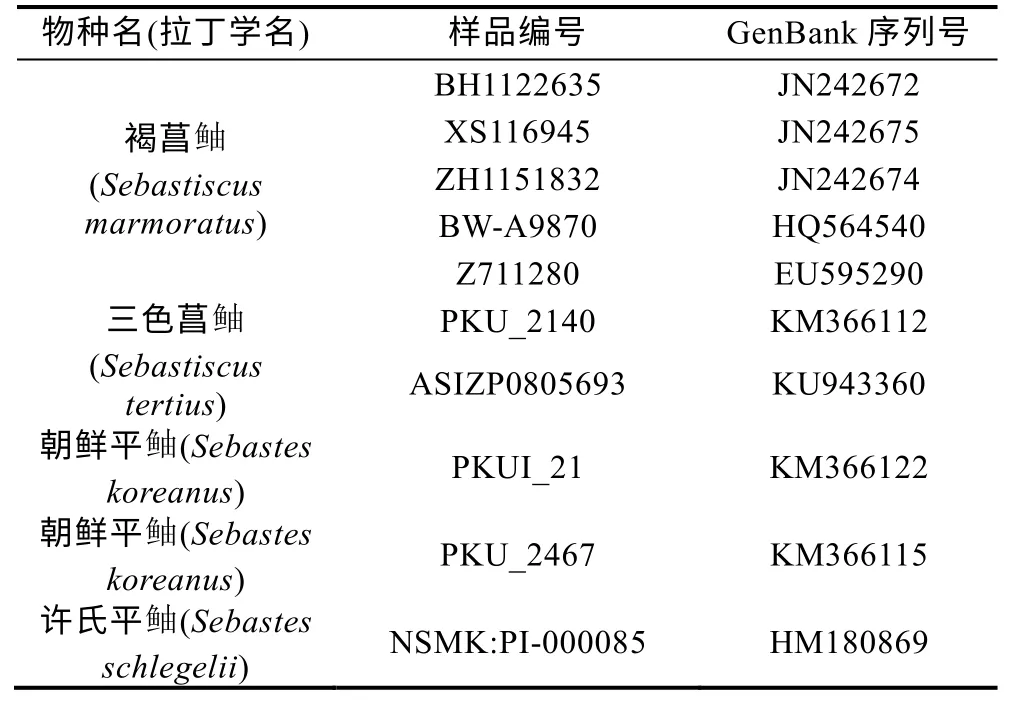
表2 本研究所用COI 序列信息 Tab.2 Information of the COI sequences cited in this study
2 结果
2.1 三色菖鲉 Sebastiscus tertius (Barsukov and Chen, 1978)形态特征(图2)
基本体型和体色见图2。所用19 尾标本鱼体呈长椭圆形(侧面观), 体色呈红色到淡粉色。所得标本均身披栉鳞。头大, 头部棘棱显著, 眼上侧位。眶前骨下后角有1 枚棘。前鳃盖骨有5 枚棘, 鳃盖骨有2 枚棘。胸鳍较为宽大, 且胸鳍基底中部有一较大暗斑。
三色菖 鲉的可数性状见表3。其中, 背鳍数、腹鳍数、腹鳍数、臀鳍数、鳃弓数和脊椎骨数均为定值; 胸鳍鳍条数范围为18—19, 大部分为19(71%); 尾鳍鳍条数范围为 13—17, 大部分为 16(46%); 上鳃耙数范围为6—7, 下鳃耙数范围为13—17, 其中上鳃耙数为7, 下鳃耙数为16 的个体较多(25%)。

图2 三色菖鲉(OUC_FEL 180404), 254.3mm SL, 采自舟山近海 Fig.2 S. tertius (OUC_FEL 180404), 254.3mm SL, collected from coastal waters of Zhoushan

表3 舟山近海的三色菖鲉标本的可数性状及菖鲉属鱼类可数性状参考数值(*表示信息缺失) Tab.3 Countable characters of S. tertius specimens from Zhoushan coastal waters and the reference value for Sebastiscus fish (*: data missing)
可量性状比见表 4。其中, 全长为体长的1.2—1.3 倍, 体长为体高的2.7—3.4 倍, 为头长的2.4—2.8 倍, 为眼后头长的4.0—5.5 倍。头长为眼径的3.7—5.5 倍, 为眼间距的4.8—8.5 倍, 为上颌长的4.5—6.4 倍。体长为尾柄长的5.4—7.1 倍, 尾柄长为尾柄高的1.5—2.1 倍。

表4 舟山近海的三色菖鲉标本的可量性状比 Tab.4 Ratios between measurable characters of S. tertius from Zhoushan
2.2 DNA 条形码分析
2.2.1 12S rRNA 基因片段分析 对所得标本中的19 尾三色菖鲉的12S rRNA 基因片段进行双向测序后, 共获得 19 条长度为 168bp 的序列, 其GenBank 号为MN854386—MN854404。将本研究所得序列与GenBank 中学名为Sebastiscus tertius以及5条与三色菖 鲉同属的学名为Sebastiscus marmoratus的同源序列(表1)进行比较, 采用邻接法以汤氏平 鲉和许氏平 鲉为外群构建系统发育树, 结果表明, 在该系统树上明显分为两个组群(图3)。其中, GenBank 中序列号为LC026726 和AB969892 的学名为Sebastiscus tertius的2 条序列与本研究的19尾三色菖 鲉的12S rRNA 序列先聚类, 该组内平均遗传距离为0.000, 而GenBank 中的学名为Sebastiscus marmoratus的5 条序列聚为一支, 该组内平均遗传距离为0.002 (0.000—0.006), 两组群间的遗传距离为0.026。
2.2.2 COI 基因片段序列分析 对所得标本中的19 尾三色菖鲉的COI 基因片段进行双向测序, 共获得 19 条长度为652bp 的序列, 其GenBank 号为MN863509—MN863527。将本研究所得序列与GenBank 中学名为Sebastiscus tertius以及5 条与三色菖鲉同属的学名为Sebastiscus marmoratus的同源序列(表2)进行比较, 采用邻接法以朝鲜平鲉和许氏平鲉为外群构建系统发育树, 结果表明, 在该系统树上明显分为两个组群(图4)。其中, GenBank 中序列号为KM366112 和KU943360 的学名为Sebastiscus tertius的2 条序列与学名为Sebastiscus marmoratus的5 条序列先聚类, 该组内的平均遗传距离为0.002 (0.000—0.004), 而本研究的19 尾三色菖鲉聚为一支, 该组内的平均遗传距离为0.002 (0.000—0.011), 两组群间的遗传距离为0.044。
3 讨论
浙江海域原有记录的菖鲉属鱼类仅1 种, 即褐菖鲉(吴常文, 1999; 赵盛龙等, 2006; 俞存根等, 2010)。中国大陆近海海域原有记录的菖鲉属有2 种, 分别是褐菖鲉和白斑菖鲉(金鑫波, 1987)。本次研究所发现的三色菖鲉为浙江省海域记录的第2 种菖鲉属鱼类, 也是我国除台湾海域外所记录到的第3 种菖鲉属鱼类。本研究通过对舟山海域获得的三色菖鲉标本分别进行传统形态学以及分子生物学上的鉴定, 为中国近海海域菖鲉属鱼类的鉴定提供依据。
三色菖鲉的原记录分布范围为朝鲜半岛、日本、台湾以及印度尼西亚东部的海域(Nakaboet al, 2013), 于我国原本只记录于台湾海域(邵广昭等, 1993), 在我国大陆近海尚未有所记载。本研究所得样品增补了该物种的分布记录, 从我国近海范围来看, 本研究将其分布范围从我国台湾海域扩大到东海北部的舟山海域; 从世界范围来看, 本研究填补了北方朝鲜半岛、日本海域的已有分布记录与南方台湾、印度尼西亚东部海域的已有分布记录之间的空白海域, 完善了三色菖鲉的世界分布范围记录。

图3 基于12S rRNA 序列, 以汤氏平鲉和许氏平鲉为外群运用邻接法构建的三色菖鲉系统发育树 Fig.3 Phylogenetic tree of Sebastiscus tertius using Sebastes thompsoni and Sebastes schlegelii as outgroups based on neighbor-joining method and 12S rRNA sequences
在菖鲉属鱼类中, 三色菖鲉与褐菖鲉形态非常相似。有学者将三色菖鲉认为是生活在深层海域的褐菖鲉(Matsubara, 1943a)。在褐菖鲉与三色菖鲉的比较研究中, 没有单独的形态特征可以较为明显的区分两个物种, 只能通过多种性状的结合如胸鳍鳍条数的数量、鳃耙的数量和鱼体的颜色来进行识别(Barsukovet al, 1978)。在Barsukov 等(1978)的研究中, 褐菖鲉和三色菖鲉之间最大的不同体现为胸鳍鳍条的数量: 大多数(80.7%)的褐菖鲉有18 个, 变动范围在17—19 之间; 大多数(83%)的三色菖鲉有19个, 变动范围在18—20 之间。在本研究中, 所得标本的胸鳍鳍条数的数量为 18—19 之间, 且大部分(71%)为19。Barsukov 等(1978)还发现, 就体色而言, 三色菖鲉的体色是棕红色到粉色之间并带有小斑点, 比褐菖鲉的体色更加鲜艳。本研究所采集到的标本的体色均与该描述相符合。此外, 有研究表明, 三色菖鲉的体型可以成长到比褐菖鲉大(Tokimuraet al, 2001), 且三色菖鲉所生活的水层比褐菖鲉更深(Amaoka, 1984; Shimizu, 1984a, b)。
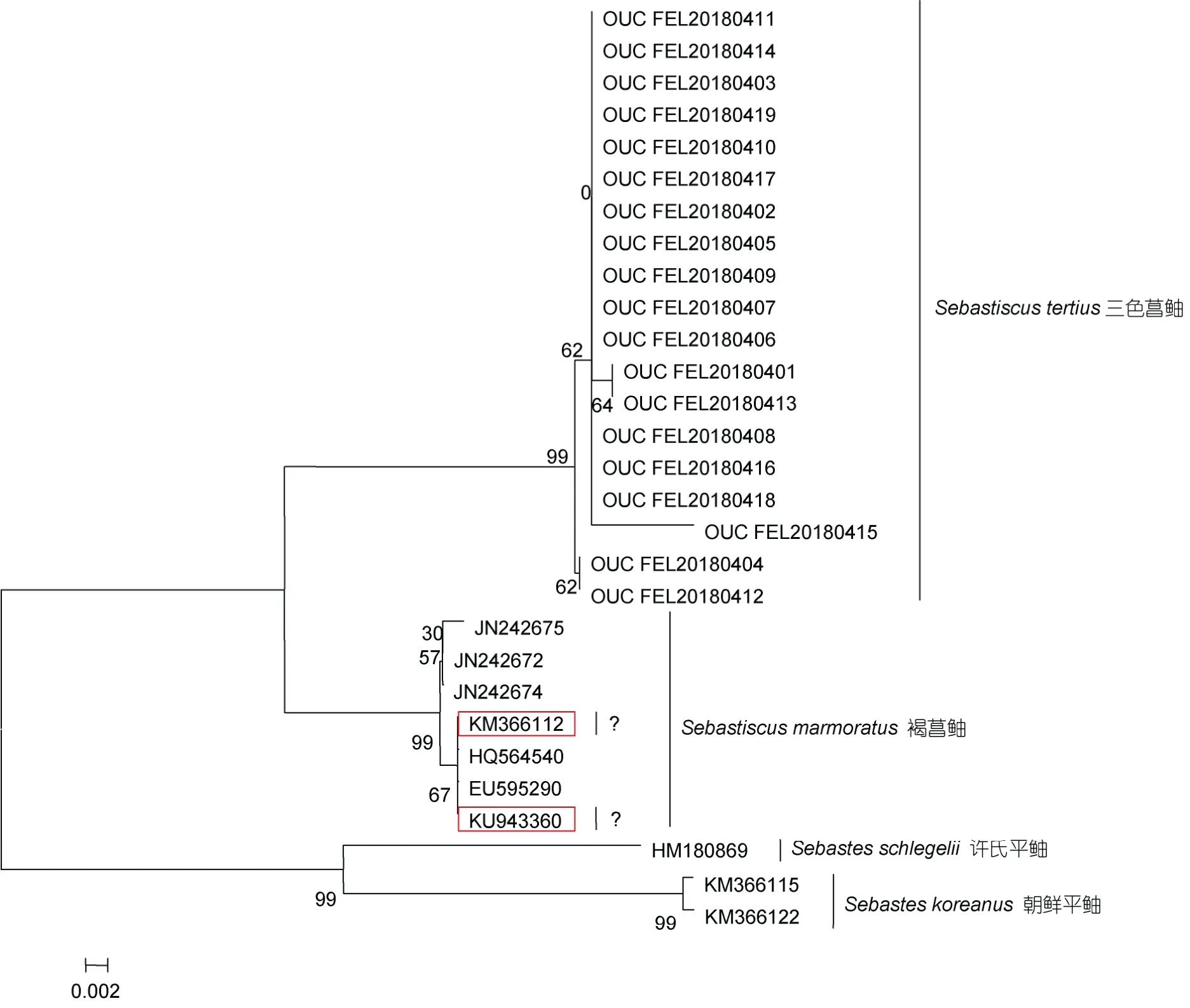
图4 基于COI 序列, 以朝鲜平 鲉 和许氏平 鲉 为外群运用邻接法构建的三色菖 鲉系统发育树 Fig.4 Phylogenetic tree of Sebastiscus tertius using Sebastes koreanus and Sebastes schlegelii as outgroups based on neighbor-joining method and COI sequences
线粒体12S rRNA 基因序列具有相对保守的进化速率, 可以很好地区分种间和属间的亲缘关系, 并已经广泛地应用于鱼类等脊椎动物的分子系统学等研究(唐优良等, 2011)。Miya 等(2015)通过对比大量物种线粒体DNA 的12S rRNA 基因序列, 找到了 其中的一个高变区片段(163—185bp), 并设计了相关引物, 以此对鱼类的科、属、种进行了鉴定。Wang等(2020)也通过使用该序列片段, 从所得1357 个个体中准确鉴别出 37 个物种。本研究所得三色菖鲉12S rRNA 序列与GenBank 中已有的褐菖鲉序列明显分为两个组群, 其同属内的种间遗传距离为0.026, 高于种内遗传变异的物种划界阈值(≥0.02) (Hebertet al, 2004), 且与GenBank 中已有的三色菖鲉序列(序列号为LC026726 和AB969892)完全一致, 可作为三色菖鲉的微型DNA 条形码。
Hebert 等(2003b)通过对大量物种的线粒体COI基因序列进行分析, 表明COI 基因序列能够很好地区分动物物种, 是适合用来对物种进行鉴定的DNA条形码标准基因。本研究所得三色菖鲉COI 序列与GenBank 中已有的三色菖鲉序列(序列号为KM366112 和KU943360)通过构建系统发育树明显分为两个组群, 且网上已有的三色菖 鲉序列与GenBank 已有的褐菖鲉序列聚为一支, 其同属内的种间遗传距离为0.044, 超过了种内平均遗传变异的物种划界阈值(≥0.02) (Hebertet al, 2004), 推测这两条序列所对应的样品可能为褐菖鲉而非三色菖鲉。
4 结论
本文报道了浙江舟山海域一新纪录种: 三色菖鲉, 并描述其形态以及研究其DNA 条形码, 表明在鉴定形态极其相似的物种时对其DNA 条形码研究的必要性。三色菖鲉学名为Sebastiscus tertius(Barsukov and Chen, 1978), 其广泛分布于西太平洋, 在我国分布于东海、南海以及台湾海域。本研究结果对于中国菖鲉属鱼类的分类具有积极的参考作用, 同时为中 国近海菖鲉属鱼类研究提供基础资料。此外, 如果三色菖鲉为近几年来才进入舟山近海海域的物种, 其出现可能会对舟山当地生态系统产生不利影响, 进一步的生物学和遗传学研究有助于更好地了解该物种的种群历史和该物种的分布范围变化。
猜你喜欢
昆明医科大学学报(2022年4期)2022-05-23
中国宝玉石(2021年5期)2021-11-18
黄河·黄土·黄种人(华夏文明)(2021年6期)2021-09-28
少年文艺·开心阅读作文(2021年8期)2021-09-05
现代仪器与医疗(2021年1期)2021-06-09
石油化工建设(2020年1期)2020-08-24
小学科学(学生版)(2019年5期)2019-05-21
学苑创造·B版(2017年10期)2017-12-21
兵器知识(2017年10期)2017-10-19
学生天地·初中(2014年1期)2014-02-17
